
The Evolution of an Ancient Coastal Lake (Lerna, Peloponnese, Greece)

 , , , ,
, , , ,  ,
,  , , and
, , and
Abstract
:1. Introduction
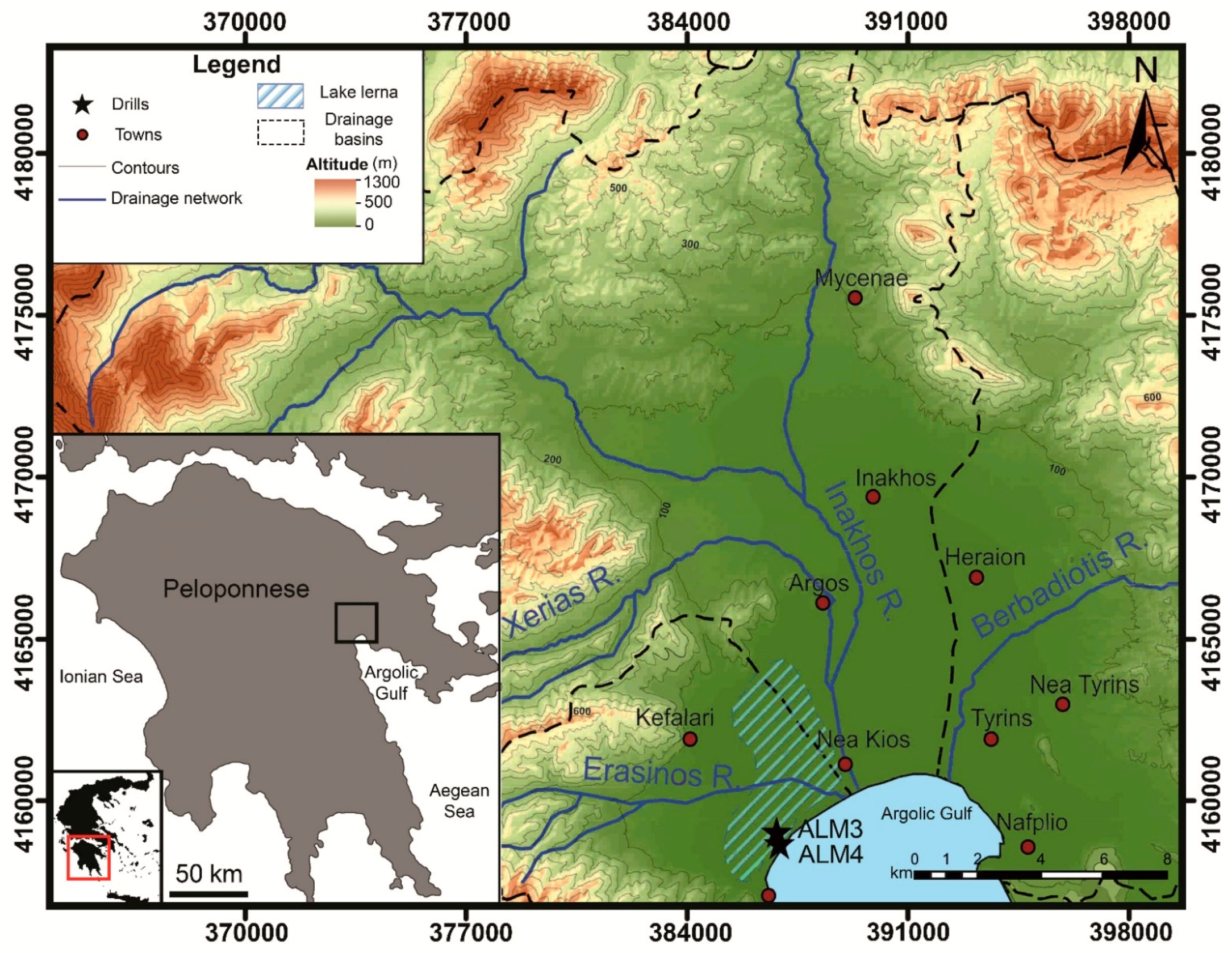
2. Geology and Geomorphology
3. Materials and Methods
4. Results
4.1. Radiocarbon Dating
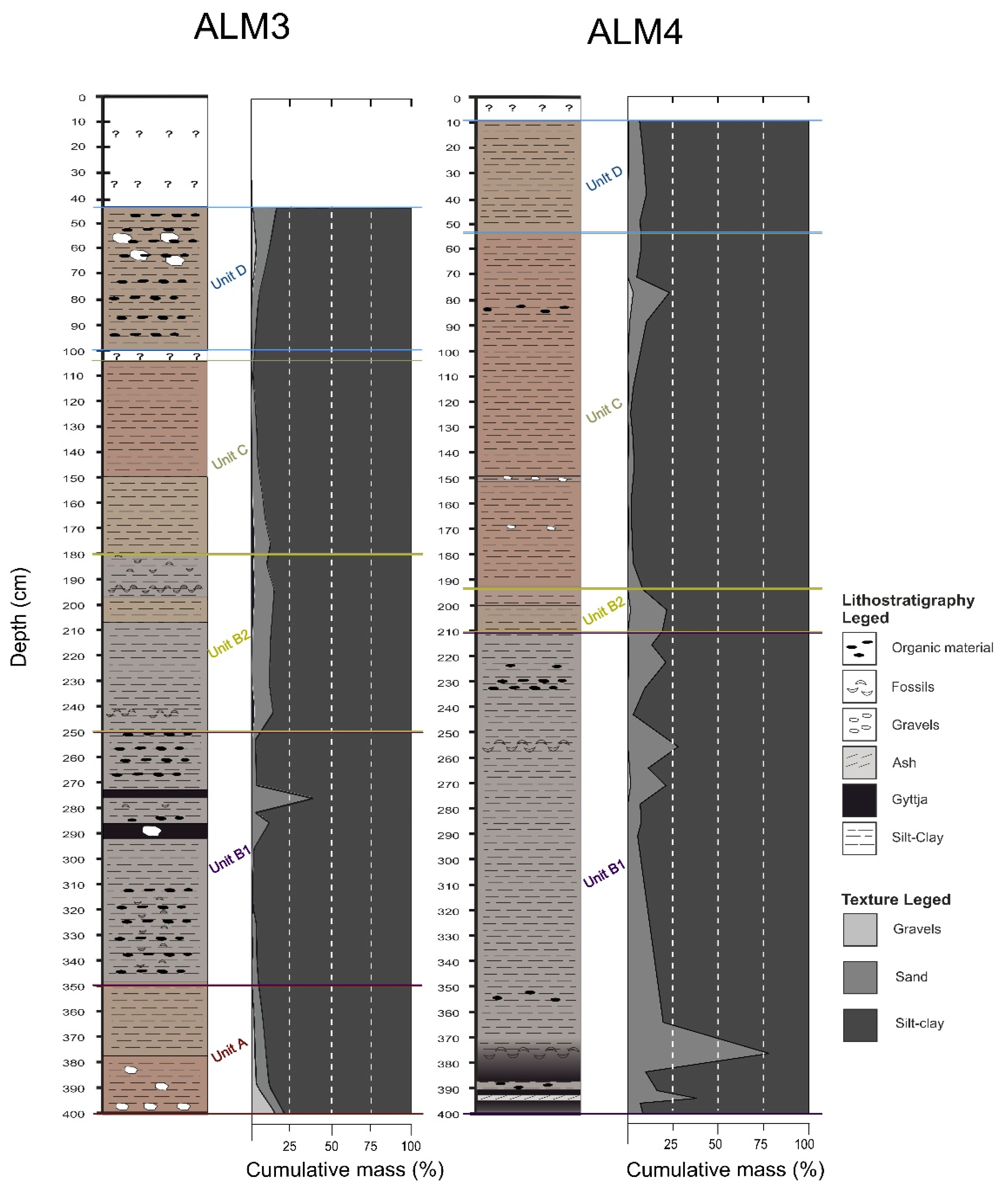
4.2. Cores’ Lithostratigraphy
4.2.1. Fluvial Deposits—Sedimentological Unit A (SUA)
4.2.2. Shallow Lake Swamp Deposits—Sedimentological Unit B (SUB)
4.2.3. Lake Deposits—Sedimentological Unit C (SUC)
4.2.4. Anthropogenic Deposits—Sedimentological Unit D (SUD)
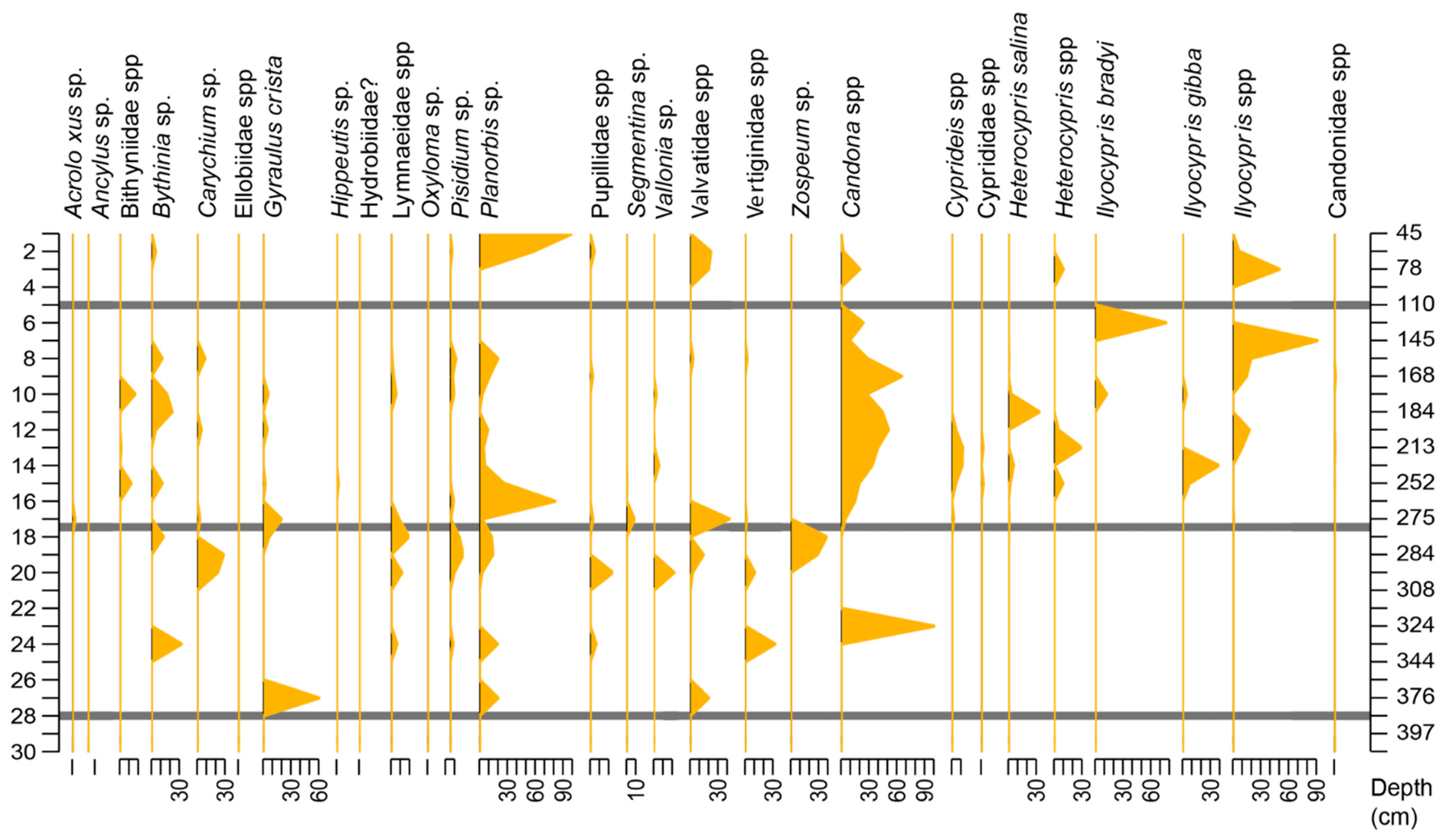
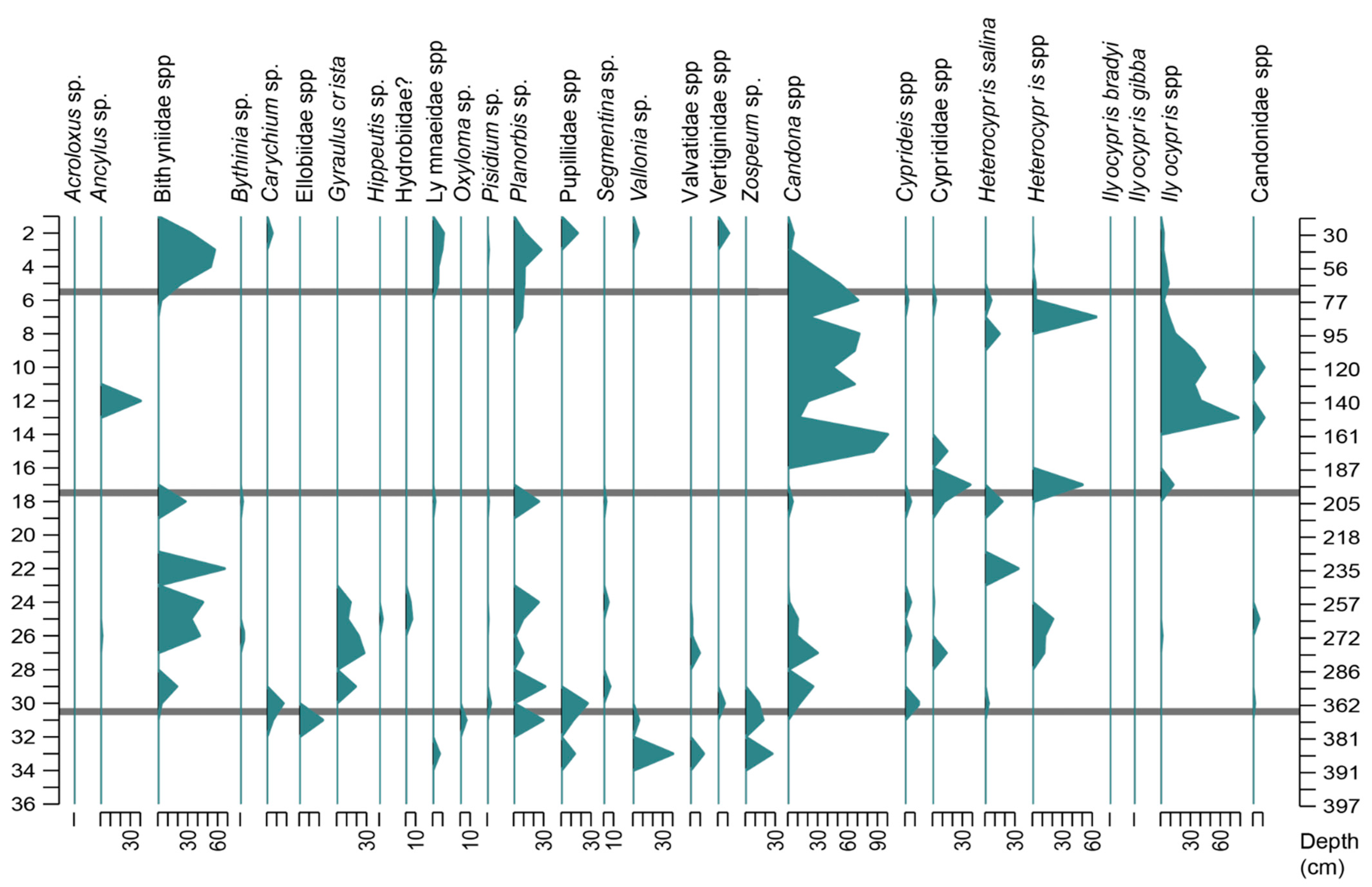
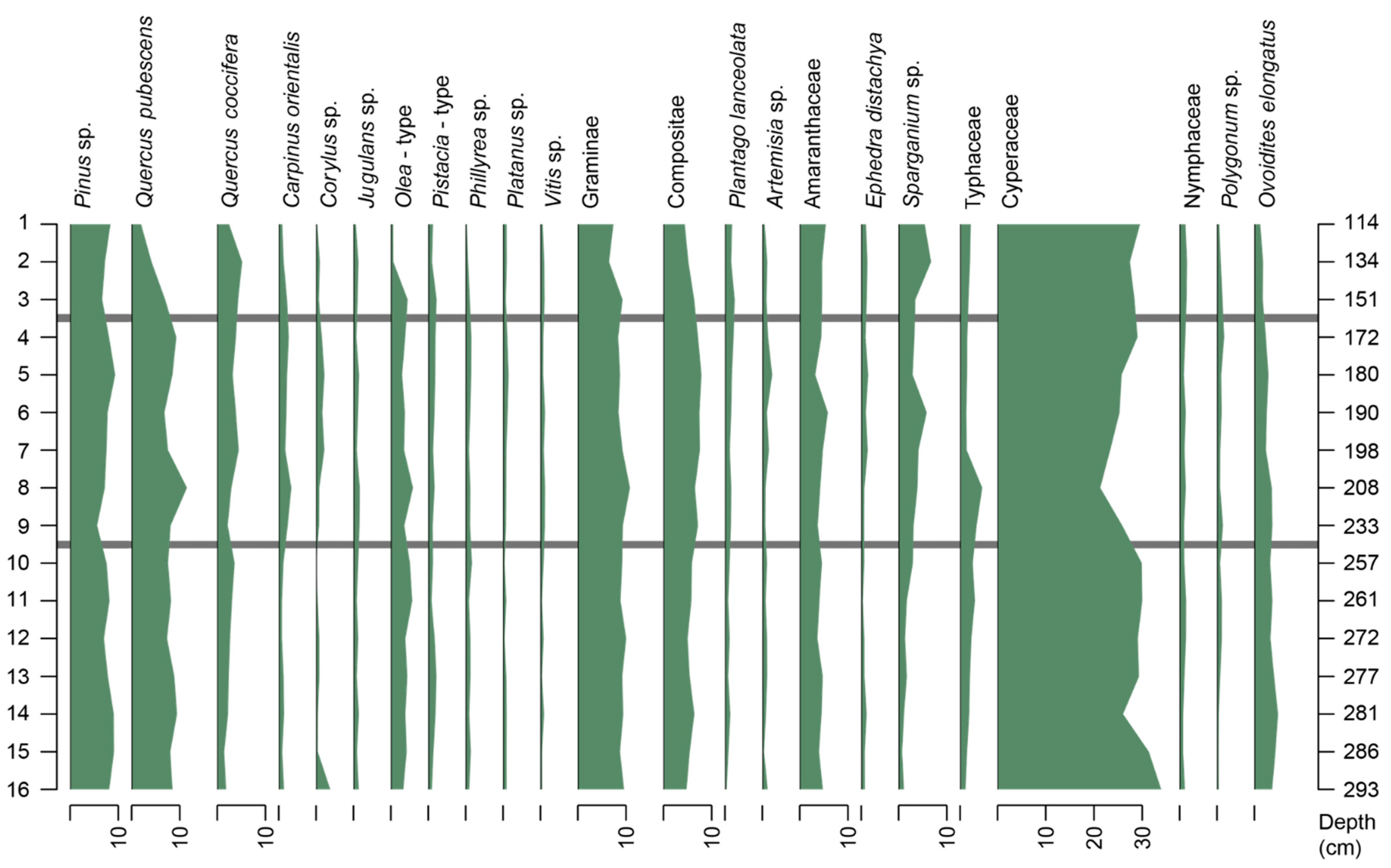
4.3. Palaeobiological Analysis
4.3.1. Palaeofauna

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Class | Families | Genera | Species | General Habitat | Environment |
|---|---|---|---|---|---|
| Gastropoda | Planorbidae | Planorbis | cf. planorbis | Freshwater | Low energy temporary and permanent ponds, streams, rivers, springs, lakes [32] |
| Planorbis | cf. atticus | ||||
| Planorbis | sp. | ||||
| Gyraulus | crista | ||||
| Hippeutis | sp. | ||||
| Segmentina | sp. | ||||
| Valvatidae | Valvata | cristata | Freshwater | Cold, clean lakes, rivers, streams [7] | |
| Valvata | sp. | ||||
| Bithyniidae | Bithynia | sp. | Freshwater | Quiet muddy rivers, lakes, ponds, canals, swamps [7] | |
| Lymnaeidae | Stagnicola | sp. | Freshwater | Flowing rivers and streams, lakes to stagnant ponds, swamps [7] | |
| – | sp. | ||||
| Ellobiidae | Carychium | sp. | Permanently wet epigean environments | Aphotic, permanently wet terrestrial biomes [7] | |
| Zospeum | sp. | Permanently wet subterranean environments | Cave karstic rock crevice dweller [35] | ||
| Vertiginidae | Vertigo | antivertigo | Terrestrial on water margin vegetation | Wet, unimproved pasture, marshes and tall fen or water margin vegetation [36] | |
| Pupillidae | Pupilla | sp. | Detritus feeder (dead plant remains) [36] | ||
| Valloniidae | Vallonia | sp. | Terrestrial moist environment with plant debris | Humid and wet habitats in lowlands, on humid and uncultivated meadows and in calcareous swamps [36] | |
| Acroloxidae | Acroloxus | sp. | Fresh water | Lakes [7] | |
| Ancylus | sp. | ||||
| Hydrobiidae | “Hydrobia” | sp. | Freshwater | Springs, streams, rivers, lakes, groundwater systems, estuarine marshes [36] | |
| Succineidae | Oxyloma | sp. | Terrestrial on water margin vegetation | Feeds on wilting plant parts [36] | |
| Bivalvia | Sphaerioidea | Pisidium | personatum | Freshwater | Bottom dwelling filter feeder [37] |
| Pisidium | sp. |
4.3.2. Palaeoflora
5. Discussion
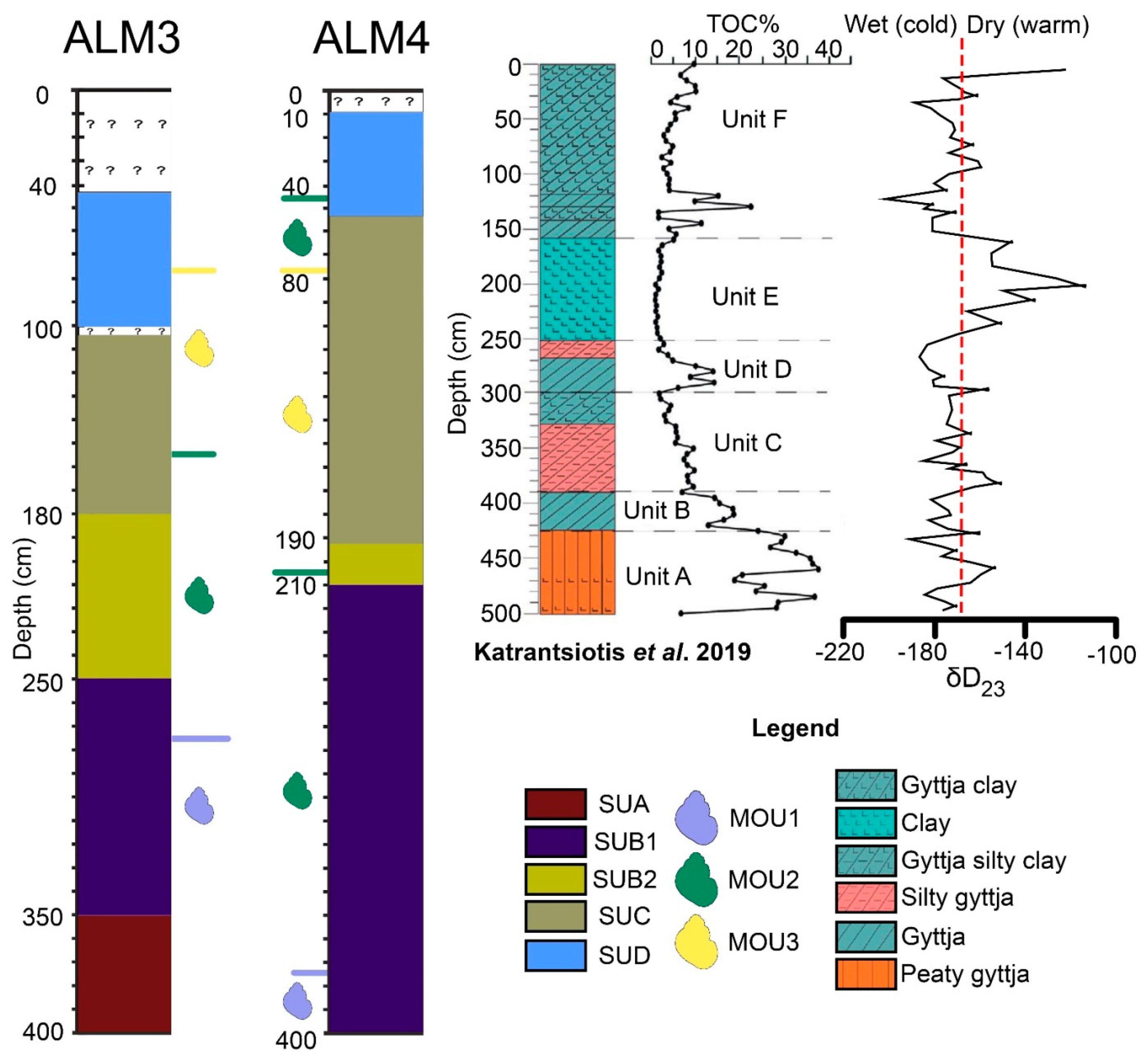
5.1. Sedimentology
- Unit B (390–425 cm) of [12], characterised as gyttja with high TOC values, is equivalent to SUB1 of ALM3 (250–350 cm) and ALM4 (350–400 cm) (Figure 8), as these are the levels with the highest concentrations of organic material. The peat deposits seen here between gyttja are rather thin. In [12], Unit B was estimated to have been deposited around 4000 BP.
- The deepest part of Unit C from [12]’s core could be related to the lower SUB2 of ALM3 (208–250 cm) and the middle SUB1 of ALM4 (270–350 cm) (Figure 8). These units have similar silty sediment, as they are characterised by medium to low concentrations in TOC. In [12], the Unit C was dated around 3700 BP.
- The upper Unit C and Unit D of [12] correspond to the upper part of SUB2 (208–180 cm) of ALM3 and the upper part of SUB1 (270–210 cm) and SUB2 of ALM4 (Figure 8). The latter units have similarities in sediment composition as well as in organic material concentration (gyttja silty clay). In [12], the these units were dated around 2500 BP.
- The lower part of Unit F (120–155 cm) of [12] can be compared with lower SUD of ALM3 (75–100 cm) and upper SUC of ALM4 (55–100 cm) (Figure 8). These units are all characterised by clayey sediment with similar concentrations of organic material. The age given for lower Unit F of [12] is approximately 1500 BP.
5.2. Palaeofauna and Flora
5.3. Palaeobiological Evolution of the Lake
5.4. What Is Causing These Environmental Fluctuations?
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Economidou, E. The Ecological Value of Coastal Ecosystems. Ekistics 1982, 49, 98–101. [Google Scholar]
- Paine, L. The Sea and Civilization: A Maritime History of the World; Knopf Doubleday Publishing Group: New York, NY, USA, 2013. [Google Scholar]
- Mariolakos, I.D. Ancient Greece and Water: Climatic Changes, Extreme Events, Water Management, and Rivers in Ancient Greece. Handb. Environ. Chem. 2017, 59, 3–30. [Google Scholar] [CrossRef]
- Zangger, E. Prehistoric Coastal Environments in Greece: The Vanished Landscapes of Dimini Bay and Lake Lema. J. Field Archaeol. 1991, 18, 1–15. [Google Scholar] [CrossRef]
- Oertel, G.F. Coastal Lakes and Lagoons. Encycl. Earth Sci. Ser. 2005, 14, 263–266. [Google Scholar] [CrossRef]
- Barbieri, R.; Zogaris, S.; Kalogianni, E.; Stoumboudi, M.T.; Chatzinikolaou, Y.; Giakoumi, S.; Kapakos, Y.; Kommatas, D.; Koutsikos, N.; Tachos, V.; et al. Freshwater Fishes and Lampreys of Greece: An Annotated Checklist; Monographs on Marine Sciences No. 8; Hellenic Centre for Marine Research: Athens, Greece, 2015; p. 130. [Google Scholar]
- Strong, E.E.; Gargominy, O.; Ponder, W.F.; Bouchet, P. Global Diversity of Gastropods (Gastropoda; Mollusca) in Freshwater. Hydrobiologia 2008, 595, 149–166. [Google Scholar] [CrossRef]
- Caskey, J.L.; Blackburn, E.T. Lerna in the Argolid; American School of Classical Studies at Athens: Athens, Greece, 1997. [Google Scholar]
- Weiberg, E.; Unkel, I.; Kouli, K.; Holmgren, K.; Avramidis, P.; Bonnier, A.; Dibble, F.; Finné, M.; Izdebski, A.; Katrantsiotis, C.; et al. The Socio-Environmental History of the Peloponnese during the Holocene: Towards an Integrated Understanding of the Past. Quat. Sci. Rev. 2016, 136, 40–65. [Google Scholar] [CrossRef] [Green Version]
- Norström, E.; Katrantsiotis, C.; Finné, M.; Risberg, J.; Smittenberg, R.H.; Bjursäter, S. Biomarker Hydrogen Isotope Composition (ΔD) as Proxy for Holocene Hydroclimatic Change and Seismic Activity in SW Peloponnese, Greece. J. Quat. Sci. 2018, 33, 563–574. [Google Scholar] [CrossRef]
- Gejvall, N.-G. Prehistoric Environmental Destruction, Lerna (Greece). Museum Int. 1973, 25, 93–96. [Google Scholar] [CrossRef]
- Katrantsiotis, C.; Norström, E.; Smittenberg, R.H.; Finne, M.; Weiberg, E.; Hättestrand, M.; Avramidis, P.; Wastegård, S. Climate Changes in the Eastern Mediterranean over the Last 5000 years and Their Links to the High-Latitude Atmospheric Patterns and Asian Monsoons. Glob. Planet. Chang. 2019, 175, 36–51. [Google Scholar] [CrossRef]
- Janssen, J.; Rodwell, J.; Criado, M.G.; Gubbay, S.; Haynes, T.; Nieto, A.; Sanders, N.; Landucci, F.; Loidi, J.; Ssymank, A.; et al. European Red List of Habitats Part 2. Terrestrial and Freshwater Habitats; Publications Office of the European Union: Luxembourg, 2016. [Google Scholar] [CrossRef]
- Cuttelod, A.; Seddon, M.; Neubert, E. European Red List of Non-Marine Molluscs; Publications Office of the European Union: Luxembourg, 2011. [Google Scholar] [CrossRef]
- Kelletat, D.; Kowalczyk, G.; Schröder, B.; Winter, K.-P. A Synoptic View on the Neotectonic Development of the Peloponnesian Coastal Regions. Z. Dtsch. Geol. Ges. 1976, 127, 447–465. [Google Scholar] [CrossRef]
- van Andel, T.H.; Zangger, E.; Demitrack, A. Land Use and Soil Erosion in Prehistoric and Historical Greece. J. Field Archaeol. 1990, 17, 379. [Google Scholar] [CrossRef]
- Finke, E.A. Landscape Evolution of the Argive Plain, Greece: Paleoecology, Holocene Depositional History, and Coastline Changes; Stanford University: Stanford, CA, USA, 1988. [Google Scholar]
- Zangger, E. The Geoarchaeology of the Argolid. Am. J. Archaeol. 1993, 100, 636. [Google Scholar] [CrossRef]
- Koutsouveli-Nomikou, A. Contribution a l’Etude Quaternaire de la Région d’Argos (Nord-Est du Péloponnèse, Gréce); Université d’Aix: Marseille, France, 1980. [Google Scholar]
- Gaki-Papanastassiou, K. The Geomorphological Evolution in and around the Argive Plain since the Quaternary; National and Kapodistrian University of Athens: Athens, Greece, 1991. [Google Scholar] [CrossRef]
- Seni, A.; Gaki-Papanastassiou, K.; Karymbalis, E.; Zouva, C. Anticipated Sea-Level Rise Vulnerability Assessment Along Part of the Eastern Argolic Gulf and the Argive Plain With the Use of G.I.S. In Proceedings of the 8th Pan-Hellenic Geographical Conference, Athens, Greece, 4–7 October 2007; pp. 146–155. [Google Scholar]
- Gaki-Papanastassiou, K.; Maroukian, H. Late Quaternary controls on river behaviour in the eastern part of the Argive plain, eastern Peloponnesus, Greece. In Proceedings of the International Symposium on Mediterranean Quaternary River Environments, Cambridge, UK, 28–29 September 1992; Balkema: Rotterdam, The Netherlands, 1995; pp. 89–95. [Google Scholar]
- Liritzis, I.; Raftopoulou, M. Argolid: Connection of Prehistoric Legends with Geoenvironmental and Archaeological Evidence. Tubaar 1999, 2, 87–99. [Google Scholar] [CrossRef] [Green Version]
- De Wever, P. Etude Géologique des Series Apparaissant en Fenêtre sous l’ Allochtone Pindique (Série de Tripolitza et Série Épimetamorphic de Zarouchla), Péloponnèse Septentrional, Gréce; Université de Lille: Lille, Greece, 1975. [Google Scholar]
- Tataris, A.; Kounis, G.; Maragoudakis, N. Geological Map of Greece on a Scale 1:50,000, “Vagia” Sheet; IGME: Madrid, Spain, 1970. [Google Scholar]
- Exidavelonis, P.; Taktikos, S. Geological Map of Greece on a Scale 1:50,000, “Tripolis” Sheet; IGME: Madrid, Spain, 1988. [Google Scholar]
- Geological Map of Greece on a Scale 1:50,000, “Argos” Sheet; IGME: Madrid, Spain, 1960.
- Theodoropoulos, D.; Papapetrou-Zamani, A. Geological and Geomorphological Investigations of the Argos Plain. Ann. Géologiques Pays Hélléniques 1970, 22, 269–294. [Google Scholar]
- Folk, R.L.; Ward, W.C. Brazos River Bar [Texas]; a Study in the Significance of Grain Size Parameters. J. Sediment. Res. 1957, 27, 3–26. [Google Scholar] [CrossRef]
- Blott, S.J.; Pye, K. Gradistat: A Grain Size Distribution and Statistics Package for the Analysis of Unconsolidated Sediments. Earth Surf. Process. Landf. 2001, 26, 1237–1248. [Google Scholar] [CrossRef]
- Psilovikos, A.A. Sediment Analysis at Gibraltar Point, Lincolnshire. East Midl. Geogr. 1979, 7, 128–133. [Google Scholar]
- Rossetti, G.; Bartoli, M.; Martens, K. Limnological Characteristics and Recent Ostracods (Crustacea, Ostracoda) of Freshwater Wetlands in the Parco Oglio Sud (Northern Italy). Ann. Limnol. Int. J. Limnol. 2004, 40, 329–341. [Google Scholar] [CrossRef] [Green Version]
- Aneta, S. Woodland Ponds as an Important Habitat of Hippeutis Complanatus (Linnaeus 1758) Occurrence—Effect of Environmental Factors and Habitat Preferences. Ekológia 2014, 33, 101–115. [Google Scholar] [CrossRef] [Green Version]
- Ruiz, F.; Abad, M.; Bodergat, A.M.; Carbonel, P.; Rodríguez-Lázaro, J.; González-Regalado, M.L.; Toscano, A.; García, E.X.; Prenda, J. Freshwater Ostracods as Environmental Tracers. Int. J. Environ. Sci. Technol. 2013, 10, 1115–1128. [Google Scholar] [CrossRef] [Green Version]
- Weigand, A.M.; Jochum, A.; Slapnik, R.; Schnitzler, J.; Zarza, E.; Klussmann-Kolb, A. Evolution of Microgastropods (Ellobioidea, Carychiidae): Integrating Taxonomic, Phylogenetic and Evolutionary Hypotheses. BMC Evol. Biol. 2013, 13, 18. [Google Scholar] [CrossRef] [Green Version]
- Welter-Schultes, F.W. European Non-Marine Molluscs, a Guide for Species Identification/Bestimmungsbuch für Europäische Land- und Süsswassermollusken; Planet Poster Editions: Goettingen, Germany, 2012. [Google Scholar]
- McMahon, R.F.; Bogan, A.E. 11—MOLLUSCA: BIVALVIA. In Ecology and Classification of North American Freshwater Invertebrates; Thorp, J.H., Covich, A.P., Eds.; Academic Press: Cambridge, MA, USA, 2001; pp. 331–429. [Google Scholar] [CrossRef]
- Abdelhady, A.A.; Khalil, M.M.; Ismail, E.; Fan, J.; Zhang, S.; Xiao, J. Water Chemistry and Substrate Type as Major Determinants of Molluscan Feeding Habit and Life–Mode in Lagoon Sediments. Estuar. Coast. Shelf Sci. 2019, 220, 120–130. [Google Scholar] [CrossRef]
- Jahns, S. On the Holocene Vegetation History of the Argive Plain (Peloponnese, Southern Greece). Veg. Hist. Archaeobot. 1993, 2, 187–203. [Google Scholar] [CrossRef]
- Norström, E.; West, J.; Kouli, K.; Katrantsiotis, C.; Hättestrand, M.; Smittenberg, R.H. Evaluation of Anhydrosugars as a Molecular Proxy for Paleofire Activity: A Case Study on a Holocene Sediment Core from Agios Floros, Peloponnese, Greece. Org. Geochem. 2021, 153, 104193. [Google Scholar] [CrossRef]
- Finné, M.; Holmgren, K.; Shen, C.C.; Hu, H.M.; Boyd, M.; Stocker, S. Late Bronze Age Climate Change and the Destruction of the Mycenaean Palace of Nestor at Pylos. PLoS ONE 2017, 12, e0189447. [Google Scholar] [CrossRef] [Green Version]
- Weiberg, E.; Hughes, R.E.; Finné, M.; Bonnier, A.; Kaplan, J.O. Mediterranean Land Use Systems from Prehistory to Antiquity: A Case Study from Peloponnese (Greece). J. Land Use Sci. 2019, 14, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Angel, J.L. Ecology and Population in the Eastern Mediterranean. World Archaeol. 1972, 4, 88–105. [Google Scholar] [CrossRef]
- Boyd, M. Speleothems from Warm Climates: Holocene Records from the Caribbean and Mediterranean Regions; Department of Physical Geography, Stockholm University: Stockholm, Sweden, 2015. [Google Scholar]
- Weiberg, E.; Bevan, A.; Kouli, K.; Katsianis, M.; Woodbridge, J.; Bonnier, A.; Engel, M.; Finné, M.; Fyfe, R.; Maniatis, Y.; et al. Long-Term Trends of Land Use and Demography in Greece: A Comparative Study. Holocene 2019, 29, 742–760. [Google Scholar] [CrossRef] [Green Version]
- Knapp, A.B.; Manning, S.W. Crisis in Context: The End of the Late Bronze Age in the Eastern Mediterranean. Am. J. Archaeol. 2016, 120, 99–149. [Google Scholar] [CrossRef] [Green Version]
- Middleton, G.D. The Collapse of Palatial Society in LBA Greece and the Postpalatial Period; University of Michigan Press: Ann Arbor, MI, USA, 2010. [Google Scholar] [CrossRef] [Green Version]
- Weiberg, E.; Finné, M. Mind or Matter? People-Environment Interactions and the Demise of Early Helladic II Society in the Northeastern Peloponnese. Am. J. Archaeol. 2013, 117, 1–31. [Google Scholar] [CrossRef]
- Angel, J.L. The People of Lerna: Analysis of a Prehistoric Aegean Population; American School of Classical Studies at Athens: Princeton, NJ, USA, 1971; Volume 2. [Google Scholar]
- Fuchs, M. An Assessment of Human versus Climatic Impacts on Holocene Soil Erosion in NE Peloponnese, Greece. Quat. Res. 2007, 67, 349–356. [Google Scholar] [CrossRef]
- Weiberg, E.; Bonnier, A.; Finné, M. Land Use, Climate Change and ‘Boom-Bust’ Sequences in Agricultural Landscapes: Interdisciplinary Perspectives from the Peloponnese (Greece). J. Anthropol. Archaeol. 2021, 63, 101319. [Google Scholar] [CrossRef]
- Reese, D.S. Faunal Remains from Late Helladic Lerna (Argolid, Greece). Mediterr. Archaeol. Archaeom. 2008, 8, 5–25. [Google Scholar]
- Reese, D.S. Faunal Remains from Early Helladic II Lerna (Argolid Greece). Mediterr. Archaeol. Archaeom. 2013, 13, 289–320. [Google Scholar]








| Sample | Age (BP) | δ13C (‰) | Code | Core Depth | Bulk Sediment Colour |
|---|---|---|---|---|---|
| LTL19897A | 3200 ± 45 | 11.3 ± 0.4 | ALM3 1–2 3.8α | 154 ± 0.5 cm | Brown |
| LTL20303 | 1515 ± 45 | 27.1 ± 0.5 | ALM3 190–193.5 | 190–193.5 cm | Brown-Grey |
| LTL19748A | 5640 ± 45 | 26.9 ± 0.5 | ALM3 2–3 88.5 | 288 ± 0.5 cm | Grey-Black |
| LTL20301A | 596 ± 45 | 21.0 ± 0.5 | ALM4 203–205 | 203–205 cm | Brown |
| LTL20302 | 4610 ± 45 | 28.8 ± 0.6 | ALM4 328–330 | 328–330 cm | Black |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koskeridou, E.; Thivaiou, D.; Psarras, C.; Rentoumi, E.; Evelpidou, N.; Saitis, G.; Petropoulos, A.; Ioakim, C.; Katopodis, G.; Papaspyropoulos, K.; et al. The Evolution of an Ancient Coastal Lake (Lerna, Peloponnese, Greece). Quaternary 2022, 5, 22. https://doi.org/10.3390/quat5020022
Koskeridou E, Thivaiou D, Psarras C, Rentoumi E, Evelpidou N, Saitis G, Petropoulos A, Ioakim C, Katopodis G, Papaspyropoulos K, et al. The Evolution of an Ancient Coastal Lake (Lerna, Peloponnese, Greece). Quaternary. 2022; 5(2):22. https://doi.org/10.3390/quat5020022
Chicago/Turabian StyleKoskeridou, Efterpi, Danae Thivaiou, Christos Psarras, Evangelia Rentoumi, Niki Evelpidou, Giannis Saitis, Alexandros Petropoulos, Chryssanthi Ioakim, George Katopodis, Konstantinos Papaspyropoulos, and et al. 2022. "The Evolution of an Ancient Coastal Lake (Lerna, Peloponnese, Greece)" Quaternary 5, no. 2: 22. https://doi.org/10.3390/quat5020022