
The Stegodon Bonebed of the Middle Pleistocene Archaeological Site Mata Menge (Flores, Indonesia): Taphonomic Agents in Site Formation

 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
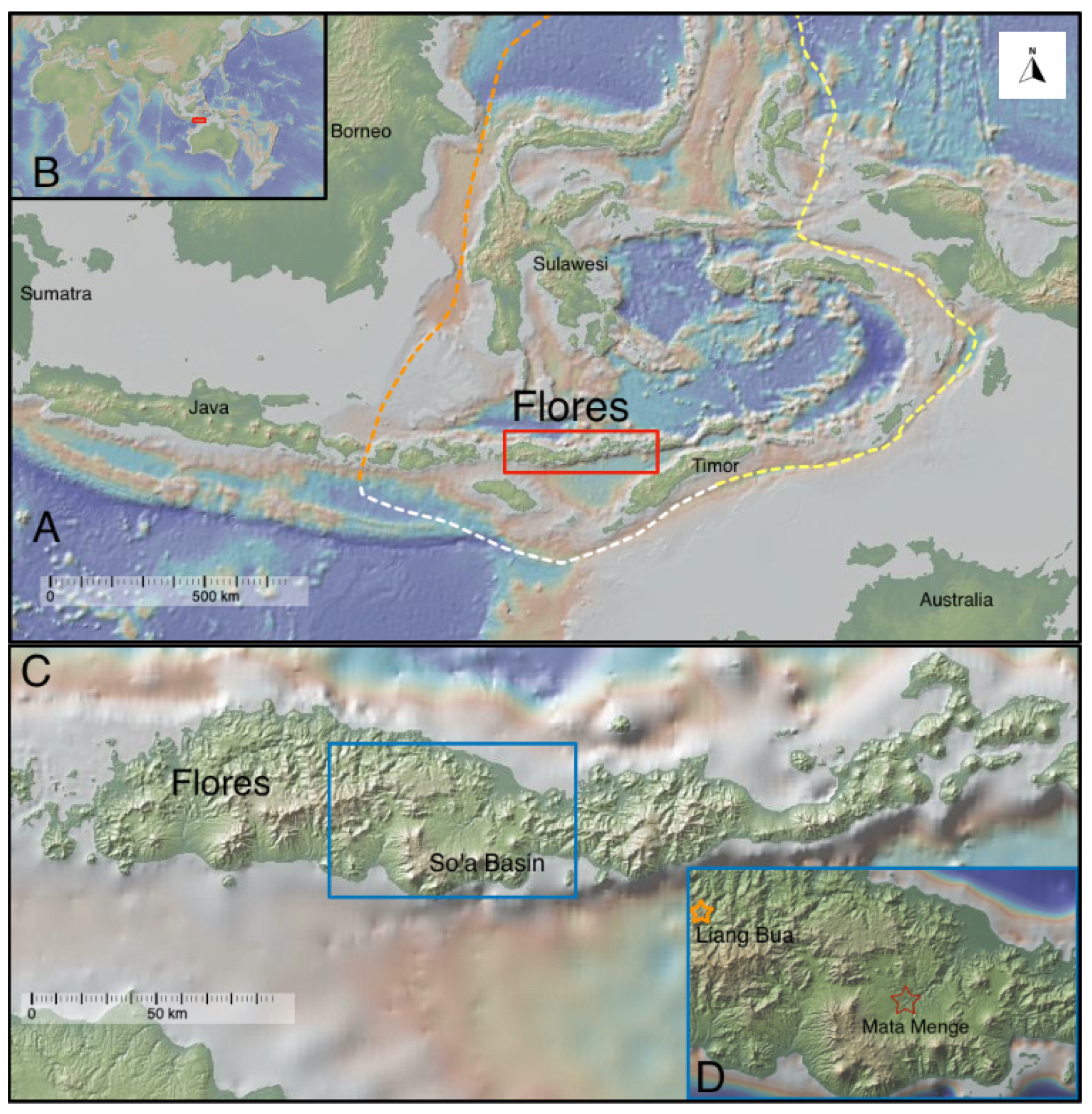
1.1. Geological Context
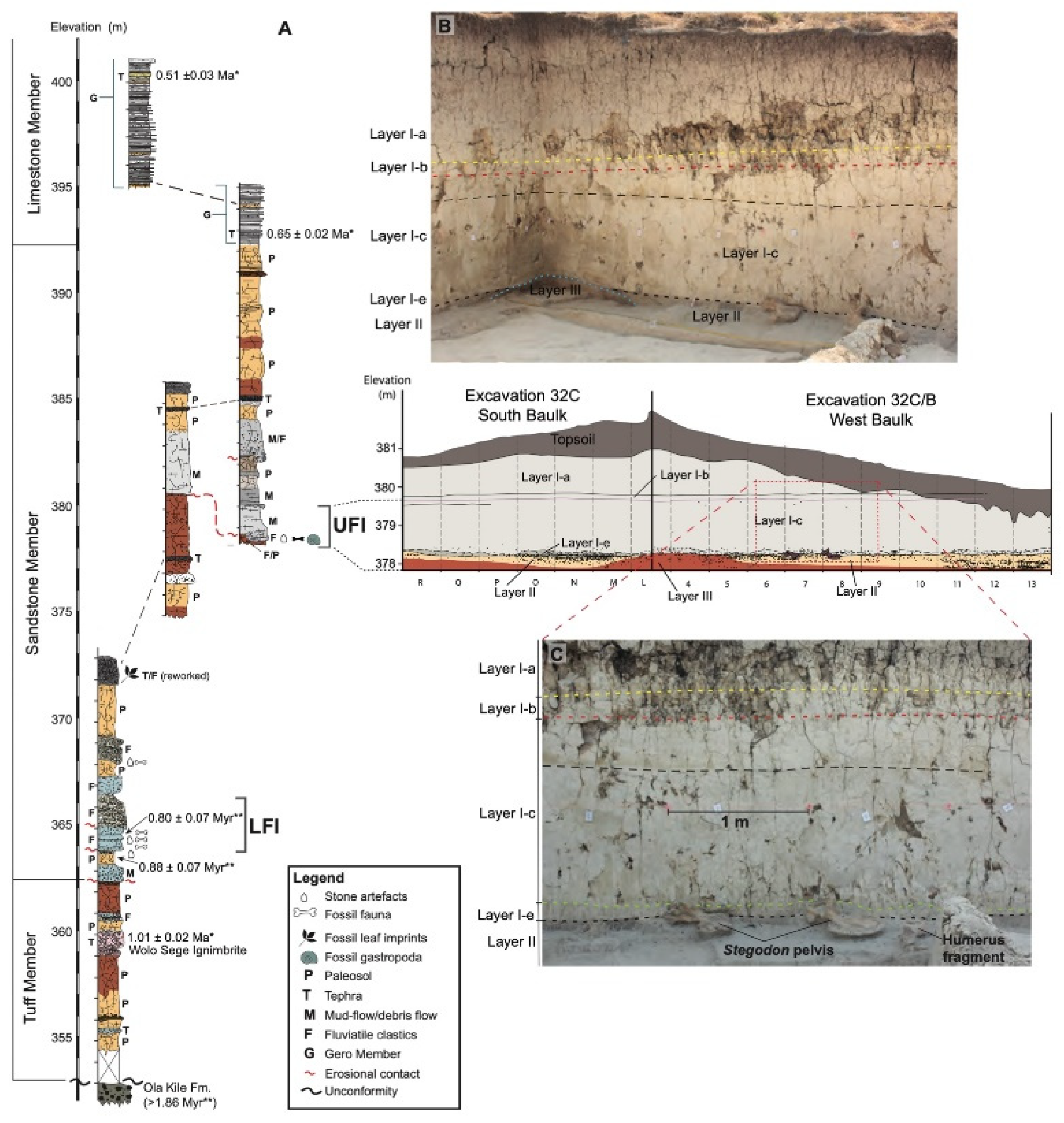
1.2. Stratigraphic Context for Mata Menge
1.3. Location and Fauna
1.4. Taphonomic Processes
2. Materials and Methods
2.1. Materials
2.2. Taphonomic Analysis
- Stage 0—No signs of cracking or flaking, bone may have tissue attached;
- Stage 1—Cracking usually seen parallel to fibre structure, also mosaic cracking;
- Stage 2—Outer bone show flaking, long thin to extensive flaking;
- Stage 3—External bone patchy to mostly removed;
- Stage 4—Coarsely fibrous rough bone surface, cracks well opened; or
- Stage 5—Bone is fragile and easily broken, loss of original bone shape possible.
3. Results
3.1. Subset 1-Assemblage
3.1.1. Fragmentation Analysis
3.1.2. Weathering
3.1.3. Bone Surface Modifications
3.2. Subset 2-Long Limb Bones
3.2.1. Identification
3.2.2. Weathering
3.2.3. Fracturing
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hooijer, D.A. A stegodon from flores. Treubia 1957, 24, 119–128. [Google Scholar]
- Verhoeven, T. Pleistozäne Funde in Flores. Anthropos 1958, 53, 264–265. [Google Scholar]
- Aziz, F.; Morwood, M.J. Introduction: geology, palaeontology and archaeology of the Soa Basin, Central Flores, Indonesia. In Pleistocene Geology, Palaeontology and Archaeology of the Soa Basin, Central Flores, Indonesia; Aziz, F., Morwood, M.J., van den Bergh, G.D., Eds.; Pusat Survei Geologi, Badan Geologi, Departemen Energi dan Sumber Daya Mineral: Bandung, Indonesia, 2009; pp. 1–18. [Google Scholar]
- van den Bergh, G.D.; Kurniawan, I.; Morwood, M.J.; Lentfer, C.J.; Suyono; Setiawan, R.; Aziz, F. Environmental reconstruction of the Middle Pleistocene archaeological/palaeontological site Mata Menge, Flores, Indonesia. In Pleistocene Geology, Palaeontology and Archaeology of the Soa Basin, Central Flores, Indonesia; Aziz, F., Morwood, M.J., van den Bergh, G.D., Eds.; Pusat Survei Geologi, Badan Geologi, Departemen Energi dan Sumber Daya Mineral: Bandung, Indonesia, 2009; pp. 59–94. [Google Scholar]
- van den Bergh, G.D.; Mubroto, B.; Aziz, F.; Sondaar, P.Y.; Vos, J. Did Homo erectus reach the Island of Flores? Bull. Indo-Pac. Prehist. Assoc. 1996, 14, 27–36. [Google Scholar] [CrossRef]
- van den Bergh, G.D. The Late Neogene Elephantoid-Bearing Faunas of Indonesia and Their Palaeozoogeographic Implications: A Study of the Terrestrial Faunal Succession of Sulawesi, Flores and Java, Including Evidence for Early Hominid Dispersal East of Wallace’s Line. Ph.D. Thesis, University of Utrecht, Utrecht, The Netherlands, 1999. [Google Scholar]
- Sondaar, P.Y.; van den Bergh, G.D.; Mubroto, B.; Aziz, F.; de Vos, J.; Batu, U.L. Middle Pleistocene faunal turnover and colonization of Flores (Indonesia) by Homo erectus. Comptes Rendus de l’Academie des Sciences de Paris 1994, 319, 1255–1262. [Google Scholar]
- Brumm, A.; Aziz, F.; van den Bergh, G.D.; Morwood, M.J.; Moore, M.W.; Kurniawan, I.; Hobbs, D.R.; Fullagar, R. Early stone technology on Flores and its implications for Homo floresiensis. Nature 2006, 441, 624–628. [Google Scholar] [CrossRef]
- Brumm, A.; Jensen, G.M.; van den Bergh, G.D.; Morwood, M.J.; Kurniawan, I.; Aziz, F.; Storey, M. Hominins on Flores, Indonesia, by one million years ago. Nature 2010, 464, 748–752. [Google Scholar] [CrossRef]
- Brumm, A.; van den Bergh, G.D.; Storey, M.; Kurniawan, I.; Alloway, B.V.; Setiawan, R.; Setiyabudi, E.; Gruen, R.; Moore, M.W.; Yurnaldi, D.; et al. Age and context of the oldest known hominin fossils from Flores. Nature 2016, 534, 249. [Google Scholar] [CrossRef] [Green Version]
- Morwood, M.J.; Aziz, F.; van den Bergh, G.D.; Sondaar, P.Y.; De Vos, J. Stone Artefacts from the 1994 Excavation at Mata Menge, West Central Flores, Indonesia. Aust. Archaeol. 1997, 44, 26–34. [Google Scholar] [CrossRef]
- Morwood, M.J.; O’Sullivan, P.B.; Aziz, F.; Raza, A. Fission-track ages of stone tools and fossils on the east Indonesian Island of Flores. Nature 1998, 392, 173–176. [Google Scholar] [CrossRef]
- O’Sullivan, P.B.; Morwood, M.; Hobbs, D.; Aziz Suminto, F.; Situmorang, M.; Raza, A.; Maas, R. Archaeological implications of the geology and chronology of the Soa basin, Flores, Indonesia. Geology 2001, 29, 607. [Google Scholar] [CrossRef]
- van den Bergh, G.D.; Kaifu, Y.; Kurniawan, I.; Kono, R.T.; Brumm, A.; Setiyabudi, E.; Aziz, F.; Morwood, M.J. Homo floresiensis-like fossils from the early middle Pleistocene of Flores. Nature 2016, 534, 245–248. [Google Scholar] [CrossRef]
- Yurnaldi, D.; Setiawan, R.; Patriani, E. The Magnetostratigraphy and the Age of So’a Basin Fossil-Bearing Sequence, Flores, Indonesia. Indones. J. Geosci. 2018, 5. [Google Scholar] [CrossRef]
- Sutikna, T.; Tocheri, M.W.; Morwood, M.J.; Saptomo, E.W.; Jatmiko; Awe, R.D.; Wasisto, S.; Westaway, K.E.; Aubert, M.; Li, B.; et al. Revised stratigraphy and chronology for Homo floresiensis at Liang Bua in Indonesia. Nature 2016, 532, 366–369. [Google Scholar] [CrossRef]
- Meijer, H.J.M.; D’Errico, F.; Queffelec, A.; Kurniawan, I.; Setiabudi, E.; Sutisna, I.; Brumm, A.; van den Bergh, G.D. Characterization of bone surface modifications on an early to middle Pleistocene bird assemblage from Mata Menge (Flores, Indonesia) using multifocus and confocal microscopy. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2019, 529, 1–11. [Google Scholar] [CrossRef]
- Meijer, H.J.M.; Kurniawan, I.; Setiabudi, E.; Brumm, A.; Sutikna, T.; Setiawan, R.; van den Bergh, G.D. Avian remains from the early/middle Pleistocene of the So’a Basin, central Flores, Indonesia, and their palaeoenvironmental significance. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2015, 440, 161–171. [Google Scholar] [CrossRef]
- Ali, J.R.; Heaney, L.R. Wallace’s line, Wallacea, and associated divides and areas: history of a tortuous tangle of ideas and labels. Biol. Rev. 2021. [Google Scholar] [CrossRef]
- Suminto; Morwood, M.J.; Kurniawan, I.; Aziz, F.; van den Bergh, G.D.; Hobbs, D.R. Geology and fossil sites of the Soa Basin, Flores, Indonesia. In Pleistocene Geology, Palaeontology and Archaeology of the Soa Basin, Central Flores, Indonesia; Aziz, F., Morwood, M.J., van de Bergh, G.D., Eds.; Pusat Survei Geologi, Badan Geologi, Departemen Energi dan Sumber Daya Mineral: Bandung, Indonesia, 2009; pp. 19–40. [Google Scholar]
- Aziz, F.; van den Bergh, G.D.; Morwood, M.J.; Hobbs, D.R.; Kurniawan, I.; Collins, J.; Jatmiko. Excavations at Tangi Talo, central Flores, Indonesia. In Pleistocene Geology, Palaeontology and Archaeology of the Soa Basin, Central Flores, Indonesia; Aziz, F., Morwood, M.J., van den Bergh, G.D., Eds.; Pusat Survei Geologi, Badan Geologi, Departemen Energi dan Sumber Daya Mineral: Bandung, Indonesia, 2009; pp. 41–58. [Google Scholar]
- Sondaar, P.Y. Faunal evolution and the mammalial biostratigraphy of Java. Cour. Forsch. Inst. Senckenberg 1984, 69, 219–235. [Google Scholar]
- Morwood, M.J.; Aziz, F.; O’Sullivan, P.; Nasruddin; Hobbs, D.R.; Raza, A. Archaeological and palaeontological research in central Flores, east Indonesia: Results of fieldwork 1997-98. Antiquity 1999, 73, 273–286. [Google Scholar] [CrossRef]
- van den Bergh, G.D.; de Vos, J.; Sondaar, P.Y. The late Quaternary palaeogeography of mammal evolution in the Indonesian Archipelago. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2001, 171, 385–408. [Google Scholar] [CrossRef]
- van den Bergh, G.D.; Awe, R.D.; Morwood, M.J.; Sutikna, T.; Jatmiko; Wahyu Saptomo, E. The youngest stegodon remains in Southeast Asia from the Late Pleistocene archaeological site Liang Bua, Flores, Indonesia. Quat. Int. 2008, 182, 16–48. [Google Scholar] [CrossRef]
- van den Bergh, G.D. Insular Fossil Fauna through Deep Geological Time of Wallacea; Geological Survey Center Indonesia, University of Wollongong Australia: Wollongong, NSW, Australia, 2019. [Google Scholar]
- Faith, J.T.; Behrensmeyer, A.K. Changing patterns of carnivore modification in a landscape bone assemblage, Amboseli Park, Kenya. J. Archaeol. Sci. 2006, 33, 1718–1733. [Google Scholar] [CrossRef]
- Haynes, G. Longitudinal studies of African elephant death and bone deposits. J. Archaeol. Sci. 1988, 15, 131–157. [Google Scholar] [CrossRef]
- Haynes, G. Mammoth landscapes: Good country for hunter-gatherers. Quat. Int. 2006, 142, 20–29. [Google Scholar] [CrossRef]
- Haynes, G.; Krasinski, K. Taphonomic Fieldwork in Southern Africa and its Application in Studies of the Earliest Peopling of North America. J. Taphon. 2010, 8, 181–202. [Google Scholar]
- D’Amore, D.C.; Blumenschine, R.J. Komodo monitor (Varanus komodoensis) feeding behaviour and dental function reflected through tooth marks on bone surfaces and the application to ziphodont paleobiology. Paleobiology 2009, 35, 525–552. [Google Scholar] [CrossRef]
- Njau, J.K.; Blumenschine, R.J. A diagnosis of crocodile feeding traces on larger mammal bone, with fossil examples from the Plio-Pleistocene Olduvai Basin, Tanzania. J. Hum. Evol. 2006, 50, 142–162. [Google Scholar] [CrossRef]
- Sahle, Y.; Zaatari Sireen, E.; White Tim, D. Hominid butchers and biting crocodiles in the African Plio–Pleistocene. Proc. Natl. Acad. Sci. USA 2017, 114, 13164. [Google Scholar] [CrossRef] [Green Version]
- Domínguez-Rodrigo, M.; Baquedano, E. Distinguishing butchery cut marks from crocodile bite marks through machine learning methods. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef]
- van den Bergh, G.D.; Meijer, H.J.M.; Due Awe, R.; Morwood, M.J.; Szabo, K.A.; van den Hoek Ostenda, L.W.; Sutikna, T.; Saptomo, E.W.; Piper, P.J.; Dobney, K.M. The Liang Bua faunal remains: a 95 k.yr. sequence from Flores, East Indonesia. J. Hum. Evol. 2009, 57, 527–537. [Google Scholar] [CrossRef]
- Badgley, C. Counting individuals in mammalian fossil assemblages from fluvial environments. PALAIOS 1986, 1, 328–338. [Google Scholar] [CrossRef]
- Brumm, A.; Moore, M.W.; van den Bergh, G.D.; Kurniawan, I.; Morwood, M.J.; Aziz, F. Stone technology at the Middle Pleistocene site of Mata Menge, Flores, Indonesia. J. Archaeol. Sci. 2010, 37, 451–473. [Google Scholar] [CrossRef]
- Dennell, R.W.; Coard, R.; Turner, A. Predators and scavengers in Early Pleistocene southern Asia. Quat. Int. 2008, 192, 78–88. [Google Scholar] [CrossRef]
- Lyman, R.L. Archaeofaunas and Butchery Studies: A Taphonomic Perspective. Adv. Archaeol. Method Theory 1987, 10, 249–337. [Google Scholar]
- Arroyo-Cabrales, J.; Johnson, E.; Morett, L. Mammoth bone technology in the Basin of Mexico. In Bones for Tools—Tools for Bones: the Interplay Between Objects and Objectives; Seetah, K., Gravina, B., Eds.; McDonald Institute for Archaeological Research: Cambridge, UK, 2012; pp. 97–113. [Google Scholar]
- Morlan, R.E. Toward the definition of criteria for the recognition of artificial bone alterations. Quat. Res. 1984, 22, 160–171. [Google Scholar] [CrossRef]
- Villa, P.; Mahieu, E. Breakage patterns of human long bones. J. Hum. Evol. 1991, 21, 27–48. [Google Scholar] [CrossRef]
- Fernandez-Jalvo, Y.; Andrews, P.; Denys, C.; Sese, C.; Stoetzel, E.; Marin-Monfort, D.; Pesquero, D. Taphonomy for taxonomists; implications of predation in small mammal studies. Quat. Sci. Rev. 2016, 139, 138–157. [Google Scholar] [CrossRef]
- Prasanth, T.; Saraswathi, T.R. Histopathological and radiographic evaluation of rat molar teeth after traumatic injury—A pilot study. J. Oral Maxillofac. Pathol. 2012, 16, 313–317. [Google Scholar] [CrossRef]
- Haynes, G.; Krasinski, K.; Wojtal, P. Elephant bone breakage and surface marks made by trampling elephants: Implications for interpretations of marked and broken Mammuthus spp. bones. J. Archaeol. Sci. Rep. 2020, 33, 102491. [Google Scholar] [CrossRef]
- Behrensmeyer, A.K. Vertebrate preservation in fluvial channels. Palaeogr. Palaeoclimatol. Palaeoecol. 1988, 63, 188–199. [Google Scholar] [CrossRef]
- Frison, G.C.; Todd, L.C. The Colby Mammoth Site: Taphonomy and Archeology of a Clovis Kill in Northern Wyoming; University of New Mexica Press: Albuquerque, NM, USA, 1986. [Google Scholar]
- Voorhies, M. Taphonomy and Population Dynamics of an Early Pliocene Vertebrate Fauna, Knox County, Nebraska; University of Wyoming: Laramie, WY, USA, 1969. [Google Scholar]
- Behrensmeyer, A.K. The Taphonomy and Paleoecology of Plio-Pleistocene Vertebrate Assemblages East of Lake Rudolf, Kenya. Bull. Museum Comp. Zool. 1975, 146, 473–578. [Google Scholar]
- Coard, R. One bone, two bones, wet bones, dry bones: Transport potential under experimental conditions. J. Archaeol. Sci. 1999, 26, 1369–1375. [Google Scholar] [CrossRef]
- Coard, R.; Dennel, R.W. Taphonomy of some articulated skeletal remains: Transport potential in an artificial environment. J. Archaeol. Sci. 1995, 22, 441–448. [Google Scholar] [CrossRef]
- Pante, M.C.; Blumenschine, R.J. Fluvial transport of bovid long bones fragmented by the feeding activities of hominin and carnivore. J. Archaeol. Sci. 2010, 37, 846–854. [Google Scholar] [CrossRef]
- Hjulström, F. Studies of the morphological activity of rivers as illustrated by the River Fyris. Bull. Geol. Inst. Univ. Uppsala 1935, 25, 221–527. [Google Scholar]
- Behrensmeyer, A.K. Taphonomic and Ecological Information from Bone Weathering. Paleobiology 1978, 4, 150–162. [Google Scholar] [CrossRef] [Green Version]
- Behrensmeyer, A.K.; Miller, J.H. Building links between ecology and paleontology using taphonomic studies of recent vertebrate communities. In Paleontology in Ecology and Conservation; Louys, J., Ed.; Spring: Berlin/Heidelberg, Germany, 2012; pp. 69–91. [Google Scholar]
- Lyman, R.L.; Fox, G.L. A critical evaluation of bone weathering as an indication of bone assemblage formation. J. Archaeol. Sci. 1989, 16, 293–317. [Google Scholar] [CrossRef]
- Boschian, G.; Caramella, D.; Saccà, D.; Barkai, R. Are there marrow cavities in Pleistocene elephant limb bones, and was marrow available to early humans? New CT scan results from the site of Castel di Guido (Italy). Quat. Sci. Rev. 2019, 215, 86–97. [Google Scholar] [CrossRef]
- Outram, A.K. The Identification and Palaeoeconomic Context of Prehistoric Bone Marrow and Grease Exploitation. Ph.D. Thesis, Thesis, University of Durham, Durham, UK, 1998. [Google Scholar]
- Outram, A.K. A new approach to identifying bone marrow and grease exploitation: Why the “Indeterminate” fragments should not be ignored. J. Archaeol. Sci. 2001, 28, 401–410. [Google Scholar] [CrossRef]
- Yravedra, J.; Panera, J.; Rubio-Jara, S.; Manzano, I.; Expósito, A.; Pérez-González, A.; Soto, E.; López-Recio, M. Neanderthal and Mammuthus interactions at EDAR Culebro 1 (Madrid, Spain). J. Archaeol. Sci. 2014, 42, 500–508. [Google Scholar] [CrossRef]
- Yravedra, J.; Rubio-Jara, S.; Panera, J.; Uribelarrea, D.; Pérez-González, A. Elephants and subsistence. Evidence of the human exploitation of extremely large mammal bones from the Middle Palaeolithic site of PRERESA (Madrid, Spain). J. Archaeol. Sci. 2012, 39, 1063–1071. [Google Scholar] [CrossRef]
- Yravedra, J.; Domínguez-Rodrigo, M.; Santonja, M.; Pérez-González, A.; Panera, J.; Rubio-Jara, S.; Baquedano, E. Cut marks on the Middle Pleistocene elephant carcass of Áridos 2 (Madrid, Spain). J. Archaeol. Sci. 2010, 37, 2469–2476. [Google Scholar] [CrossRef]
- Company, J.; Pereda-Suberbiola, X. Long bone histology of a eusuchian crocodyliform from the Upper Cretaceous of Spain: Implications for growth strategy in extinct crocodiles. Cretac.s Res. 2017, 72, 1–7. [Google Scholar] [CrossRef]
- Albéric, M.; Dean, M.N.; Gourrier, A.; Wagermaier, W.; Dunlop, J.W.C.; Staude, A.; Fratzl, P.; Reiche, I. Relation between the Macroscopic Pattern of Elephant Ivory and Its Three-Dimensional Micro-Tubular Network. PLoS ONE 2017, 12, e0166671. [Google Scholar] [CrossRef] [PubMed]
- Outram, A.K. Bone Fracture and within-bone nutrients: an experimentally based method for investigating levels of marrow extraction. In Consuming Passions and Patterns of Consumption; Scarre, C., Ed.; McDonald Institute for Archaeology: Cambridge, UK, 2002; pp. 51–63. [Google Scholar]
- Boyd, D.K.; Kenmotsu, N.A. The Toyah Phase of Central Texas Late Prehistoric Economic and Social Processes, 1st ed.; Texas A&M University Press: College Station, TX, USA, 2012. [Google Scholar]
- Haynes, G.; Krasinski, K.; Wojtal, P. A Study of Fractured Proboscidean Bones in Recent and Fossil Assemblages. J. Archaeol. Method Theory 2020. [Google Scholar] [CrossRef]
- Johnson, E.V.; Parmenter, P.C.R.; Outram, A.K. A new approach to profiling taphonomic history through bone fracture analysis, with an example application to the Linearbandkeramik site of Ludwinowo 7. J. Archaeol. Sci. Rep. 2016, 9, 623–629. [Google Scholar] [CrossRef] [Green Version]
- Karr, L.; Outram, A.; Hannus, L. A Chronology of Bone Marrow and Bone Grease Exploitation at the Mitchell Prehistoric Indian Village. Plains Anthropol. 2010, 55, 215–223. [Google Scholar] [CrossRef]
- Karr, L.P. Brewster site zooarchaeology reinterpreted: understanding levels of animal exploitation and bone fat production at the Initial Middle Missouri type site. STAR Sci. Technol. Archaeol. Res. 2015, 1, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Outram, A. Distinguishing bone fat exploitation from other taphonomic processes: what caused the high level of bone fragmentation at the Middle Neolithic site of Ajvide, Gotland? In The Zooarchaeology of Fats, Oils, Milk and Dairying. Proceedings of the 9th ICAZ Conference; Oxbow Books: Oxford, UK, 2005. [Google Scholar]
- Holen, S.R. Taphonomy of two last glacial maximum mammoth sites in the central Great Plains of North America: A preliminary report on La Sena and Lovewell. Quat. Int. 2006, 142–143, 30–43. [Google Scholar] [CrossRef]
- van der Geer, A.A.E.; van den Bergh, G.D.; Lyras, G.A.; Prasetyo, U.W.; Due, R.A.; Setiyabudi, E.; Drinia, H. The effect of area and isolation on insular dwarf proboscideans. J. Biogeogr. 2016, 43, 1656–1666. [Google Scholar] [CrossRef] [Green Version]
- Behrensmeyer, A.K. Patterns of natural bone distribution on recent land surfaces: implications for archaeological site formation. In Animals and Archaeology: 1. Hunters and Their Prey; Clutton, B., Grigson, C., Eds.; British Archaeological Reports International Series; British Archaeological: Oxford, UK, 1983; Volume 163. [Google Scholar]
- Lyman, R.L. Vertebrate Taphonomy; Cambridge University Press: Cambridge, UK, 1994. [Google Scholar]
- Thompson, C.E.L.; Ball, S.; Thompson, T.J.U.; Gowland, R. The abrasion of modern and archaeological bones by mobile sediments: the importance of transport modes. J. Archaeol. Sci. 2011, 38, 784–793. [Google Scholar] [CrossRef]
- Fernandez-Jalvo, Y.; Andrews, P. Experimental effects of water abrasion on bone fragments. J. Taphon. 2003, 1, 147–163. [Google Scholar]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Powley, M.J.; Sutisna, I.; Mikac, K.M.; Wibowo, U.P.; van den Bergh, G.D. The Stegodon Bonebed of the Middle Pleistocene Archaeological Site Mata Menge (Flores, Indonesia): Taphonomic Agents in Site Formation. Quaternary 2021, 4, 31. https://doi.org/10.3390/quat4040031
Powley MJ, Sutisna I, Mikac KM, Wibowo UP, van den Bergh GD. The Stegodon Bonebed of the Middle Pleistocene Archaeological Site Mata Menge (Flores, Indonesia): Taphonomic Agents in Site Formation. Quaternary. 2021; 4(4):31. https://doi.org/10.3390/quat4040031
Chicago/Turabian StylePowley, Meagan J., Indra Sutisna, Katarina M. Mikac, Unggul Prasetyo Wibowo, and Gerrit D. van den Bergh. 2021. "The Stegodon Bonebed of the Middle Pleistocene Archaeological Site Mata Menge (Flores, Indonesia): Taphonomic Agents in Site Formation" Quaternary 4, no. 4: 31. https://doi.org/10.3390/quat4040031