
Estuary Stingray (Dasyatis fluviorum) Behaviour Does Not Change in Response to Drone Altitude


Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Site
2.2. Drone and Flight Patterns
2.3. Video Analysis
2.4. Data Analysis
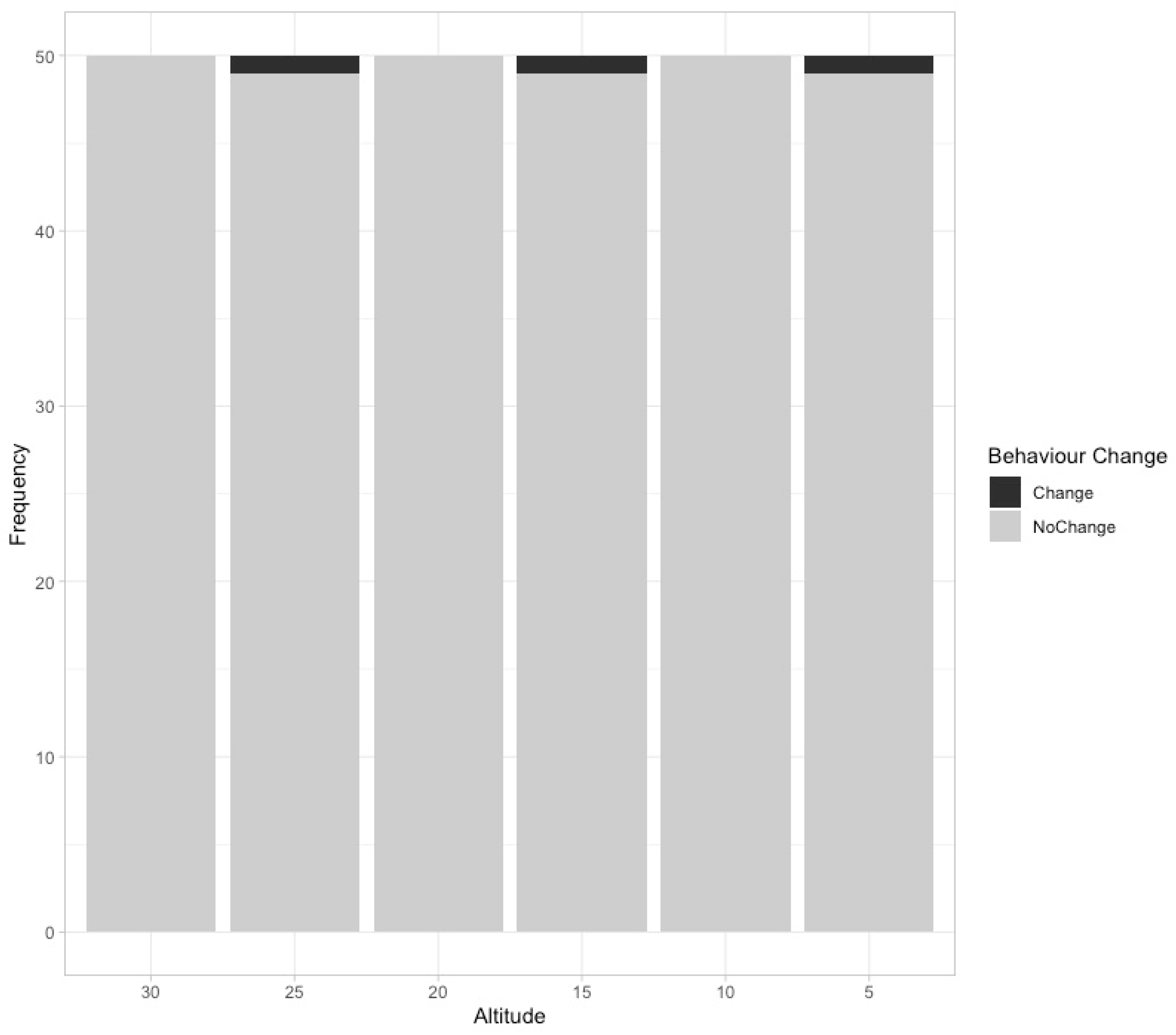
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Oleksyn, S.; Tosetto, L.; Raoult, V.; Williamson, J.E. Drone-Based Tracking of the Fine-Scale Movement of a Coastal Stingray (Bathytoshia brevicaudata). Remote Sens. 2021, 13, 40. [Google Scholar] [CrossRef]
- Schofield, G.; Esteban, N.; Katselidis, K.A.; Hays, G.C. Drones for research on sea turtles and other marine vertebrates—A review. Biol. Conserv. 2019, 238, 108214. [Google Scholar] [CrossRef]
- Pirotta, V.; Hocking, D.P.; Iggleden, J.; Harcourt, R. Drone Observations of Marine Life and Human—Wildlife Interactions off Sydney, Australia. Drones 2022, 6, 75. [Google Scholar] [CrossRef]
- Fiori, L.; Doshi, A.; Martinez, E.; Orams, M.B.; Bollard-Breen, B. The use of unmanned aerial systems in marine mammal research. Remote Sens. 2017, 9, 543. [Google Scholar] [CrossRef] [Green Version]
- Rümmler, M.-C.; Mustafa, O.; Maercker, J.; Peter, H.-U.; Esefeld, J. Measuring the influence of unmanned aerial vehicles on Adélie penguins. Polar Biol. 2016, 39, 1329–1334. [Google Scholar] [CrossRef]
- Ruiz-García, D.; Adams, K.; Brown, H.; Davis, A.R. Determining stingray movement patterns in a wave-swept coastal zone using a blimp for continuous aerial video surveillance. Fishes 2020, 5, 31. [Google Scholar] [CrossRef]
- Dulvy, N.K.; Pacoureau, N.; Rigby, C.L.; Pollom, R.A.; Jabado, R.W.; Ebert, D.A.; Finucci, B.; Pollock, C.M.; Cheok, J.; Derrick, D.H. Overfishing drives over one-third of all sharks and rays toward a global extinction crisis. Curr. Biol. 2021, 31, 4773–4787.e8. [Google Scholar] [CrossRef]
- Pacoureau, N.; Rigby, C.L.; Kyne, P.M.; Sherley, R.B.; Winker, H.; Carlson, J.K.; Fordham, S.V.; Barreto, R.; Fernando, D.; Francis, M.P. Half a century of global decline in oceanic sharks and rays. Nature 2021, 589, 567–571. [Google Scholar] [CrossRef]
- Setyawan, E.; Stevenson, B.C.; Izuan, M.; Constantine, R.; Erdmann, M.V. How Big Is That Manta Ray? A Novel and Non-Invasive Method for Measuring Reef Manta Rays Using Small Drones. Drones 2022, 6, 63. [Google Scholar] [CrossRef]
- Crook, K.A.; Sheaves, M.; Barnett, A. Species-specific foraging behaviors define the functional roles of sympatric stingrays. Limnol. Oceanogr. 2022, 67, 219–230. [Google Scholar] [CrossRef]
- Frixione, M.G.; García, M.D.J.G.; Gauger, M.F. Drone imaging of elasmobranchs: Whale sharks and golden cownose rays co-occurrence in a zooplankton hot-spot in southwestern Sea of Cortez. Food Webs 2020, 24, e00155. [Google Scholar] [CrossRef]
- Ditmer, M.A.; Werden, L.K.; Tanner, J.C.; Vincent, J.B.; Callahan, P.; Iaizzo, P.A.; Laske, T.G.; Garshelis, D.L. Bears habituate to the repeated exposure of a novel stimulus, unmanned aircraft systems. Conserv. Physiol. 2019, 7, coy067. [Google Scholar] [CrossRef] [Green Version]
- Rebolo-Ifrán, N.; Grilli, M.G.; Lambertucci, S.A. Drones as a threat to wildlife: YouTube complements science in providing evidence about their effect. Environ. Conserv. 2019, 46, 205–210. [Google Scholar] [CrossRef]
- Raoult, V.; Colefax, A.P.; Allan, B.M.; Cagnazzi, D.; Castelblanco-Martínez, N.; Ierodiaconou, D.; Johnston, D.W.; Landeo-Yauri, S.; Lyons, M.; Pirotta, V. Operational protocols for the use of drones in marine animal research. Drones 2020, 4, 64. [Google Scholar] [CrossRef]
- Mo, M.; Bonatakis, K. An examination of trends in the growing scientific literature on approaching wildlife with drones. Drone Syst. Appl. 2022, 10, 111–139. [Google Scholar] [CrossRef]
- Frixione, M.G.; Salvadeo, C. Drones, gulls and urbanity: Interaction between new technologies and human subsidized species in coastal areas. Drones 2021, 5, 30. [Google Scholar] [CrossRef]
- McIntosh, R.R.; Holmberg, R.; Dann, P. Looking without landing—Using remote piloted aircraft to monitor fur seal populations without disturbance. Front. Mar. Sci. 2018, 5, 202. [Google Scholar] [CrossRef] [Green Version]
- Christiansen, F.; Nielsen, M.L.; Charlton, C.; Bejder, L.; Madsen, P.T. Southern right whales show no behavioral response to low noise levels from a nearby unmanned aerial vehicle. Mar. Mammal Sci. 2020, 36, 953–963. [Google Scholar] [CrossRef]
- Pomeroy, P.; O’connor, L.; Davies, P. Assessing use of and reaction to unmanned aerial systems in gray and harbor seals during breeding and molt in the UK. J. Unmanned Veh. Syst. 2015, 3, 102–113. [Google Scholar] [CrossRef] [Green Version]
- Giles, A.B.; Butcher, P.A.; Colefax, A.P.; Pagendam, D.E.; Mayjor, M.; Kelaher, B.P. Responses of bottlenose dolphins (Tursiops spp.) to small drones. Aquat. Conserv. Mar. Freshw. Ecosyst. 2021, 31, 677–684. [Google Scholar] [CrossRef]
- Butcher, P.A.; Colefax, A.P.; Gorkin III, R.A.; Kajiura, S.M.; López, N.A.; Mourier, J.; Purcell, C.R.; Skomal, G.B.; Tucker, J.P.; Walsh, A.J. The drone revolution of shark science: A review. Drones 2021, 5, 8. [Google Scholar] [CrossRef]
- Fettermann de Oliveira, T. Unmanned Aerial Vehicle (UAV) Remote Sensing of Behaviour and Habitat Use of the Nationally Endangered Bottlenose Dolphin (Tursiops truncatus) off Great Barrier Island. Ph. D. Thesis, Auckland University of Technology, Auckland, New Zealand, 2018. [Google Scholar]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. arXiv 2014, arXiv:1406.5823. [Google Scholar]
- RStudio Team. RStudio: Integrated Development for R; RStudio, Inc.: Boston, MA, USA, 2015. [Google Scholar]
- R Development Core Team. R: A language and environment for statistical computing. R Foundation for Statistical Computing: Vienna, Austria; European Environment Agency: Copenhagen, Denmark, 2013. [Google Scholar]
- Wilson, K.; Fritz, L.; Kunisch, E.; Chumbley, K.; Johnson, D. Effects of research disturbance on the behavior and abundance of Steller sea lions (Eumetopias jubatus) at two rookeries in Alaska. Mar. Mamm. Sci. 2012, 28, E58–E74. [Google Scholar] [CrossRef]
- Ramos, E.A.; Maloney, B.; Magnasco, M.O.; Reiss, D. Bottlenose dolphins and antillean manatees respond to small multi-rotor unmanned aerial systems. Front. Mar. Sci. 2018, 5, 316. [Google Scholar] [CrossRef]
- Bevan, E.; Whiting, S.; Tucker, T.; Guinea, M.; Raith, A.; Douglas, R. Measuring behavioral responses of sea turtles, saltwater crocodiles, and crested terns to drone disturbance to define ethical operating thresholds. PLoS ONE 2018, 13, e0194460. [Google Scholar] [CrossRef] [PubMed]
- Durban, J.W.; Fearnbach, H.; Barrett-Lennard, L.; Perryman, W.; Leroi, D. Photogrammetry of killer whales using a small hexacopter launched at sea. J. Unmanned Veh. Syst. 2015, 3, 131–135. [Google Scholar] [CrossRef]
- Acevedo-Whitehouse, K.; Rocha-Gosselin, A.; Gendron, D. A novel non-invasive tool for disease surveillance of free-ranging whales and its relevance to conservation programs. Anim. Conserv. 2010, 13, 217–225. [Google Scholar] [CrossRef]
- Tablado, Z.; Jenni, L. Determinants of uncertainty in wildlife responses to human disturbance. Biol. Rev. 2017, 92, 216–233. [Google Scholar] [CrossRef]
- Stankowich, T.; Blumstein, D.T. Fear in animals: A meta-analysis and review of risk assessment. Proc. R. Soc. B Biol. Sci. 2005, 272, 2627–2634. [Google Scholar] [CrossRef] [Green Version]
- Mulero-Pázmány, M.; Jenni-Eiermann, S.; Strebel, N.; Sattler, T.; Negro, J.J.; Tablado, Z. Unmanned aircraft systems as a new source of disturbance for wildlife: A systematic review. PLoS ONE 2017, 12, e0178448. [Google Scholar] [CrossRef] [Green Version]
- Cooper, W.E., Jr. Factors affecting risk and cost of escape by the broad-headed skink (Eumeces laticeps): Predator speed, directness of approach, and female presence. Herpetologica 1997, 53, 464–474. [Google Scholar]
- Bateman, P.; Fleming, P. Who are you looking at? Hadeda ibises use direction of gaze, head orientation and approach speed in their risk assessment of a potential predator. J. Zool. 2011, 285, 316–323. [Google Scholar] [CrossRef]
- Smith, C.E.; Sykora-Bodie, S.T.; Bloodworth, B.; Pack, S.M.; Spradlin, T.R.; LeBoeuf, N.R. Assessment of known impacts of unmanned aerial systems (UAS) on marine mammals: Data gaps and recommendations for researchers in the United States. J. Unmanned Veh. Syst. 2016, 4, 31–44. [Google Scholar] [CrossRef] [Green Version]
- Sills, J.M.; Reichmuth, C.; Southall, B.L.; Whiting, A.; Goodwin, J. Auditory biology of bearded seals (Erignathus barbatus). Polar Biol. 2020, 43, 1681–1691. [Google Scholar] [CrossRef]
- Chapuis, L.; Collin, S.P. The auditory system of cartilaginous fishes. Rev. Fish Biol. Fish. 2022, 32, 521–554. [Google Scholar] [CrossRef]
- Oleksyn, S.; Tosetto, L.; Raoult, V.; Joyce, K.E.; Williamson, J.E. Going Batty: The Challenges and Opportunities of Using Drones to Monitor the Behaviour and Habitat Use of Rays. Drones 2021, 5, 12. [Google Scholar] [CrossRef]
- Ditmer, M.A.; Vincent, J.B.; Werden, L.K.; Tanner, J.C.; Laske, T.G.; Iaizzo, P.A.; Garshelis, D.L.; Fieberg, J.R. Bears show a physiological but limited behavioral response to unmanned aerial vehicles. Curr. Biol. 2015, 25, 2278–2283. [Google Scholar] [CrossRef] [Green Version]
- Wolf, I.D.; Croft, D.B. Minimizing disturbance to wildlife by tourists approaching on foot or in a car: A study of kangaroos in the Australian rangelands. Appl. Anim. Behav. Sci. 2010, 126, 75–84. [Google Scholar] [CrossRef]
- Lambert, F.N.; Treberg, J.R.; Anderson, W.G.; Brandt, C.; Evans, A.N. The physiological stress response of the Atlantic stingray (Hypanus sabinus) to aerial exposure. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2018, 219, 38–43. [Google Scholar] [CrossRef]
- Cicia, A.M.; Schlenker, L.S.; Sulikowski, J.A.; Mandelman, J.W. Seasonal variations in the physiological stress response to discrete bouts of aerial exposure in the little skate, Leucoraja erinacea. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2012, 162, 130–138. [Google Scholar] [CrossRef]
- Prohaska, B.K.; Bethea, D.M.; Poulakis, G.R.; Scharer, R.M.; Knotek, R.; Carlson, J.K.; Grubbs, R.D. Physiological stress in the smalltooth sawfish: Effects of ontogeny, capture method, and habitat quality. Endanger. Species Res. 2018, 36, 121–135. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Behaviour | Definition |
|---|---|
| Swimming | Moving through the water |
| Foraging | Looking or searching for food or provisions/ingesting food; flapping pectoral fins to excavate food off seafloor |
| Resting | Lying motionless on the substrate |
| Altitude (m) | Number of Individuals Exhibiting Behaviour | ||
|---|---|---|---|
| Swimming | Foraging | Resting | |
| 30 | 27 | 15 | 8 |
| 25 | 25 | 17 | 8 |
| 20 | 25 | 17 | 8 |
| 15 | 25 | 17 | 8 |
| 10 | 23 | 19 | 8 |
| 5 | 19 | 23 | 8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bourke, E.; Raoult, V.; Williamson, J.E.; Gaston, T.F. Estuary Stingray (Dasyatis fluviorum) Behaviour Does Not Change in Response to Drone Altitude. Drones 2023, 7, 164. https://doi.org/10.3390/drones7030164
Bourke E, Raoult V, Williamson JE, Gaston TF. Estuary Stingray (Dasyatis fluviorum) Behaviour Does Not Change in Response to Drone Altitude. Drones. 2023; 7(3):164. https://doi.org/10.3390/drones7030164
Chicago/Turabian StyleBourke, Emily, Vincent Raoult, Jane E. Williamson, and Troy F. Gaston. 2023. "Estuary Stingray (Dasyatis fluviorum) Behaviour Does Not Change in Response to Drone Altitude" Drones 7, no. 3: 164. https://doi.org/10.3390/drones7030164