1. Introduction
Incorporating legumes into grass-dominated pastures can have a number of agronomically important benefits. Increasing species diversity by introducing legumes to grasslands was shown to increase dry matter and protein yield compared to monocultures or grass-dominated pastures [
1,
2,
3,
4,
5,
6,
7,
8]. Furthermore, legumes’ ability to symbiotically fix nitrogen and reduce the need for fossil-fuel-based mineral N-fertilizer has gained increasing attention. Mineral and organic fertilizer applications also encompass the emission of greenhouse-gas-relevant gases such as N
2O or NH
3, which is much less the case when symbiotically fixed N by legumes is used in the system. The carbon footprint of intensively managed grassland could be reduced considerably by making use of those features. Recently, the drought tolerance of legumes has gained attention. Legumes were reported to sustain growth under restricted drought conditions in contrast to a number of highly productive grass species [
9,
10,
11]. This makes legumes a promising component for grasslands under changing climatic conditions, which include increased risks of drought conditions even in temperate regions.
Due to climate change, weather conditions in the future are predicted to be more variable [
12,
13], which also entails more variable rainfall distributions. This can lead to the more frequent occurrence of drought conditions differing in intensity and timing. The actual drought stress on plants is difficult to determine [
14], partly because it depends on numerous climatic factors such as radiation, air temperature, and wind, soil-based factors such as soil depth and soil water holding capacity, and plant-based traits such as water use efficiency and the evapotranspiration rate, which also includes factors such as the actual rooting depth. A lack of rainfall can thus affect grassland in many ways, depending on the season, length, and severity of drought, the soil, and the botanical composition of the grassland at the time. Although legumes were mentioned to be productive under moderate drought conditions [
9,
11], the same was stated for some drought-tolerant grasses [
15,
16].
In general, the effect of water relations on clover growth was mentioned to be contradicting [
17]. On the one hand, legumes have high rates of transpiration relative to grasses [
18], but then again, they also show higher water use efficiencies than non-legumes [
19]. Contradictions may also arise because the growth benefit of legumes was mentioned to be quite different under moderate compared to severe drought conditions [
20], keeping in mind that the severity of drought for plants is difficult to measure. Therefore, it remains unclear whether drought-tolerant grasses or legumes, or a mix of both, would be a reliable safeguard in terms of securing dry matter (DM) and protein productivity from grasslands under natural rainfall conditions.
Compositions of grasslands are not fixed and change due to management, environmental conditions, and the competitive abilities of neighboring plants, which in turn can be strongly affected by N fertilizer applications. The external N requirements of grasses and legumes are quite different, and the competitive advantages of either plant group depend on it. The botanical composition of mixed grasslands usually changes according to their N-input, with higher N-rates generally favoring grasses compared to legumes [
21,
22]. Trials are usually conducted at a fixed rate of N fertilizer application. Depending on the particular N-rate, this will favor grasses (high N-rate) or legumes (low N-rate), but generally not both groups in a similar way. Furthermore, with a standard N fertilizer input to mixed grasslands, the additional N-input of legumes due to N
2 fixation is not taken into account when comparing treatments. As a result, the absolute N-input for mixed grassland systems can vary considerably due to their respective legume content. Comparing the productivity of mixed grass-legume grasslands on the basis of an equal N-supply and thus equal growth potential would require taking the N-input of legumes into account.
The legume component of intensively managed, mixed grasslands in moderate climates of northern Europe (i.e., Germany) usually consists of red or white clover. They have been differentiated due to their rooting depth [
1,
2] but they differ also in additional traits. White clover is known to have a higher N-fixing potential than RC [
23,
24], and at the same time, due to the creeping growth habit, is considered less competitive than RC [
3]. The differing competitive abilities of the two legume species could be further altered by N–fertilizer additions, which may not affect the two legumes in a similar way.
The aim of the study was to assess the benefit of legumes compared to a mix of drought-tolerant grasses in a mixed grassland with Lolium perenne L. under naturally occurring drought conditions. For this purpose, variable seeding mixtures of four groupings were seeded, resulting in a wide range of legume percentages in mixed grasslands. The four groups or species were (a) L. perenne L., (b) a mix of drought-tolerant grasses, (c) T. repens, and (d) T. pratense. The trial was conducted under two different N-fertilizer regimes. One set of legume–grass mixtures received a fixed N-fertilizer rate of 180 kg N ha−1. Another set of legume-grass mixtures received an adapted, thus partly reduced, N-fertilizer rate that took the assumed N-fixation rate from legumes into account.
The trial addressed the following questions: (1) Is the improved yield performance of mixed grass–legume grasslands compared to grass monocultures primarily based on the legume-based additional N-input to the system? (2) Is there a different response of T. repens and T. pratense to N fertilizer applications? (3) Do mixed grasslands with legumes perform better under variable drought conditions than grass-dominated grasslands that include drought-tolerant grass species? (4) Is there a different response of T. repens and T. pratense to drought?
2. Materials and Methods
The trial site was a sandy loam with a pH of 5.3. The soil nutrient concentrations in 2019 were 8.3 mg P 100 g
−1 (CAL soluble) and 11.6 mg K 100 g
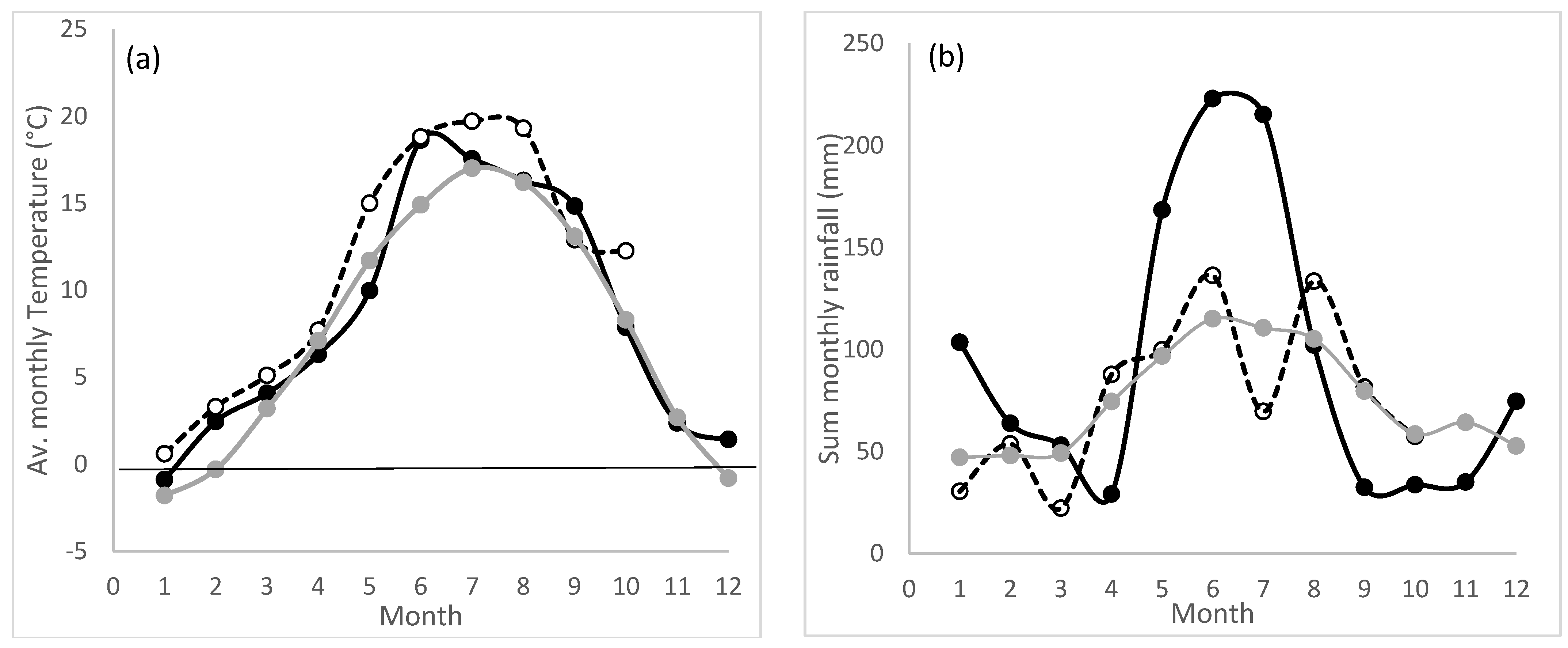
−1 (CAL soluble). The mean annual temperature and sum of annual rainfall during the trial period, as well as the average values for the period of 1960–1990, are shown in
Figure 1. The daily climate water balances, mean daily temperature and daily rainfall for 2021 and 2022 are provided in the
Supplement Table S1. Annual base fertilization of 44 kg P ha
−1 and 166 kg K ha
−1 including 7% sulphur were applied annually in spring. The ICL granulated PKpluS 12/24 (+2MgO +7S +14CaO) fertilizer, based on Polysulfat Technology, was used for fertilization.
The four groupings of (a)
T. pratense (RC), (b)
T. repens (WC), (c)
L. perenne (Lol), and (d) the GSWT-based mixture of grasses (GSWT) were seeded according to simplex designs described in [
1]. In short, monocultures of each group, as well as mixtures in percent of each group of 40-40-10-10, 70-10-10-10, and 25-25-25-25, were used. The resulting mixtures are listed in
Table 1. The GSWT seed mix is recommended for grassland (G) that is cut (S) or grazed (W) and located in the drought-prone areas (T) of Baden Württemberg. The GSWT-based mixture used in this study, termed GSWT, consisted of
Festuca pratensis (30%),
Phleum pratense (30%),
Poa pratensis (20%),
Dactylis glomerata (10%), and
Festuca rubra (10%). The original GSWT mix also contains
L. perenne and
T. repens, which were not included in our GSWT mix. For each RC, WC, and Lol, a mixture of two varieties was seeded. For RC, the varieties Milvus and Merula were used, for WC, the varieties Merlyn and Vysocan were used, and for Lol, the varieties Montova and Arvicola were used. The trial was seeded with a slot seeder on 19 May 2020. The previous crop was lucerne.
The N-treatments consisted of a fixed rate of 180 kg N ha
−1 and an adapted N-rate of 0, 20, 100, or 180 kg N ha
−1 according to the expected percentages of legumes to be established (
Table 1). The required N input, applied as calcium ammonium nitrate, was calculated based on the general N requirement of 240 kg N ha
−1 for a 5-cut meadow in the region subtracting the potential soil N mineralization of 60 kg N ha
−1 due to the previous lucerne crop. The resulting fixed N rate was 180 kg N ha
−1. The potential N
2-fixation of RC and WC were estimated according to models by [
23] (RC-Nfix = 0.026 × DM + 7; WC-Nfix = 0.031 × DM + 24) and are listed in
Table 1. Nitrogen rates for the adapted N-fertilization were calculated by subtracting the estimated N fixation from the fixed N rate of 180 kg N ha
−1 (
Table 1). The 30 treatments were arranged in a completely randomized block design. The harvests were conducted on 15 May 2021, 23 June 2021, 26 July 2021, 1 September 2021, and 19 October 2021 and 19 May 2022, 20 June 2022, 14 July 2022, 8 August 2022 and 6 October 2022 for 2021 and 2022, respectively. The N-rates were applied prior harvests 1, 2, 3, and 4. When 180 kg N ha
−1 were applied, they were split into 60-40-40-40 kg N ha
−1 before harvests 1–4, respectively. When 100 kg N ha
−1 were applied, they were split into 50-10-40-0 kg N ha
−1, and when 20 kg N ha
−1 were applied, they were split into 20-0-0-0 kg N ha
−1 before harvests 1–4, respectively. Nitrogen fertilization treatments started in 2021. In the year of establishment (2020), reduced N rates were applied, with 20 kg N ha
−1 for both fixed and adapted N-treatments with a grass content of ≥80% in the seeding mixture.
In early spring 2021 and in summer 2022, some low rainfall periods occurred (
Figure 1). In spring 2021, only 28 mm of rainfall were recorded in the 39 days between 21.3.21 and 29.4.21. In summer 2022, only 77 mm was recorded for the 44 days between 5.7.22 and 17.8.2022. The latter low rainfall period was accompanied by higher-than-average mean monthly temperatures in July and August. Values were compared to the long-term mean monthly temperatures measured between 1960 and 1990. The climatic water balances (CWB) for the period prior to each harvest were −35.5, 41.3, 150.0, 29.1, −61.4 and −14.1, −37.1, −25.3, −56.2, 35.8 for harvests one to five in 2021 and 2022, respectively. Values from the first of March onwards were used for calculating CWB for harvest 1. Details are provided in
Supplement Table S1.
Statistical analysis was conducted using R (Version 4.1.3, R development Core Team 2016, Vienna, Austria). Polynomial models were fitted using the lm function including linear and quadratic terms separately for annual values of the fixed and adapted fertilizer regimes and tested for significance of the factors. The effect of the N-fertilization regime on DM production was analyzed by comparing polynomial models for the whole dataset per year, with and without the term “fertilizer regime” fitted. Differences between those two models were analyzed by ANOVA.
3. Results
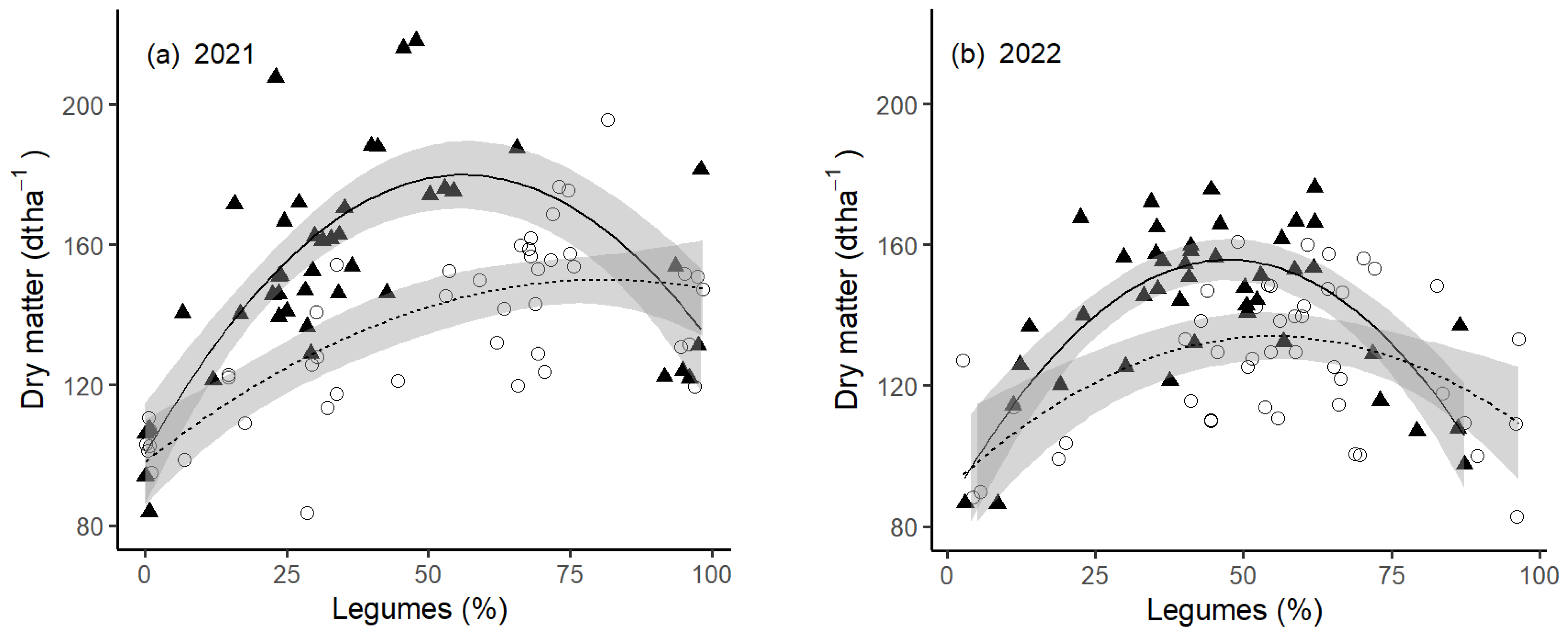
The total DM yield for all treatments across the two years is shown in
Figure 2, and the statistical analysis is shown in
Table 2. In both a comparably humid (2021) and a drought-affected year (2022), the yield of mixed-species grasslands with legumes was higher than monocultures of either grasses or legumes. This was at least the case under a fixed fertilizer regime (180 kg N ha
−1) for both years, where fitted models primarily vary in the intercept, being lower in the drought-prone year.
Treatments under the adapted-N fertilizer regime generally showed significantly lower DM production compared to treatments under a fixed fertilizer regime in both years. Nevertheless, even under an adapted fertilizer regime, the mixed-species grasslands showed increased yield compared to monocultures, although the extent was considerably lower, as the factors (x, x2) of the model suggest. Furthermore, this beneficial effect of mixed-species grassland on DM yield was more obvious in the drought-affected year than in the humid year. In the humid year, DM production increased almost linearly with legume content in the grassland, with only a small negative x2 term in the polynomial model.
The total DM yield for each treatment across the two years is shown in
Table 3. The highest DM production for the two years was gained by RC- and GSWT-dominated mixtures under a fixed fertilization scheme. The DM yield of treatments receiving an adapted- compared to fixed-N fertilizer application was generally lower but there were many exceptions. One exception were legume monocultures (RC, WC). Exceptions for mixed-species treatments were particularly observed in treatments with a high percentage of WC in the seeding mixture (40Lol-40WC; 25_25, 40GSWT 40WC), but considerably less so for seeding mixtures dominated by RC (40GSWT 40RC, 70RC).
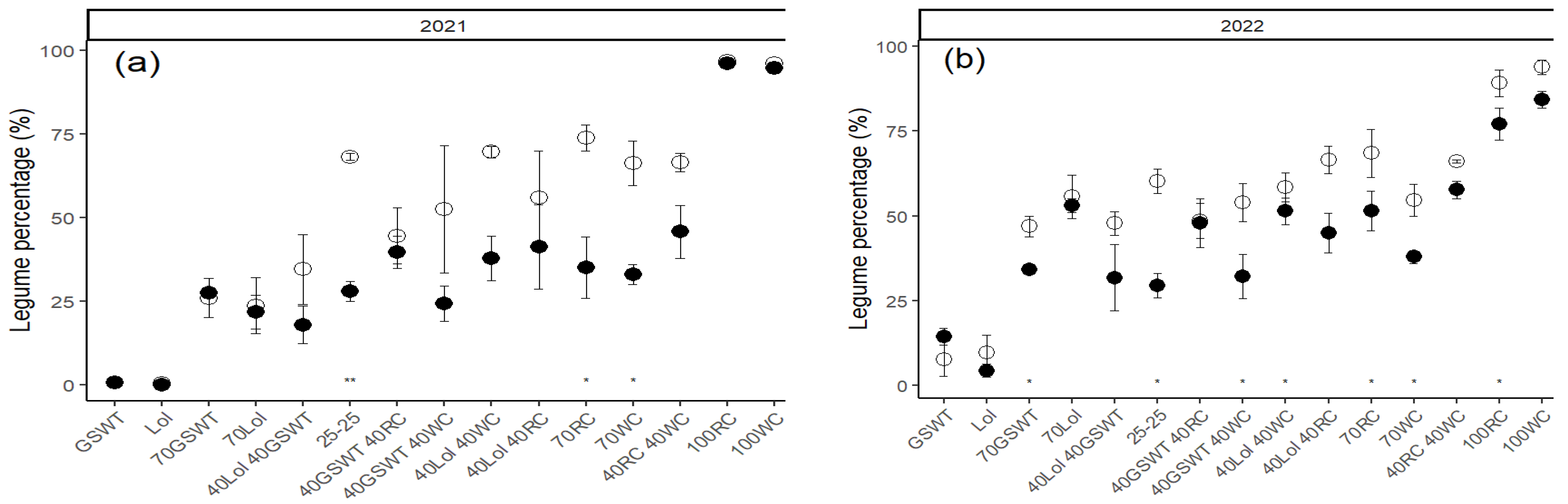
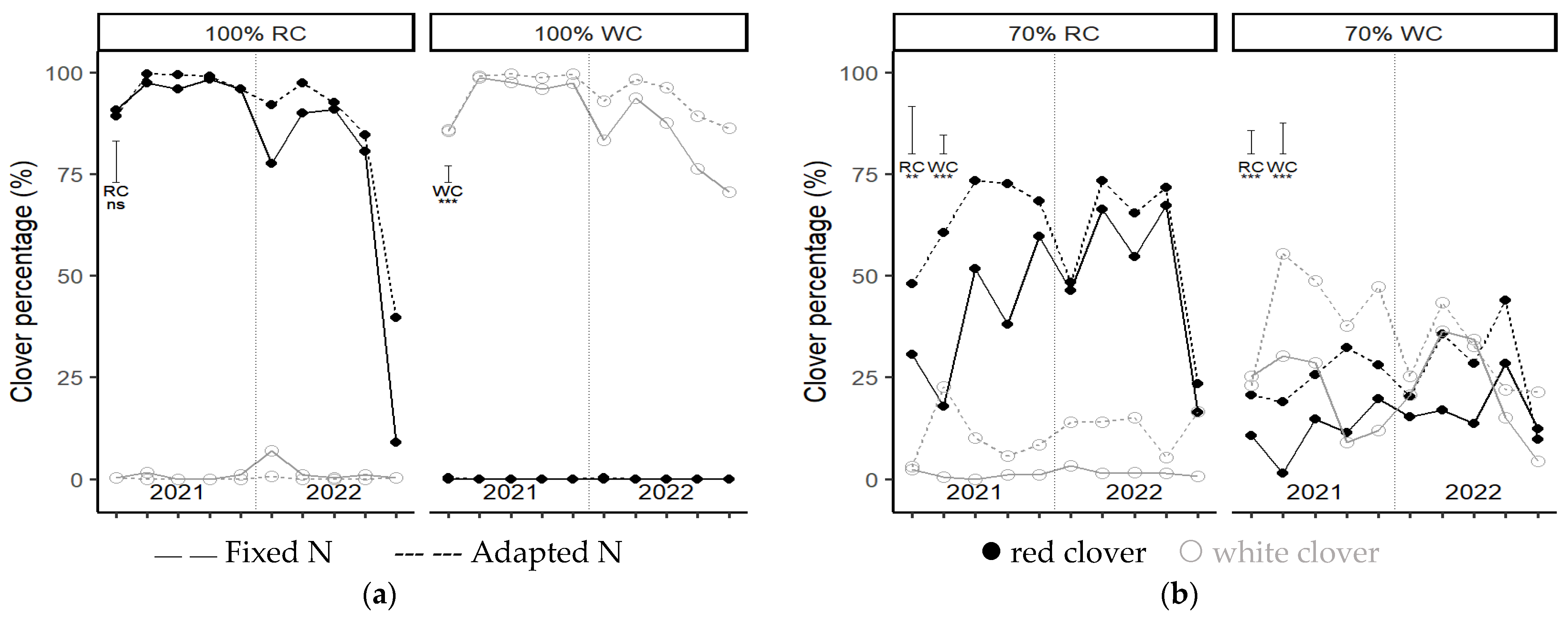
The fertilizer regime also affected the legume percentage in most mixed grass–legumes treatments. The weighted average legume content for both years is shown in
Figure 3. The legume content in mixed grasslands was most often significantly reduced due to the fixed- compared to the adapted-N fertilizer application. The legume content was generally reduced by 15–20% and occasionally by 41%. Nitrogen additions tended to reduce the legume percentage in WC-dominated seeding mixtures more than in RC-dominated seeding mixtures.
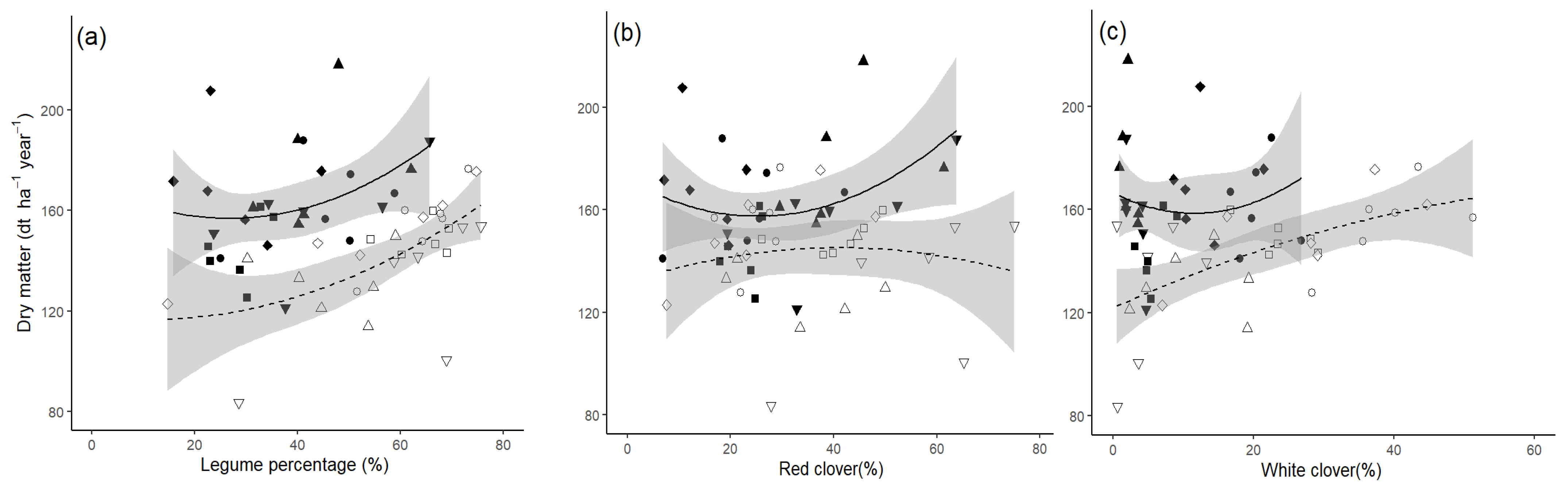
Potential differences between WC and RC in terms of the susceptibility to the fertilizer regime or DM productivity become obvious when only treatments with a seeding mixture of 50% legumes are considered.
Figure 4 shows the established percentage of legumes and the percentage of RC, WC, and their relation to DM yield under fixed or adapted-N fertilizer applications for 50% legume seeding mixtures.
Table 4 shows the statistical analysis of the models.
Considering estimated legume percentages (RC plus WC) only, it appears that legumes were well established under fixed and adapted fertilizer regimes (x-values). However, the x-values for the two species separately show that WC sustained percentages only well below 24% under fixed-N fertilization, but up to 52% in the adapted-N fertilizer regime, whereas RC established legume percentages (x-values) almost equally well under both N regimes. Similarly, an increase in the legume percentage (RC plus WC) appears to be correlated with increased yield under the adapted N fertilizer regime (y-values). However, values for the two species separately show that only the WC content is positively correlated with DM yield (y-values), while the RC content is not correlated with yield increases. Under the fixed-N fertilizer regimes, no positive relationship between legume percentages and DM yield could be observed for either species.
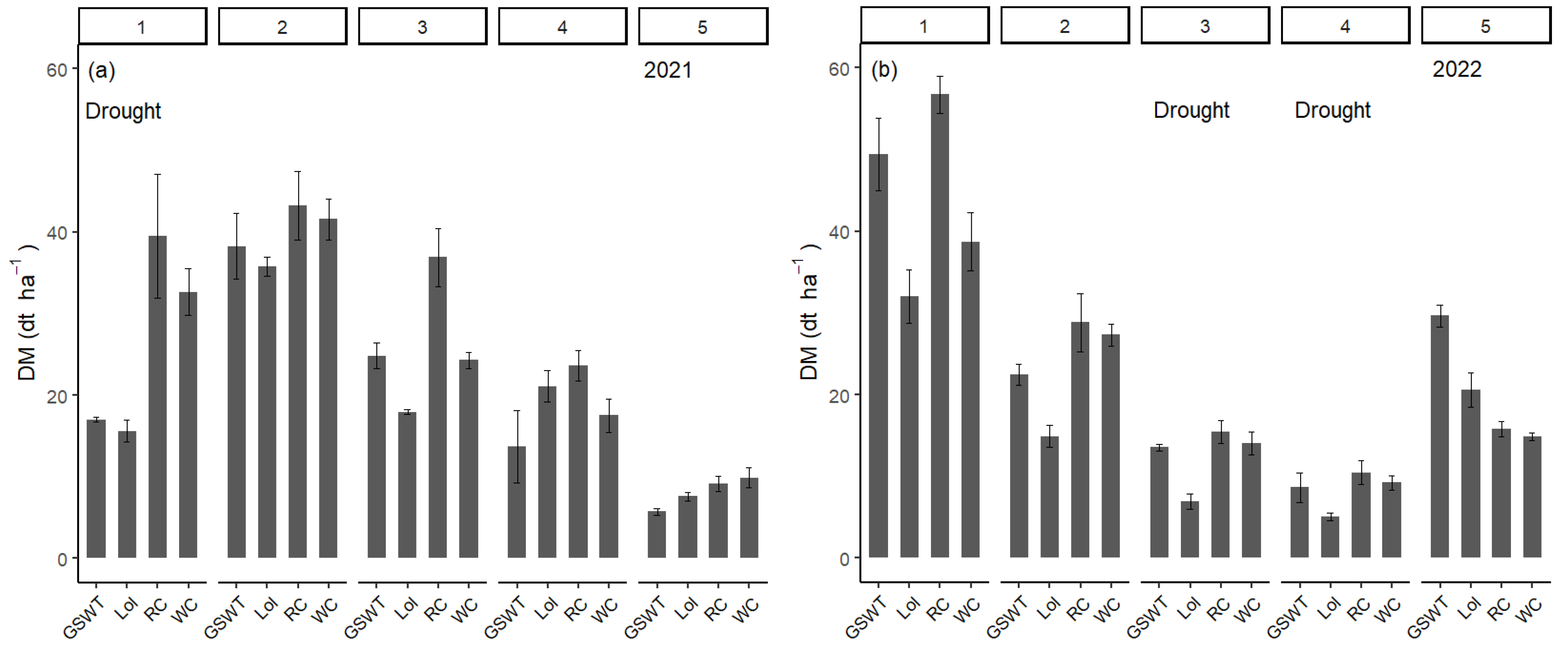
Low-rainfall conditions occurred prior harvest 1 in 2021 and prior harvests 3 and 4 in 2022. The DM production of RC, WC, Lol, and GSWT monocultures for each harvest in both years is shown in
Figure 5. In all drought periods, the DM production of both legume species was significantly higher than Lol. The growth comparison to GSWT is less clear. During the moderate spring drought in 2021, legume growth was significantly higher than from GSWT, whereas, during the pronounced summer drought in 2022, the growth of RC, WC, and GSWT monocultures was equally high. By this time, the GSWT plots were dominated by
D. glomerata. In the post-drought period in 2022, the DM production of particularly GSWT and also Lol plots was significantly higher than RC and WC plots. Grasses showing signs of post-drought compensation with similar tendencies being obvious in harvest 2 in 2021 following the spring drought. In drought-affected harvests the dry matter production, although enhanced in RC, WC, and GSWT plots, was relatively low compared to the DM yield of the remaining harvests. Dry matter production in harvests 1 and 2 and even in the post-drought periods was more substantial and contributed significantly more to the annual yield than DM yield during drought periods.
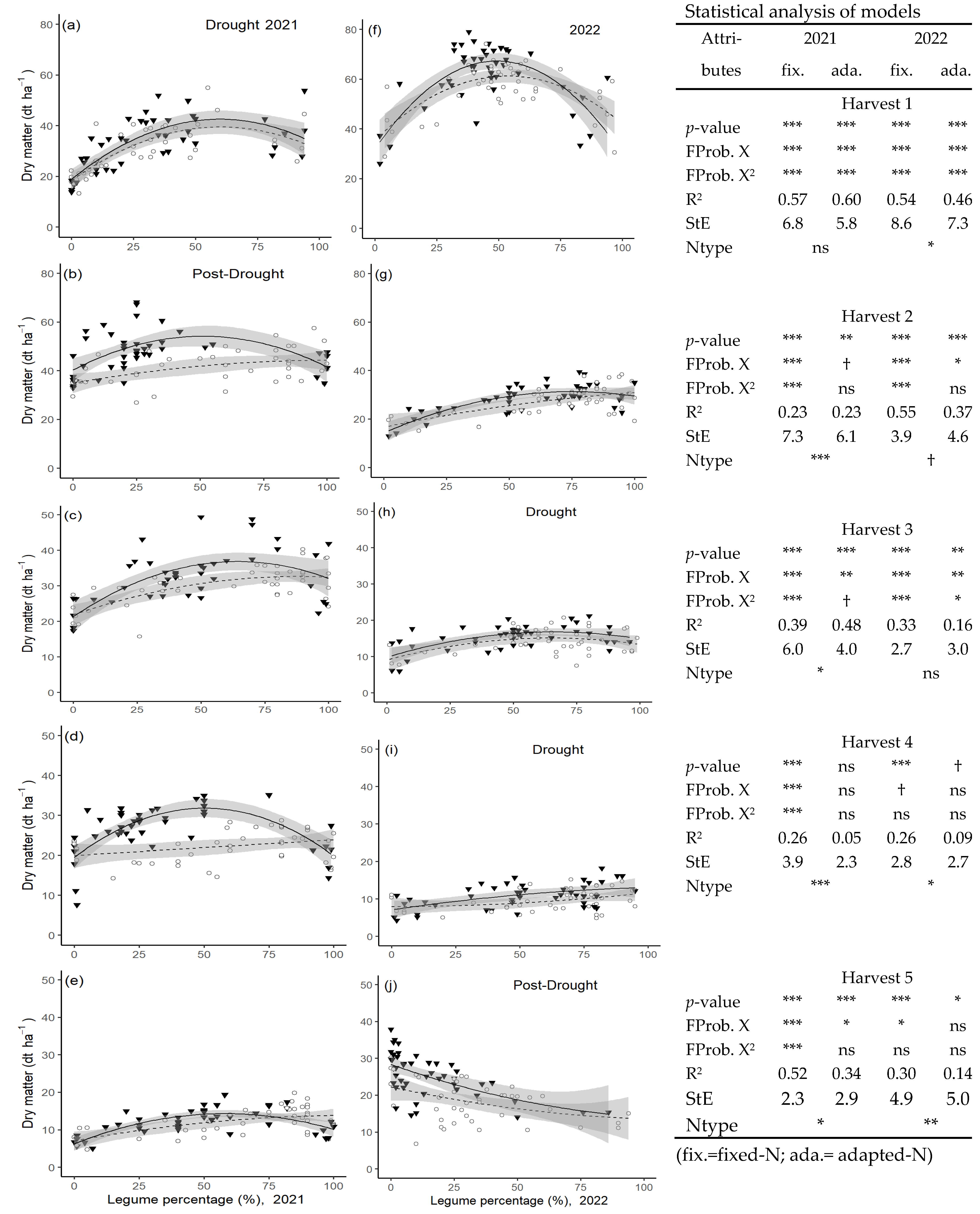
The effect of species diversity (legume content), fertilizer regime and, their interaction with drought is shown in
Figure 6. The DM yield for each harvest in 2021 and 2022 and the statistical analysis of the models are shown. When low-rainfall periods occurred prior to harvests, it is indicated in the graph. The scale of the Y-axis varies for harvests 1 and 2 compared to 3, 4, and 5.
During moderate drought periods (harv.1 2021, harv.3 2022), mixed-species swards showed increased yields compared to monocultures, and this was under fixed and adapted fertilizer regimes. At the same time, no yield difference due to the fertilizer regime could be observed. When the drought intensified (harv.4, 2022), the beneficial effect of the mixed-species sward could not be measured anymore, and only small yield differences occurred at all. In the post-drought period in 2022, significant yield differences were measurable, but the yield was positively correlated with grass content and negatively correlated with the legume content of the plots. No yield advantage for mixed-species plots could be measured in this post-drought period.
Under humid conditions (harv.2–5 2021), the yield benefit of mixed-species grassland was measurable in each harvest, but only under a fixed-N fertilizer regime. Under an adapted fertilizer application, DM production remained on a similar level, irrespective of the legume content. Under those humid conditions, the effect of the fertilizer regime was highly significant. The only exception was harvest 1 in 2022, in the second year of the experiment, when only one out of four fertilizer applications for the season had been applied by then. At this point mixed-species effects were measured both under fixed and adapted fertilizer regimes.
In the post-drought period in autumn 2022, the RC percentages in grasslands were significantly reduced compared to the previous harvests (
Figure 7), and considerable RC losses occurred in monocultures as well as in mixed grasslands. The RC loss was enhanced in the fixed compared to the adapted-N fertilized plots, with considerable variation within treatments. The WC percentages in grassland were also reduced in the post-drought period, but much less so than RC percentages.
4. Discussion
Incorporating legume species into grass-dominated grasslands increased the DM yield both in a relatively humid and a relatively dry year with naturally occurring low-rainfall conditions. This agrees with findings gained under more average weather conditions [
1,
3,
4,
5] and under artificially imposed drought conditions [
9,
10]. This so-called “transgressive overyielding” of higher DM yield of mixed compared to the highest-yielding monocultures [
3] was also observed under a fixed or adapted N-fertilizer application, but the effects were much more pronounced under the fixed N-fertilizer regime.
The estimated N-input in the fixed-N fertilizer regime varied considerably and ranged from 240 up to 480 kg N ha
−1 including mineral N-fertilizer, N
2-fixation by legumes, and soil and crop residue N-mineralization (
Table 1). Under the adapted N fertilizer regime, the N-input was generally estimated to be similar across all treatments (240 kg N ha
−1) with few exceptions (legume content 100%). A positive species diversity effect in the adapted fertilizer regime occurred in the dry year but was much less obvious in the humid year.
Yield increases went up almost linearly with legume content in the humid year under the adapted fertilizer regime. This suggests that the additional N-input in the system due to N
2 fixation, rather than the species diversity effect, may then have been the major factor allowing for yield increases in mixed grasslands. Indeed, legumes were mentioned to be particularly productive in humid conditions [
25], provided competition from grass is low. In the humid periods, the effect of the N-fertilizer regime was generally highly significant again, suggesting that N-input, whether by fertilizer or due to N
2 fixation, is the decisive growth factor.
Legumes in mixed pastures are generally discussed as one group, although there are major differences between the two predominant clover species in intense grassland [
26,
27,
28]. The higher N
2-fixing potential of WC compared to RC has been mentioned previously [
23], as well as the higher N transfer, as a percentage of the total N in WC, compared to RC [
26]. The lack of yield reductions of WC-rich swards under the adapted compared to the fixed N-fertilizer regime supports these findings. When N-fertilizer rates are reduced, accompanied grasses are increasingly dependent on direct or indirect N-transfer from adjacent legumes. Limited N-transfer from legumes in RC-dominated mixtures was likely the reason for yield reductions when N applications were reduced in those RC- mixed grasslands.
The two legume species, WC and RC, also differed in terms of their tolerance towards elevated N-fertilizer applications, which is in line with previous findings [
29]. In general, legume percentages were reduced by 15–20% and up to 41% in the fixed compared to the adapted N-fertilizer treatments (
Figure 3), but this affected the two legume species differently. Starting from 50% legumes in the seeding mixture (
Figure 4), WC percentages reached levels up to 52% under an adapted regime but were reduced to percentages of below 22% under a fixed N fertilization regime, whereas RC percentages changed comparably little due to N fertilization. However, only when WC percentages could reach levels above 20% (adapted N-fertilisation) the yield increases were substantial enough to allow for similar DM yields of the fixed- and adapted N fertilizer treatments. Legume percentage and N fertilization rate are negatively correlated and not all combinations of those two factors are possible.
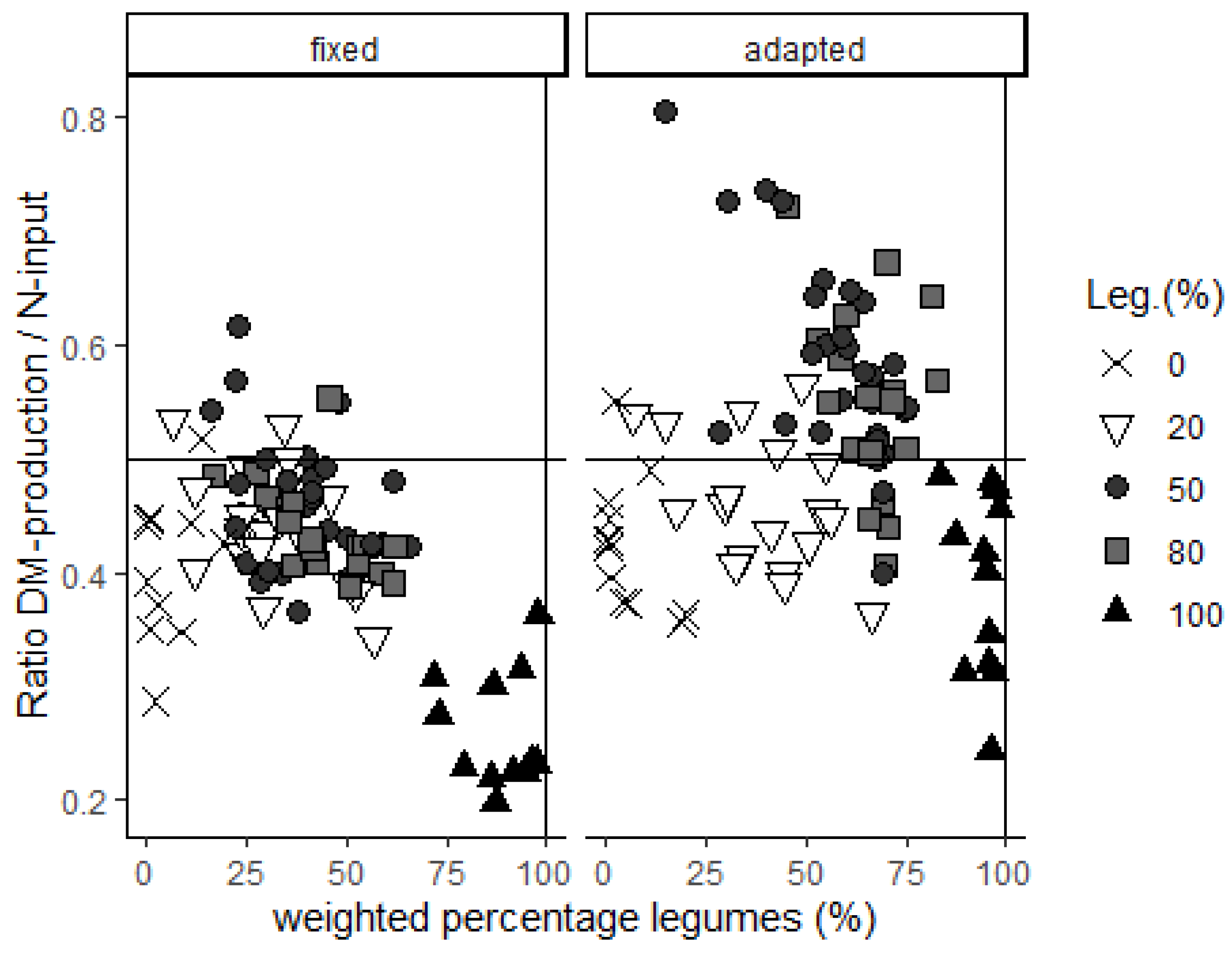
Since N-application rates change the percentage of legumes and even the predominant legume type in mixed swards, those rates determine if the highest DM output with high N-input or the best balance between DM-output/N-input is achieved. Nowadays, the optimum use of N-fertilizer is warranted since this can save on monetary costs and production-based CO
2 emissions. Nitrogen use is optimized when the ratio of TM output to N input is highest. This ratio was generally enhanced in mixtures containing a legume percentage between 50 and 80% with an adapted N-fertilizer regime, although considerable variation existed (
Figure 8).
During moderate and severe drought periods, the yield of legume monocultures was significantly higher than from Lol but on a similar level as GSWT grasses, at least during the summer of 2022. The GSWT mix was dominated by
D. glomerata by then.
Dactylis glomerata roots have the ability to extract water at low soil water potentials, which contributes to their survival under severe drought [
30] more so than other drought-tolerant grasses. Even then, the DM production of both legume species was equally high. During the moderate spring drought in 2021, legumes showed an improved DM yield compared to both Lol and the GSWT mix. Growth advantages for legumes during that time were likely to be based on reduced soil N mineralization during drought combined with a higher susceptibility of
D. glomerata to winter compared to summer droughts [
31].
Including legumes in mixed grasslands in our study increased grassland yield under a moderate spring and a moderate summer drought. During these drought periods, mixed grasslands showed higher yields than monocultures and no yield differences were observed between treatments under fixed and adapted N-fertilizer regimes. Under moderate drought conditions, fertilizer-derived N could enhance yield only to a limited level. Under those conditions, productivity was dependent on legume-derived N and the fertilizer regime had little influence. This could indicate that the positive effect of legumes under drought also seems to be based on their ability for N2 fixation and less so on other attributes. When drought intensified, the positive effect of legumes on the productivity of mixed swards was negligible, because water was the restricting factor by then.
Although legumes proved to be beneficial during drought, absolute yield gains during this time were low. Growth is largely the physical response of cells to turgor, and this elucidates that the opportunity for plants to continue growth in the face of severe water deficit is limited [
14]. Deep rooting and access to water in lower soil horizons would be an alternative strategy to keep up growth during drought [
20]. Red and WC are considered deep and shallow rooting legumes, respectively [
9]. Nevertheless, an advantage of RC in terms of DM production during moderate and severe droughts compared to WC could not be observed in our trials. In contrast, RC losses after drought were more pronounced than WC losses (see below).
Annual DM production includes regrowth in the post-drought period, where the recovery of grasses was striking following the severe drought (2022), and Lol and particularly GSWT growth outperformed legume growth by far. This is in agreement with studies that suggested a strong overcompensation after drought for a number of grass species [
16], and similarly, a slow recovery of legumes after prolonged drought periods [
9,
10]. However, the low DM production of legumes in the post-drought period was not only based on a slow recovery of plants.
The considerable decline of RC in particular and less so of WC was measured in the post-drought period, which was further intensified by enhanced N-fertilizer application. Competition from grasses was unlikely to be the reason in this case because legume reductions also occurred in legume monocultures. Although this would suggest that legumes are prone to plant losses after a severe drought, this was not the case in a parallel experiment at a similarly drought-prone location close by (data not shown). The parallel experiment consisted of clover reseeded permanent grasslands (85 kg N ha−1), where RC percentages remained on a similar level, at 60% during the drought and 50% in the post-drought period. In the current experiment, RC was reduced from 70 to 20% (70% RC, 20 kg N ha−1) from the drought to the post-drought period.
Harvesting time and frequency after drought were the major differences between the two studies. In the current experiment, the harvest occurred right after the drought period, with only a four-week growth period, whereas in the parallel experiment, no harvest occurred right after the drought, and a seven-week growth period was allowed between cuts. Red clover is known to be susceptible to frequent defoliation [
25,
27], possibly because RC is dependent on accumulating carbohydrate reserves in the taproot, which ensures regrowth after harvesting [
28]. Low root carbohydrate reserves due to drought and short cutting intervals, an interaction between management and the environment, were likely to be the reason for the significant RC decline. Management of RC-rich grassland in post-drought periods needs to be adapted accordingly to sustain legumes in mixed pastures.
5. Conclusions
Legume additions to grassland proved to be a favorable option in two quite contrasting years with yield increases being measured in both a relatively humid (2021) and a relatively dry year (2022). The positive effect of including legumes in grassland was primarily, but not solely, based on their ability for symbiotic nitrogen fixation. The species diversity effect appears to be significantly dependent on the additional N supply to the system, which was most prominent under humid growth conditions. There were significant differences between WC and RC in terms of sustaining a relevant percentage in grassland receiving medium to high N-fertilizer applications. The two legumes also differed in terms of N transfer to neighboring plants.
Under drought conditions, legumes produced a higher yield than Lol and occasionally higher but mostly similar yield to grassland consisting of drought-tolerant grasses. Legumes were particularly beneficial under moderate drought conditions when the N-supply to roots was presumably limited. Under severe drought conditions, legume productivity was relatively high and similar to the productivity of drought-tolerant grasses, but the reverse was true in the post-drought period. Legume productivity in the post-drought was considerably lower than drought-tolerant grasses. Therefore, the benefit of legumes during drought periods depends on the intensity of the drought, with legumes having an advantage under moderate and drought-tolerant grasses having an advantage under severe drought. The dry matter productivity of WC and RC under drought was similar, with the exception of the spring drought when RC showed some yield advantages.
It can be concluded that legumes improve the DM yield of mixed grasslands under various drought conditions, but their major benefit is predominately based on their productivity under adequate rainfall. Drought-tolerant grasses are advisable for severe drought conditions. Nevertheless, seeding mixed grassland including legumes was the best option to maintain or increase grassland yield under variable climatic conditions.










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}