
Excretion Dynamics of Arboviruses in Mosquitoes and the Potential Use in Vector Competence Studies and Arbovirus Surveillance

 , , , ,
, , , ,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Cells and Viruses
2.2. Mosquito Infection and Examination of Vector Competence
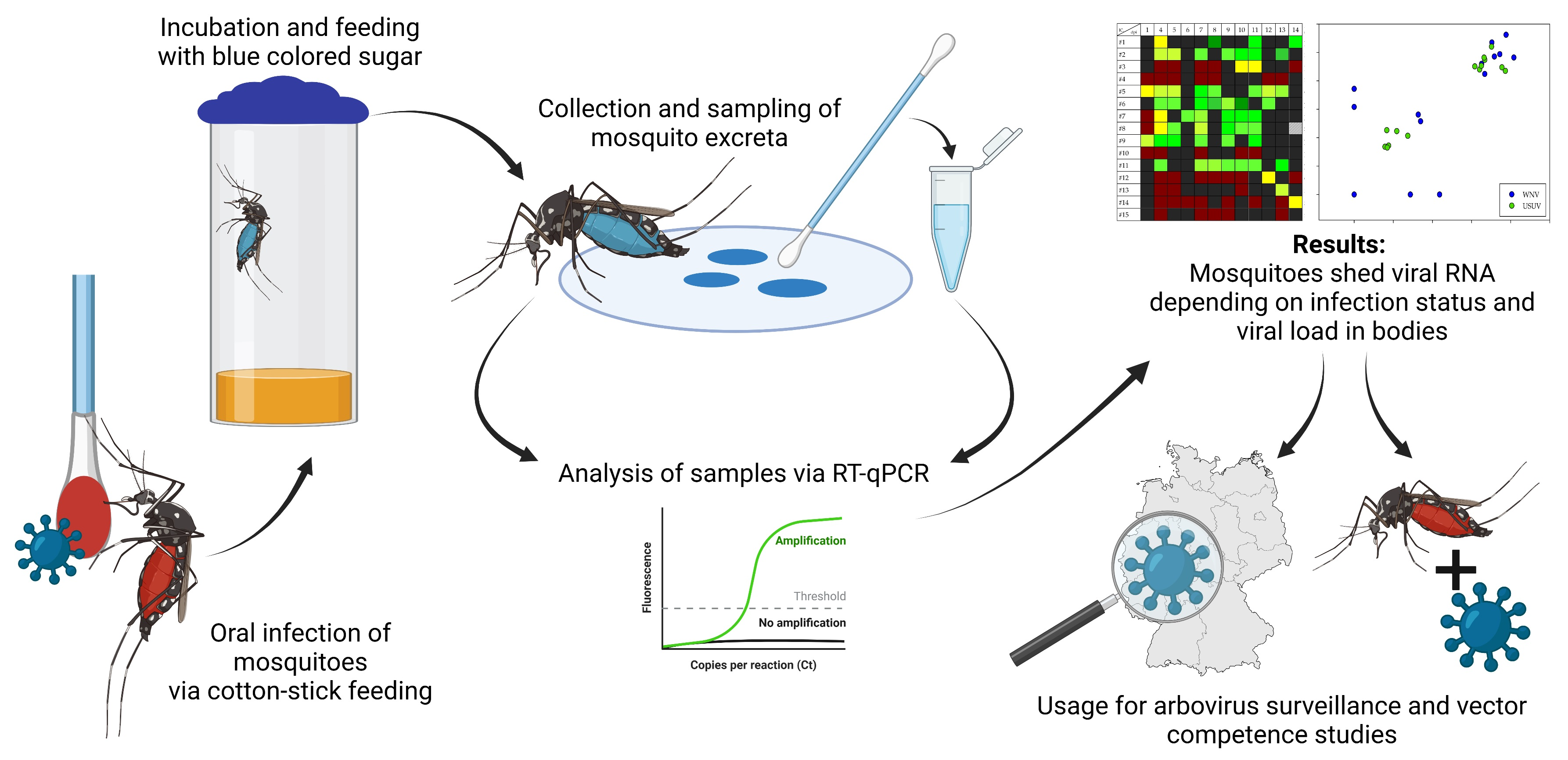
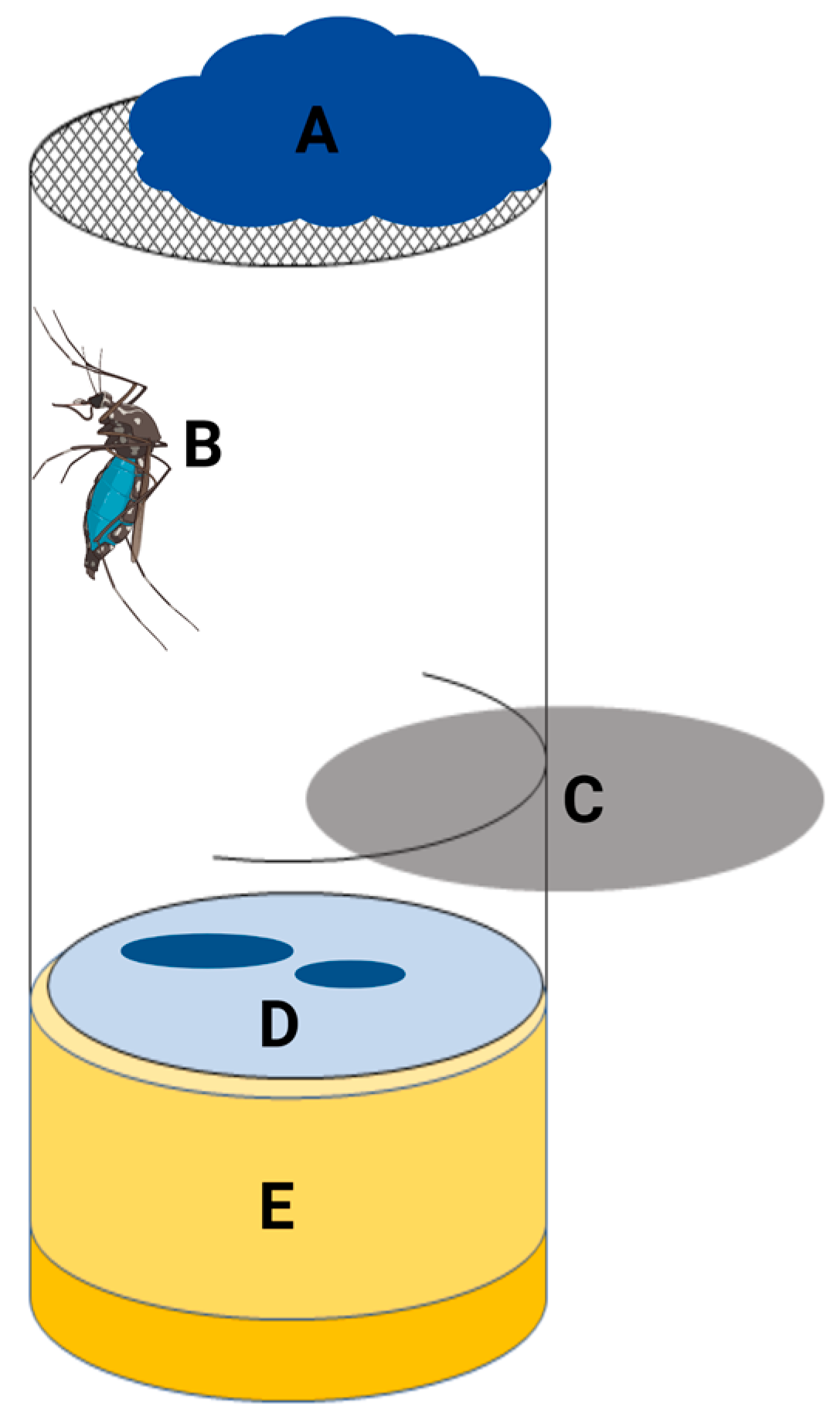
2.3. Collection of Mosquito Excreta
2.4. Nucleic Acid Extractions and Analysis
2.5. Vector Competence Indices and Infection Status
2.6. Statistical Analysis
3. Results
3.1. Vector Competence Trials
3.2. Excretion of Viral RNA via the Infectious Blood Meal
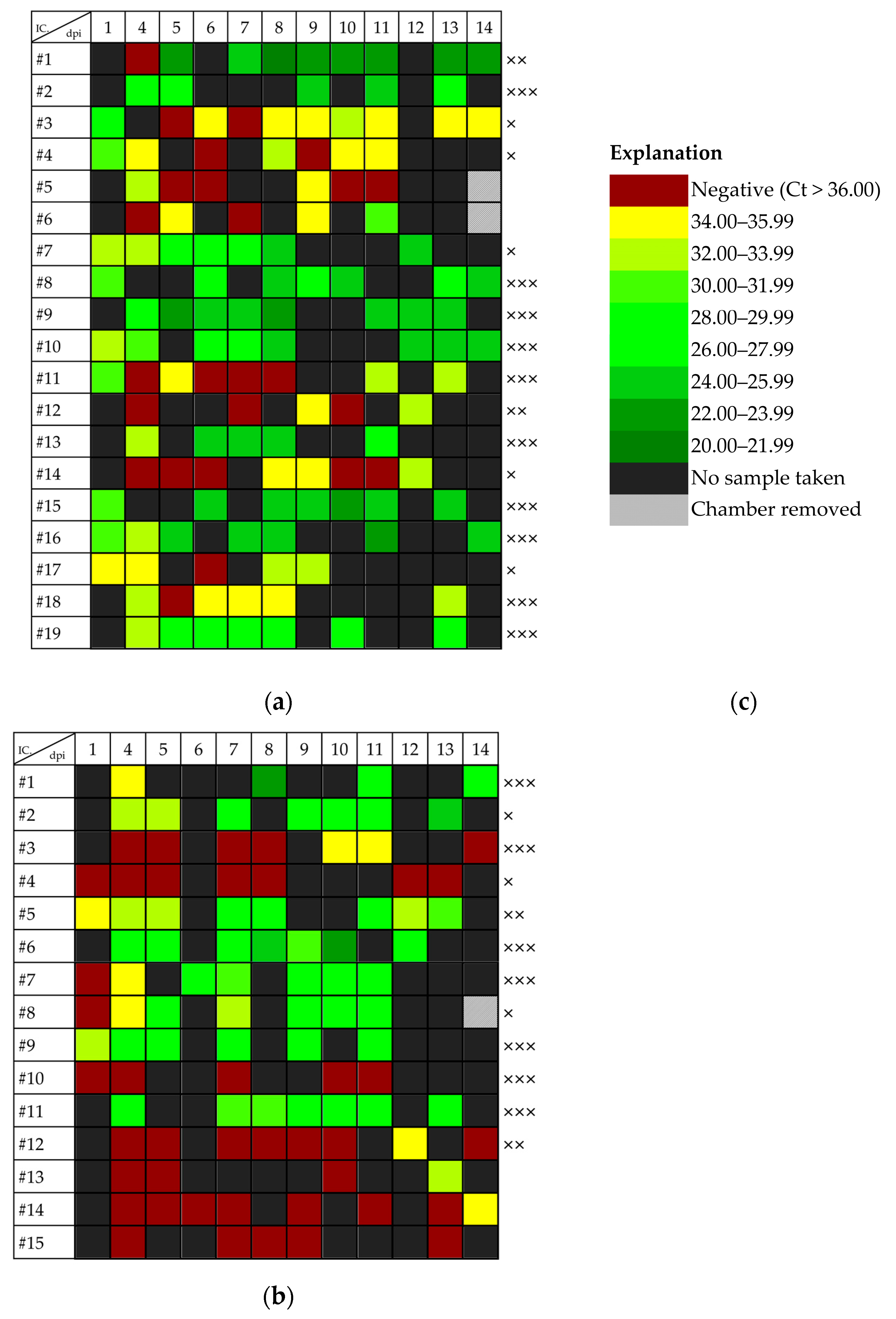
3.3. Dependence of Viral Shedding on Mosquito Infection Status
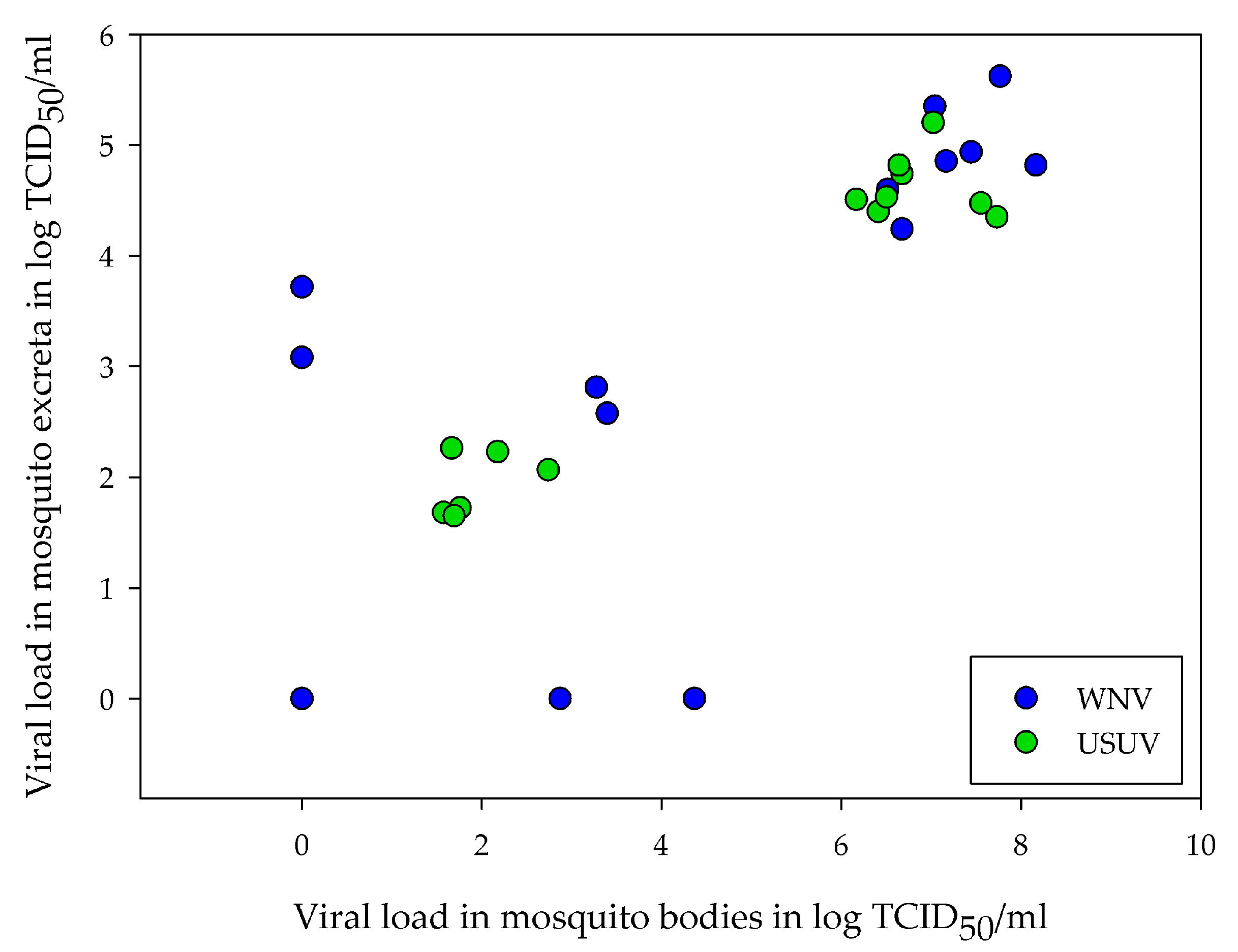
3.4. Correlation between Amount of Viral RNA in Excreta and Viral Load in Mosquitoes
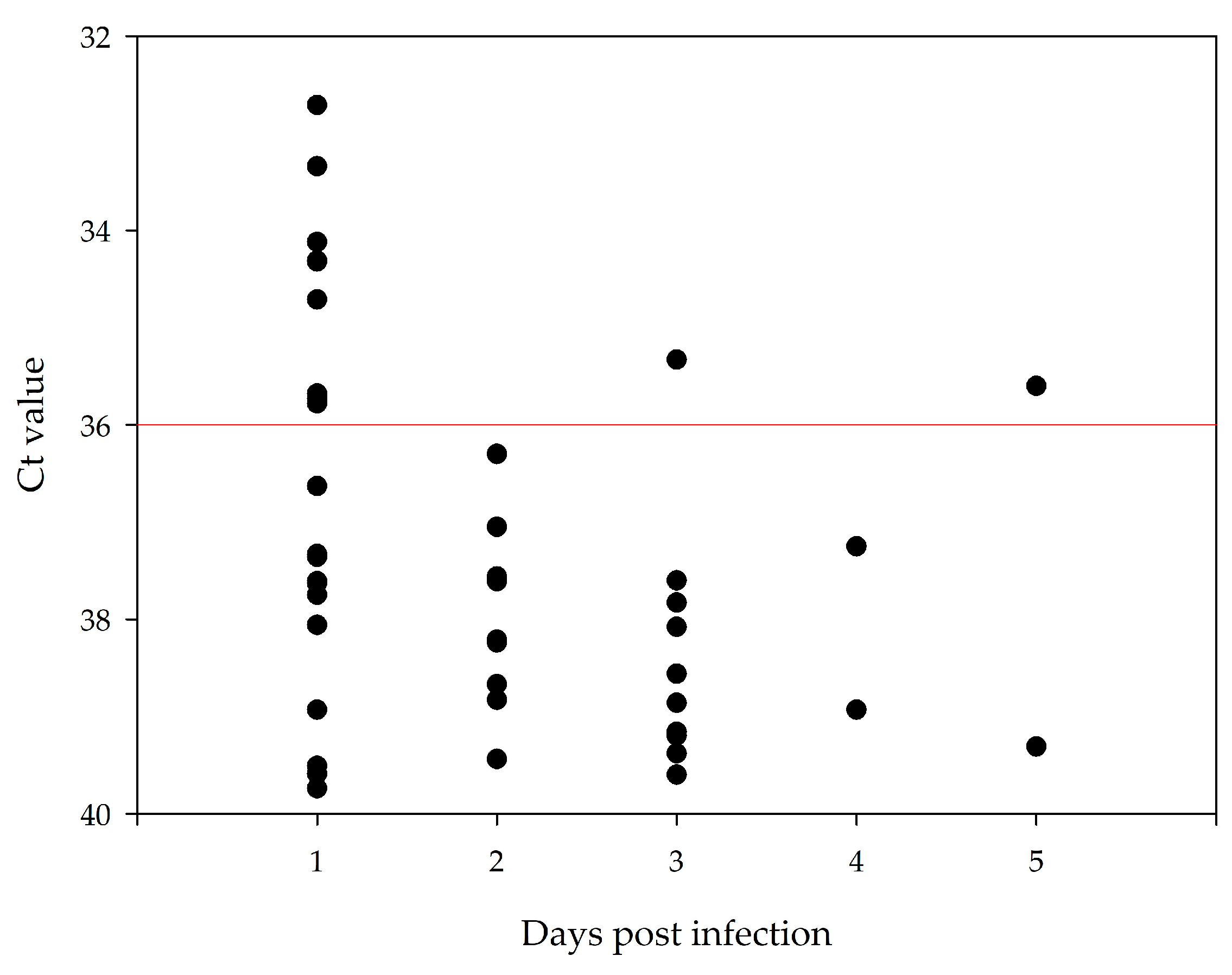
3.5. Duration of Viral Shedding after Infection
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huang, Y.-J.S.; Higgs, S.; Vanlandingham, D.L. Emergence and re-emergence of mosquito-borne arboviruses. Curr. Opin. Virol. 2019, 34, 104–109. [Google Scholar] [CrossRef] [PubMed]
- Gould, E.; Pettersson, J.; Higgs, S.; Charrel, R.; de Lamballerie, X. Emerging arboviruses: Why today? One Health 2017, 4, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Hubálek, Z. Mosquito-borne viruses in Europe. Parasitol. Res. 2008, 103 (Suppl. 1), S29–S43. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, U.; Bergmann, F.; Fischer, D.; Müller, K.; Holicki, C.M.; Sadeghi, B.; Sieg, M.; Keller, M.; Schwehn, R.; Reuschel, M.; et al. Spread of West Nile Virus and Usutu Virus in the German Bird Population, 2019–2020. Microorganisms 2022, 10, 807. [Google Scholar] [CrossRef]
- Bergmann, F.; Trachsel, D.S.; Stoeckle, S.D.; Bernis Sierra, J.; Lübke, S.; Groschup, M.H.; Gehlen, H.; Ziegler, U. Seroepidemiological survey of West Nile virus infections in horses from Berlin/Brandenburg and North Rhine-Westphalia, Germany. Viruses 2022, 14, 243. [Google Scholar] [CrossRef]
- Frank, C.; Schmidt-Chanasit, J.; Ziegler, U.; Lachmann, R.; Preußel, K.; Offergeld, R. West Nile Virus in Germany: An Emerging Infection and Its Relevance for Transfusion Safety. Transfus. Med. Hemotherapy 2022, 49, 192–204. [Google Scholar] [CrossRef]
- Hubálek, Z.; Rudolf, I.; Nowotny, N. Arboviruses pathogenic for domestic and wild animals. Adv. Virus Res. 2014, 89, 201–275. [Google Scholar] [CrossRef]
- Byas, A.D.; Ebel, G.D. Comparative Pathology of West Nile Virus in Humans and Non-Human Animals. Pathogens 2020, 9, 48. [Google Scholar] [CrossRef] [Green Version]
- Domanović, D.; Gossner, C.M.; Lieshout-Krikke, R.; Mayr, W.; Baroti-Toth, K.; Dobrota, A.M.; Escoval, M.A.; Henseler, O.; Jungbauer, C.; Liumbruno, G.; et al. West Nile and Usutu Virus Infections and Challenges to Blood Safety in the European Union. Emerg. Infect. Dis. 2019, 25, 1050–1057. [Google Scholar] [CrossRef] [Green Version]
- Kampen, H.; Werner, D. Die wiederkehrende Notwendigkeit von Stechmücken-Surveillance und -Forschung. Bundesgesundheitsblatt Gesundheitsforschung Gesundheitsschutz 2015, 58, 1101–1109. [Google Scholar] [CrossRef]
- Constant, O.; Gil, P.; Barthelemy, J.; Bolloré, K.; Foulongne, V.; Desmetz, C.; Leblond, A.; Desjardins, I.; Pradier, S.; Joulié, A.; et al. One Health surveillance of West Nile and Usutu viruses: A repeated cross-sectional study exploring seroprevalence and endemicity in Southern France, 2016 to 2020. Euro Surv. 2022, 27, 2200068. [Google Scholar] [CrossRef]
- Vilibic-Cavlek, T.; Savic, V.; Sabadi, D.; Peric, L.; Barbic, L.; Klobucar, A.; Miklausic, B.; Tabain, I.; Santini, M.; Vucelja, M.; et al. Prevalence and molecular epidemiology of West Nile and Usutu virus infections in Croatia in the ‘One health’ context, 2018. Transbound. Emerg. Dis. 2019, 66, 1946–1957. [Google Scholar] [CrossRef]
- Bellini, R.; Calzolari, M.; Mattivi, A.; Tamba, M.; Angelini, P.; Bonilauri, P.; Albieri, A.; Cagarelli, R.; Carrieri, M.; Dottori, M.; et al. The experience of West Nile virus integrated surveillance system in the Emilia-Romagna region: Five years of implementation, Italy, 2009 to 2013. Euro Surv. 2014, 19, 20953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Figuerola, J.; Jiménez-Clavero, M.Á.; Ruíz-López, M.J.; Llorente, F.; Ruiz, S.; Hoefer, A.; Aguilera-Sepúlveda, P.; Jiménez-Peñuela, J.; García-Ruiz, O.; Herrero, L.; et al. A One Health view of the West Nile virus outbreak in Andalusia (Spain) in 2020. Emerg. Microbes Infect. 2022, 11, 2570–2578. [Google Scholar] [CrossRef]
- Jöst, H.; Bialonski, A.; Maus, D.; Sambri, V.; Eiden, M.; Groschup, M.H.; Günther, S.; Becker, N.; Schmidt-Chanasit, J. Isolation of usutu virus in Germany. Am. J. Trop. Med. Hyg. 2011, 85, 551–553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scheuch, D.E.; Schäfer, M.; Eiden, M.; Heym, E.C.; Ziegler, U.; Walther, D.; Schmidt-Chanasit, J.; Keller, M.; Groschup, M.H.; Kampen, H. Detection of Usutu, Sindbis, and Batai viruses in mosquitoes (Diptera: Culicidae) collected in Germany, 2011–2016. Viruses 2018, 10, 389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kampen, H.; Holicki, C.M.; Ziegler, U.; Groschup, M.H.; Tews, B.A.; Werner, D. West Nile virus mosquito vectors (Diptera: Culicidae) in Germany. Viruses 2020, 12, 493. [Google Scholar] [CrossRef]
- Gu, W.; Novak, R.J. Short report: Detection probability of arbovirus infection in mosquito populations. Am. J. Trop. Med. Hyg. 2004, 71, 636–638. [Google Scholar] [CrossRef]
- Girod, R.; Guidez, A.; Carinci, R.; Issaly, J.; Gaborit, P.; Ferrero, E.; Ardillon, V.; Fontaine, A.; Dusfour, I.; Briolant, S. Detection of Chikungunya Virus Circulation Using Sugar-Baited Traps during a Major Outbreak in French Guiana. PLoS Neglected Trop. Dis. 2020, 10, e0004876. [Google Scholar] [CrossRef] [Green Version]
- Flies, E.J.; Toi, C.; Weinstein, P.; Doggett, S.L.; Williams, C.R. Converting Mosquito Surveillance to Arbovirus Surveillance with Honey-Baited Nucleic Acid Preservation Cards. Vector Borne Zoonotic Dis. 2015, 15, 397–403. [Google Scholar] [CrossRef]
- Hall-Mendelin, S.; Ritchie, S.A.; Johansen, C.A.; Zborowski, P.; Cortis, G.; Dandridge, S.; Hall, R.A.; van den Hurk, A.F. Exploiting mosquito sugar feeding to detect mosquito-borne pathogens. Proc. Natl. Acad. Sci. USA 2010, 107, 11255–11259. [Google Scholar] [CrossRef] [PubMed]
- Fynmore, N.; Lühken, R.; Kliemke, K.; Lange, U.; Schmidt-Chanasit, J.; Lurz, P.W.W.; Becker, N. Honey-baited FTA cards in box gravid traps for the assessment of Usutu virus circulation in mosquito populations in Germany. Acta Trop. 2022, 235, 106649. [Google Scholar] [CrossRef] [PubMed]
- Fynmore, N.; Lühken, R.; Maisch, H.; Risch, T.; Merz, S.; Kliemke, K.; Ziegler, U.; Schmidt-Chanasit, J.; Becker, N. Rapid assessment of West Nile virus circulation in a German zoo based on honey-baited FTA cards in combination with box gravid traps. Parasites Vectors 2021, 14, 449. [Google Scholar] [CrossRef] [PubMed]
- Ramírez, A.L.; Hall-Mendelin, S.; Doggett, S.L.; Hewitson, G.R.; McMahon, J.L.; Ritchie, S.A.; van den Hurk, A.F. Mosquito excreta: A sample type with many potential applications for the investigation of Ross River virus and West Nile virus ecology. PLoS Neglected Trop. Dis. 2018, 12, e0006771. [Google Scholar] [CrossRef]
- Fontaine, A.; Jiolle, D.; Moltini-Conclois, I.; Lequime, S.; Lambrechts, L. Excretion of dengue virus RNA by Aedes aegypti allows non-destructive monitoring of viral dissemination in individual mosquitoes. Sci. Rep. 2016, 6, 24885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- L’Ambert, G.; Gendrot, M.; Briolant, S.; Nguyen, A.; Pages, S.; Bosio, L.; Palomo, V.; Gomez, N.; Benoit, N.; Savini, H.; et al. Analysis of trapped mosquito excreta as a noninvasive method to reveal biodiversity and arbovirus circulation. Mol. Ecol. Resour. 2022, 23, 410–423. [Google Scholar] [CrossRef] [PubMed]
- Meyer, D.B.; Ramirez, A.L.; van den Hurk, A.F.; Kurucz, N.; Ritchie, S.A. Development and Field Evaluation of a System to Collect Mosquito Excreta for the Detection of Arboviruses. J. Med. Entomol. 2019, 56, 1116–1121. [Google Scholar] [CrossRef]
- Ramírez, A.L.; Colmant, A.M.G.; Warrilow, D.; Huang, B.; Pyke, A.T.; McMahon, J.L.; Meyer, D.B.; Graham, R.M.A.; Jennison, A.V.; Ritchie, S.A.; et al. Metagenomic Analysis of the Virome of Mosquito Excreta. mSphere 2020, 5, e00587-20. [Google Scholar] [CrossRef]
- Minetti, C.; Pilotte, N.; Zulch, M.; Canelas, T.; Tettevi, E.J.; Veriegh, F.B.D.; Osei-Atweneboana, M.Y.; Williams, S.A.; Reimer, L.J. Field evaluation of DNA detection of human filarial and malaria parasites using mosquito excreta/feces. PLoS Neglected Trop. Dis. 2020, 14, e0008175. [Google Scholar] [CrossRef] [Green Version]
- Ramírez, A.L.; van den Hurk, A.F.; Mackay, I.M.; Yang, A.S.P.; Hewitson, G.R.; McMahon, J.L.; Boddey, J.A.; Ritchie, S.A.; Erickson, S.M. Malaria surveillance from both ends: Concurrent detection of Plasmodium falciparum in saliva and excreta harvested from Anopheles mosquitoes. Parasit Vectors 2019, 12, 355. [Google Scholar] [CrossRef] [Green Version]
- Ramírez, A.L.; van den Hurk, A.F.; Meyer, D.B.; Ritchie, S.A. Searching for the proverbial needle in a haystack: Advances in mosquito-borne arbovirus surveillance. Parasit Vectors 2018, 11, 320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beermann, S.; Dobler, G.; Faber, M.; Frank, C.; Habedank, B.; Hagedorn, P.; Kampen, H.; Kuhn, C.; Nygren, T.; Schmidt-Chanasit, J.; et al. Auswirkungen von Klimaveränderungen auf Vektor- und Nagetier-assoziierte Infektionskrankheiten. J. Health Monit. 2023, 8, 7–35. [Google Scholar] [CrossRef]
- Körsten, C.; AL-Hosary, A.A.; Holicki, C.M.; Schäfer, M.; Tews, B.A.; Vasić, A.; Ziegler, U.; Groschup, M.H.; Silaghi, C. Simultaneous Coinfections with West Nile Virus and Usutu Virus in Culex pipiens and Aedes vexans Mosquitoes. Transbound. Emerg. Dis. 2023, 2023, 6305484. [Google Scholar] [CrossRef]
- Heitmann, A.; Jansen, S.; Lühken, R.; Leggewie, M.; Schmidt-Chanasit, J.; Tannich, E. Forced salivation as a method to analyze vector competence of mosquitoes. J. Vis. Exp. 2018, 138, e57980. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, B.; Depner, K.; Schirrmeier, H.; Beer, M. A universal heterologous internal control system for duplex real-time RT-PCR assays used in a detection system for pestiviruses. J. Virol. Methods 2006, 136, 200–209. [Google Scholar] [CrossRef] [PubMed]
- Eiden, M.; Vina-Rodriguez, A.; Hoffmann, B.; Ziegler, U.; Groschup, M.H. Two new real-time quantitative reverse transcription polymerase chain reaction assays with unique target sites for the specific and sensitive detection of lineages 1 and 2 West Nile virus strains. J. Vet. 2010, 22, 748–753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwaiger, M.; Cassinotti, P. Development of a quantitative real-time RT-PCR assay with internal control for the laboratory detection of tick borne encephalitis virus (TBEV) RNA. J. Clin. Virol. 2003, 27, 136–145. [Google Scholar] [CrossRef] [PubMed]
- Körsten, C.; AL-Hosary, A.A.; Schäfer, M.; Tews, B.A.; Werner, D.; Kampen, H.; Vasic, A.; Silaghi, C. Vector Competence of German Aedes punctor (Kirby, 1837) for West Nile Virus Lineages 1 and 2. Viruses 2022, 14, 2787. [Google Scholar] [CrossRef]
- Ramírez, A.L.; Hall-Mendelin, S.; Hewitson, G.R.; McMahon, J.L.; Staunton, K.M.; Ritchie, S.A.; van den Hurk, A.F. Stability of West Nile Virus (Flaviviridae: Flavivirus) RNA in Mosquito Excreta. J. Med. Entomol. 2019, 56, 1135–1138. [Google Scholar] [CrossRef]
- Gooding, R.H. Digestive processes of haematophagous insects. I. A literature review. Quaest. Entomol. 1972, 8, 5–60. [Google Scholar]
- Vogels, C.B.; Göertz, G.P.; Pijlman, G.P.; Koenraadt, C.J. Vector competence of European mosquitoes for West Nile virus. Emerg. Microbes Infect. 2017, 6, e96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komar, N.; Langevin, S.; Hinten, S.; Nemeth, N.; Edwards, E.; Hettler, D.; Davis, B.; Bowen, R.; Bunning, M. Experimental infection of North American birds with the New York 1999 strain of West Nile virus. Emerg. Infect. Dis. 2003, 9, 311–322. [Google Scholar] [CrossRef] [PubMed]
- Franz, A.W.E.; Kantor, A.M.; Passarelli, A.L.; Clem, R.J. Tissue Barriers to Arbovirus Infection in Mosquitoes. Viruses 2015, 7, 3741–3767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boelke, M.; Bestehorn, M.; Marchwald, B.; Kubinski, M.; Liebig, K.; Glanz, J.; Schulz, C.; Dobler, G.; Monazahian, M.; Becker, S.C. First Isolation and Phylogenetic Analyses of Tick-Borne Encephalitis Virus in Lower Saxony, Germany. Viruses 2019, 11, 462. [Google Scholar] [CrossRef] [Green Version]
- Topp, A.-K.; Springer, A.; Dobler, G.; Bestehorn-Willmann, M.; Monazahian, M.; Strube, C. New and Confirmed Foci of Tick-Borne Encephalitis Virus (TBEV) in Northern Germany Determined by TBEV Detection in Ticks. Pathogens 2022, 11, 126. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Garver, L.S.; Bingham, K.M.; Hang, J.; Jochim, R.C.; Davidson, S.A.; Richardson, J.H.; Jarman, R.G. Feasibility of Using the Mosquito Blood Meal for Rapid and Efficient Human and Animal Virus Surveillance and Discovery. Am. J. Trop. Med. Hyg. 2015, 93, 1377–1382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez, M.C.; Cime-Castillo, J.; Argotte-Ramos, R.; Vargas, V.; Correa-Morales, F.; Sánchez-Tejeda, G.; Lanz-Mendoza, H. Detection of NS1 protein from dengue virus in excreta and homogenates of wild-caught Aedes aegypti mosquitoes using monoclonal antibodies. Pathog. Dis. 2022, 80, ftac002. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus Strain | GenBank Accession No. | Titer 1 |
|---|---|---|
| WNV lineage 1 (Italy, 2008) | JF719066 | 5.62 × 107 |
| WNV lineage 2 (Germany, 2018) | MH924836 | 1.33 × 109 |
| USUV lineage Europe 3 (Germany, 2011) | HE599647 | 5.01 × 107 |
| USUV lineage Europe 3 (Germany, 2016) | KY084498 | 5.62 × 107 |
| TBEV Western Neudoerfl | U27495 | 1.00 × 108 |
| Experiment No. | Virus | Mosquito Species | Incubation Period (Days) | Number of Mosquitoes per Incubation Chamber | Material Used for Excreta Collection |
|---|---|---|---|---|---|
| #1 | WNV lineage 1 (Italy, 2008) | AeV | 14 | 1 or 5 | FTA cards; Parafilm and 1 × PBS |
| #2 | WNV lineage 1 (Italy, 2008) | CxM | 20 | 5 | FTA cards; Parafilm and 1 × PBS |
| #3 | WNV lineage 2 (Germany, 2018) | AeV | 14 | 1 | Parafilm and AVL buffer |
| #4 | TBEV Western Neudoerfl | CxM | 14 | 1 | Parafilm and AVL buffer |
| #5 | USUV lineage Europe 3 (Germany, 2016) | CxM | 14 | 1 | Parafilm and AVL buffer; only if excreta were visible |
| #6 | USUV lineage Europe 3 (Germany, 2011) | CxM | 14 | 1 | Parafilm and AVL buffer; only if excreta were visible |
| #7 | WNV lineage 2 (Germany, 2018) | CxM | 14 | 1 | Parafilm and AVL buffer; only if excreta were visible |
| Experiment No. | Virus | Mosquito Species | Blood Meal Titer 1 | Mosquitoes Examined 2 | IR % (n/n) | DR % (n/n) | TR % (n/n) |
|---|---|---|---|---|---|---|---|
| #1 | WNV lineage 1 (Italy, 2008) | AeV | 6.125 | 10 | 0 (0/10) | N/A | N/A |
| #2 | WNV lineage 1 (Italy, 2008) | CxM | 5.500 | 2 | 0 (0/2) | N/A | N/A |
| #3 | WNV lineage 2 (Germany, 2018) | AeV | 7.438 | 8 | 0 (0/8) | N/A | N/A |
| #4 | TBEV Western Neudoerfl | CxM | 7.063 | 24 | 0 (0/24) | N/A | N/A |
| #5 | USUV lineage Europe 3 (Germany, 2016) | CxM | 6.063 | 18 | 5.56 (1/18) | 0 (0/1) | N/A |
| #6 | USUV lineage Europe 3 (Germany, 2011) | CxM | 7.125 | 16 | 100.00 (16/16) | 75.00 (12/16) | 83.33 (10/12) |
| #7 | WNV lineage 2 (Germany, 2018) | CxM | 6.844 | 14 | 78.54 (11/14) | 81.82 (9/11) | 77.78 (7/9) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Körsten, C.; Vasić, A.; AL-Hosary, A.A.; Tews, B.A.; Răileanu, C.; Silaghi, C.; Schäfer, M. Excretion Dynamics of Arboviruses in Mosquitoes and the Potential Use in Vector Competence Studies and Arbovirus Surveillance. Trop. Med. Infect. Dis. 2023, 8, 410. https://doi.org/10.3390/tropicalmed8080410
Körsten C, Vasić A, AL-Hosary AA, Tews BA, Răileanu C, Silaghi C, Schäfer M. Excretion Dynamics of Arboviruses in Mosquitoes and the Potential Use in Vector Competence Studies and Arbovirus Surveillance. Tropical Medicine and Infectious Disease. 2023; 8(8):410. https://doi.org/10.3390/tropicalmed8080410
Chicago/Turabian StyleKörsten, Christin, Ana Vasić, Amira A. AL-Hosary, Birke A. Tews, Cristian Răileanu, Cornelia Silaghi, and Mandy Schäfer. 2023. "Excretion Dynamics of Arboviruses in Mosquitoes and the Potential Use in Vector Competence Studies and Arbovirus Surveillance" Tropical Medicine and Infectious Disease 8, no. 8: 410. https://doi.org/10.3390/tropicalmed8080410