
A Novel Designed Sandwich ELISA for the Detection of Echinococcus granulosus Antigen in Camels for Diagnosis of Cystic Echinococcosis
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Approval
2.2. Study Area
2.3. Animals
2.4. Samples
2.4.1. Blood
2.4.2. Hydatid Cysts
2.5. Assessment of Fertility and Viability of the Hydatid Cysts
2.6. Separation of Germinal Layers
2.7. DNA Extraction
2.8. Polymerase Chain Reaction (PCR) and Electrophoresis
2.9. Sequencing and Phylogenetic Analysis
2.10. Preparation of Hydatid Cyst Germinal Layer Antigen (GlAg)
2.11. Rabbit IgG Polyclonal Antibodies (Anti-Echinococcus GlAb)
2.12. Purification of Rabbit IgG Polyclonal Antibodies Using Protein A Affinity Chromatography
2.13. Assessment of Rabbit IgG Polyclonal Antibodies Reactivity Against GlAg Using Indirect ELISA
2.14. Conjugation of Rabbit IgG Polyclonal Antibodies with HRP Enzyme
2.15. Characterization of Rabbit IgG Polyclonal Antibodies and Crude GlAg
2.16. Sandwich ELISA
2.17. Statistical Analysis
3. Results
3.1. Microscopical Examination and PM Diagnosis
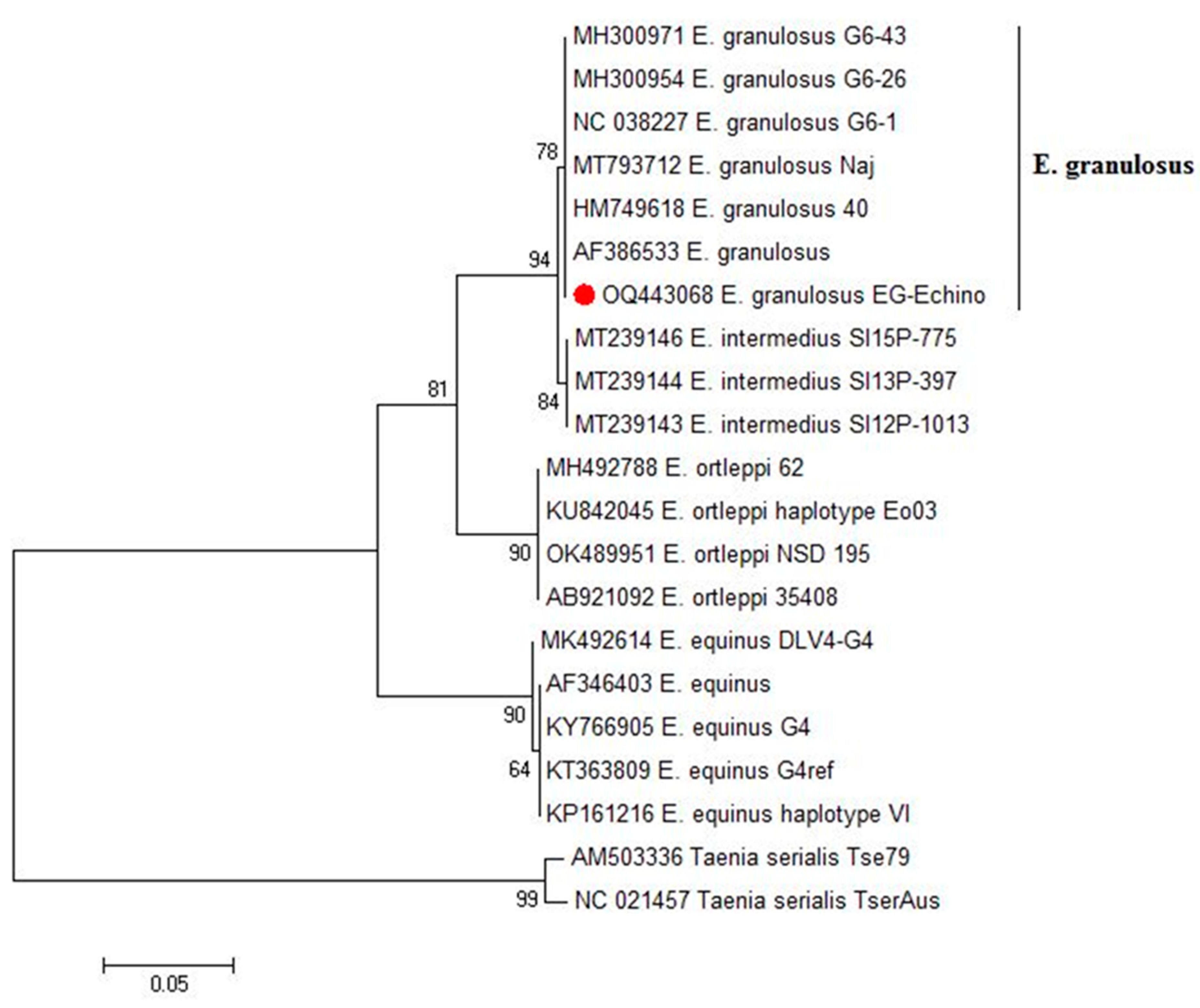
3.2. Standard PCR and Phylogenetic Analysis
3.3. Electrophoretic Profile of GlAg and Rabbit IgG Polyclonal Antibodies
3.4. Diagnostic Potency of Rabbit IgG Polyclonal Antibodies against GlAg by Indirect ELISA
3.5. Immunoreactivity and Cross-Reactivity of Rabbit IgG Polyclonal Antibodies for Detection of the Circulating E. granulosus Antigens in Camels’ Sera by Sandwich ELISA
3.6. Detection of Circulating E. granulosus Antigen in Camel Sera for Diagnosis of CE by Sandwich ELISA
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- AlSalman, A.; Mathewson, A.; Martin, I.W.; Mahatanan, R.; Talbot, E.A. Cystic Echinococcosis in Northern New Hampshire, USA. Emerg. Infect. Dis. 2023, 29, 1057–1058. [Google Scholar] [CrossRef] [PubMed]
- Budke, C.M.; Casulli, A.; Kern, P.; Vuitton, D.A. Cystic and alveolar echinococcosis: Successes and continuing challenges. PLoS Negl. Trop. Dis. 2017, 11, e0005477. [Google Scholar] [CrossRef] [Green Version]
- Tamarozzi, F.; Legnardi, M.; Fittipaldo, A.; Drigo, M.; Cassini, R. Epidemiological distribution of Echinococcus granulosus s.l. infection in human and domestic animal hosts in European Mediterranean and Balkan countries: A systematic review. PLoS Negl. Trop. Dis. 2020, 14, e0008519. [Google Scholar] [CrossRef] [PubMed]
- Deplazes, P.; Rinaldi, L.; Alvarez Rojas, C.A.; Torgerson, P.R.; Harandi, M.F.; Romig, T.; Antolova, D.; Schurer, J.M.; Lahmar, S.; Cringoli, G.; et al. Global Distribution of Alveolar and Cystic Echinococcosis. Adv. Parasitol. 2017, 95, 315–493. [Google Scholar] [PubMed] [Green Version]
- Hua, R.Q.; Du, X.D.; He, X.; Gu, X.B.; Xie, Y.; He, R.; Xu, J.; Peng, X.R.; Yang, G.Y. Genetic diversity of Echinococcus granulosus sensu lato in China: Epidemiological studies and systematic review. Transbound. Emerg. Dis. 2022, 69, e1382. [Google Scholar] [CrossRef] [PubMed]
- Sen, P.; Demirdal, T.; Nemli, S.A. Evaluation of clinical, diagnostic and treatment aspects in hydatid disease: Analysis of an 8-year experience. Afr. Health Sci. 2019, 19, 2431–2438. [Google Scholar] [CrossRef]
- Nigo, K.L.S.; John, B.T.; Lobojo, D.L.; Lita, E.P.; Osman, A.Y.; Shuaib, Y.A. Prevalence and financial losses of cystic echinococcosis in slaughtered goats at Gumbo Slab in Juba County, South Sudan. Parasitologia 2022, 2, 54–62. [Google Scholar] [CrossRef]
- Craig, P.; Mastin, A.; Kesteren, F.; Boufana, B.B. Echinococcus granulosus: Epidemiology and state-of-the-art of diagnostics in animals. Vet. Parasitol. 2015, 213, 132–148. [Google Scholar] [CrossRef] [Green Version]
- WHO. World Health Organization. Echinococcosis, Geneva, Weekly Epidemiological Record, 2019, 94, 574–579. Available online: https://extranet.who.int/iris/restricted/bitstream/handle/10665/330003/WER9448-eng-fre.pdf?ua=1 (accessed on 16 February 2023).
- Latif, A.A.; Tanveer, A.; Maqbool, A.; Siddiqi, N.; Kyaw-Tanner, M.; Traub, R.J. Morphological and molecular characterisation of Echinococcus granulosus in livestock and humans in Punjab, Pakistan. Vet. Parasitol. 2010, 170, 44–49. [Google Scholar] [CrossRef]
- Ahmed, M.E.; Eltom, K.H.; Musa, N.O.; Ali, I.A.; Elamin, F.M.; Grobusch, M.P.; Aradaib, I.E. First report on circulation of Echinococcus ortleppi in the one humped camel (Camelus dromedaries), Sudan. BMC Vet. Res. 2013, 9, 127. [Google Scholar] [CrossRef] [Green Version]
- Abbas, I.E.A.; Al-Kappany, Y.M.; Al-Araby, M.A. Prevalence and molecular characterization of hydatid cyst isolates from cattle in Egypt. Asian J. Anim. Vet. Adv. 2016, 11, 794–804. [Google Scholar] [CrossRef] [Green Version]
- Lahmar, S.; Trifi, M.; Ben Naceur, S.; Bouchhima, T.; Lahouar, N.; Lamouchi, I.; Maâmouri, N.; Selmi, R.; Dhibi, M.; Torgerson, P.R. Cystic echinococcosis in slaughtered domestic ruminants from Tunisia. J. Helminthol. 2013, 87, 318–325. [Google Scholar] [CrossRef] [Green Version]
- Kassem, H.H.; Abdel-Kader, A.K.; Nass, S.A. Prevalence of hydatid cysts in slaughtered animals in Sirte, Libya. J. Egypt Soc. Parasitol. 2013, 43, 33–40. [Google Scholar] [PubMed]
- Omer, R.A.; Dinkel, A.; Romig, T.; Mackenstedt, U.; Elnahas, A.A.; Aradaib, I.E.; Ahmed, M.E.; Elmalik, K.H.; Adam, A. A molecular survey of cystic echinococcosis in Sudan. Vet. Parasitol. 2010, 169, 340–346. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, K.; Thomas, R.; Peter, K.; Omer, R.A. A molecular survey on cystic echinococcosis in Sinnar area, Blue Nile state (Sudan). Chin. Med. J. 2011, 124, 2829–2833. [Google Scholar] [PubMed]
- Ibrahim, M.M. Study of cystic echinococcosis in slaughtered animals in Al Baha region, Saudi Arabia: Interaction between some biotic and abiotic factors. Acta Trop. 2010, 113, 26–33. [Google Scholar] [CrossRef]
- Omondi, H.A.; Gitau, G.; Gathura, P.; Mulinge, E.; Zeyhle, E.; Kimeli, P.; Bett, B. Prevalence and genotyping of Echinococcus granulosus sensu lato from livestock in north-eastern Kenya. J. Helminthol. 2020, 94, e205. [Google Scholar] [CrossRef]
- Abdel Aaty, H.; Abdel-Hameed, D.; Alam-Eldin, Y.; El-Shennawy, S.; Aminou, H.; Makled, S.S.; Darweesh, S.K. Molecular genotyping of Echinococcus granulosus in animal and human isolates from Egypt. Acta Trop. 2012, 121, 125–128. [Google Scholar] [CrossRef]
- Omar, M.; Sultan, K.; Haridy, M.; Omran, A. Prevalence of cystic echinococcosis in slaughtered ruminants in different abattoirs, Upper Egypt. Am. J. Anim. Vet. Sci. 2013, 8, 117–121. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, A.B.; Ras, R.; Mahmoud, A.F.; El-Ghazaly, E.; Widmer, G.; Dahshan, H.; Elsohaby, I. Prevalence and bacterial isolation from hydatid cysts in dromedary camels (Camelus dromedarius) slaughtered at Sharkia abattoirs, Egypt. J. Parasit. Dis. 2021, 45, 236–243. [Google Scholar] [CrossRef]
- El-Dakhly, K.M.; Arafa, W.M.; El-Nahass, E.N.; Shokier, K.A.M.; Noaman, A.F. The current prevalence and diversity of cystic echinococcosis in slaughtered animals in Egypt. J. Parasit. Dis. 2019, 43, 711–717. [Google Scholar] [CrossRef]
- Khalifa, N.; Khater, H.F.; Fahmy, A.H.; Radwan, E.I.M.; Afify, S.A. Genotyping and phylogenetic analysis of cystic Echinococcosis isolated from camels and humans in Egypt. Am. J. Epidemiol. Infect. Dis. 2014, 2, 74–82. [Google Scholar] [CrossRef]
- Elshahawy, I.S.; El-Seify, M.A.; Ahamed, Z.K.; Fawaz, M.M. Occurrence and Phylogenetic Description of Cystic Echinococcosis Isolate from Egyptian Camel (Camelus Dromedarius). Helminthologia 2022, 59, 253–264. [Google Scholar] [CrossRef]
- Shoulah, S.A.; Gaballa, M.M.S.; Marawan, M.A.; Saqr, S.A.; Abdelhady, A.; Alzahrani, H.A.; Wakid, M.H.; Al-Jabr, O.A.; Selim, A. Pathological findings and oxidative stress status associated with hydatidosis in dromedary camels. Vet. Sci. 2023, 10, 74. [Google Scholar] [CrossRef] [PubMed]
- Craig, P.S.; Hegglin, D.; Lightowlers, M.W.; Torgerson, P.R.; Wang, Q. Chapter Two—Echinococcosis: Control and Prevention. Adv. Parasitol. 2017, 96, 55–158. [Google Scholar] [PubMed]
- Toaleb, N.I.; Derbala, A.A.; Abdel-Rahman, E.H. Comparative diagnostic evaluation of crude and isolated fractions of Echinococcus granulosus in dogs. Glob. Vet. 2011, 7, 587–592. [Google Scholar]
- Toaleb, N.I.; Helmy, M.S.; El Shanawany, E.E.; Abdel-Rahman, E.H. A simple and efficient purification method of native immunoreactive antigen for diagnosis of camel hydatidosis. Vet. World 2020, 13, 141–146. [Google Scholar] [CrossRef] [Green Version]
- Hassanain, M.A.; Toaleb, N.I.; Shaapan, R.M.; Hassanain, N.A.; Maher, A.; Yousif, A.B. Immunological detection of human and camel cystic echinococcosis using different antigens of hydatid cyst fluid, protoscoleces, and germinal layers. Vet. World 2021, 14, 270–275. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Wen, H.; Li, J.; Lin, R.; McManus, D.P. Immunology and immunodiagnosis of cystic echinococcosis: An update. Clin. Dev. Immunol. 2012, 2012, 101895. [Google Scholar] [CrossRef]
- Keong, B.; Wilkie, B.; Sutherland, T.; Fox, A. Hepatic cystic echinococcosis in Australia: An update on diagnosis and management. ANZ J. Surg. 2018, 88, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Manzano-Román, R.; Sánchez-Ovejero, C.; Hernández-González, A.; Casulli, A.; Siles-Lucas, M. Serological diagnosis and follow-up of human cystic echinococcosis: A new hope for the future? Biomed Res. Int. 2015, 2015, 428205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mamuti, W.; Sako, Y.; Nakao, M.; Xiao, N.; Nakaya, K.; Ishikawa, Y.; Yamasaki, H.; Lightowlers, M.W.; Ito, A. Recent advances in characterization of Echinococcus antigen B. Parasitol. Int. 2006, 55, S57–S62. [Google Scholar] [CrossRef] [PubMed]
- Rahimi, H.; Sadjjadi, S.; Sarkari, B. Performance of antigen B isolated from different hosts and cyst locations in diagnosis of cystic echinococcosis. Iran. J. Parasitol. 2011, 6, 12–19. [Google Scholar]
- Kandil, O.M.; El-Hakim, A.E.; Gad, A.A.M.; Abu El-Ezz, N.M.; Mahmoud, M.S.; Hendawy, S.H.; Salama, D.B. Camel hydatidosis diagnostic kit: Optimization of turnip and horseradish peroxidase conjugates using glutaraldehyde method. J. Parasit. Dis. 2020, 44, 230–238. [Google Scholar] [CrossRef]
- Devi, C.S.; Parija, S.C. A new serum hydatid antigen detection test for diagnosis of cystic echinococcosis. Am. J. Trop. Med. Hyg. 2003, 69, 525–528. [Google Scholar] [CrossRef] [Green Version]
- Sadjjadi, S.M.; Sedaghat, F.; Hosseini, S.V.; Sarkari, B. Serum antigen and antibody detection in echinococcosis: Application in serodiagnosis of human hydatidosis. Korean J. Parasitol. 2009, 47, 153–157. [Google Scholar] [CrossRef] [Green Version]
- Chaya, D.; Parija, S.C. Evaluation of a newly designed sandwich enzyme linked immunosorbent assay for the detection of hydatid antigen in serum, urine and cyst fluid for diagnosis of cystic echinococcosis. Trop. Parasitol. 2013, 3, 125–131. [Google Scholar]
- El Deeb, S.; Aly, I.; Mahna, N.; Faried, A.; Zalat, R.; Younis, M. Purification and characterization of Echinococcus granulosus cathepsin-B protein and evaluation of its role as a diagnostic marker. Glob. Vet. 2017, 18, 137–145. [Google Scholar]
- Shariff, G.M.; Parija, S.C. Co-agglutination (Co-A) test for circulating antigen in hydatid disease. J. Med. Microbiol. 1993, 38, 231–234. [Google Scholar] [CrossRef] [PubMed]
- El-Sayed, A.M.; El-Diasty, M.M.; Elbeskawy, M.A.; Zakaria, M.; Younis, E.E. Prevalence of camel brucellosis at Al-Shalateen area. Mansoura Vet. Med. J. 2017, 18, 33–44. [Google Scholar] [CrossRef]
- Dyab, A.K.; Mohamed, G.M.; Abdella, O.H. Seroprevalence of Hydatidosis in Camels of Assuit Province, Egypt. Madridge J. Vaccines 2017, 1, 5–8. [Google Scholar] [CrossRef] [Green Version]
- Smyth, J.D. Studies on tapeworm physiology. XI. In vitro cultivation of Echinococcus granulosus from protoscolex to the strobilar stage. Parasitol. 1967, 57, 111–133. [Google Scholar] [CrossRef] [Green Version]
- Daryani, A.; Alaei, R.; Arab, R.; Sharif, M.; Dehghan, M.H.; Ziaei, H. The prevalence, intensity and viability of hydatid cysts in slaughtered animals in the Ardabil province of Northwest Iran. J. Helminthol. 2007, 81, 13–17. [Google Scholar] [CrossRef]
- Aboelhadid, S.M.; El-Dakhly, K.M.; Yanai, T.; Fukushi, H.; Hassanin, K.M. Molecular characterization of Echinococcus granulosus in Egyptian donkeys. Vet. Parasitol. 2013, 193, 292–296. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the folin phenol reagent. Biol. Chem. J. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Guobadia, E.E.; Fagbemi, B.O. The isolation of Fasciola gigantica-specific antigens and their use in the serodiagnosis of fascioliasis in sheep by the detection of circulating antigens. Vet. Parasitol. 1997, 68, 269–282. [Google Scholar] [CrossRef] [PubMed]
- Engvall, E.; Perlmann, P. Enzyme-linked immunosorbent assay (ELISA). Quantitative assay of immunoglobulin G. Immunochemistry 1971, 8, 871–874. [Google Scholar] [CrossRef]
- Abd El Hafez, S.M.; Anwar, A.M.; Ibrahim, A.M.; Mahmoud, M.B.; Hassan, H.M. Preparation of fluoresce isothiocyanate conjugated IgG (FITC) anti-camel and anti-buffalo. Nat. Sci. J. 2010, 8, 342–347. [Google Scholar]
- Avrameas, S. Coupling of enzymes to proteins with glutaraldehyde. Use of the conjugates for the detection of antigens and antibodies. Immunochem. J. 1969, 5, 43–52. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Anvarov, K.; Khajibaev, A.; Hong, S.; Hong, S. Serodiagnosis of echinococcosis by ELISA using cystic fluid from Uzbekistan sheep. Korean J. Parasitol. 2013, 51, 313–317. [Google Scholar] [CrossRef] [PubMed]
- Parikh, R.; Mathai, A.; Parikh, S.; Sekhar, G.C.; Thomas, R. Understanding and using sensitivity, specificity and predictive values. Ind. J. Ophthal. 2008, 56, 45–50. [Google Scholar] [CrossRef]
- Kern, P.; da Silva, A.M.; Akhan, O.; Müllhaupt, B.; Vizcaychipi, K.A.; Budke, C.; Vuitton, D.A. The echinococcosis: Diagnosis, clinical management and burden of disease. Adv. Parasitol. 2017, 96, 259–369. [Google Scholar]
- Abo-Aziza, F.A.M.; Oda, S.S.; Aboelsoued, D.; Farag, T.K.; Almuzaini, A.M. Variabilities of hydatidosis in domestic animals slaughtered at Cairo and Giza abattoirs, Egypt. Vet. World 2019, 12, 998–1007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torgerson, P.R. The economic effects of echinococcosis. Acta Trop. 2003, 85, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Beigh, A.B.; Darzi, M.M.; Bashir, S.; Kashani, B.; Shah, A.; Shah, S.A. Gross and histopathological alterations associated with cystic echinococcosis in small ruminants. J. Parasit. Dis. 2017, 41, 1028–1033. [Google Scholar] [CrossRef]
- Moudgil, A.D.; Moudgil, P.; Asrani, R.K.; Agnihotri, R.K. Hydatidosis in slaughtered sheep and goats in India: Prevalence, genotypic characterization and pathological studies. J. Helminthol. 2020, 94, e27. [Google Scholar] [CrossRef] [Green Version]
- Haroun, E.M.; Omer, O.H.; Mahmoud, O.M.; Draz, A. Serological studies on hydatidosis in camels in Saudi Arabia. Department of Veterinary Medicine, College of Agriculture and Veterinary Medicine. Res. J. Vet. Sci. 2008, 1, 71–73. [Google Scholar]
- Thompson, R.C.A.; Lymbery, A.J. The nature, extent and significance of variation within the genus Echinococcus. Adv. Parasitol. 1988, 27, 210–258. [Google Scholar]
- Schwabe, C.W. Current status of hydatid disease: A zoonosis of increasing importance. In The biology of Echinococcus and Hydatid Disease; Thompson, R.C.A., Ed.; George Allen & Unwin: London, UK, 1988; pp. 81–113. [Google Scholar]
- Laurimäe, T.; Kinkar, L.; Romig, T.; Omer, R.A.; Casulli, A.; Umhang, G.; Gasser, R.B.; Jabbar, A.; Sharbatkhori, M.; Mirhendi, H.; et al. The benefits of analysing complete mitochondrial genomes: Deep insights into the phylogeny and population structure of Echinococcus granulosus sensu lato genotypes G6 and G7. Infect. Genet. Evol. 2018, 64, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Cardona, G.A.; Carmena, D. A review of the global prevalence, molecular epidemiology and economics of cystic echinococcosis in production animals. Vet. Parasitol. 2013, 192, 10–32. [Google Scholar] [CrossRef]
- Romig, T.; Deplazes, P.; Jenkins, D.; Giraudoux, P.; Massolo, A.; Craig, P.S.; Wassermann, M.; Takahashi, K.; de la Rue, M. Ecology and Life Cycle Patterns of Echinococcus Species. Adv. Parasitol. 2017, 95, 213–314. [Google Scholar] [PubMed]
- Siles-Lucas, M.; Casulli, A.; Conraths, F.J.; Müller, N. Laboratory diagnosis of Echinococcus spp. in human patients and infected animals. Adv. Parasitol. 2017, 96, 159–257. [Google Scholar] [PubMed]
- Carmena, D.; Benito, A.; Eraso, E. Antigens for the immunodiagnosis of Echinococcus granulosus infection. An update. Acta Trop. 2011, 98, 74–86. [Google Scholar] [CrossRef]
- Golassa, L.; Abebe, T.; Hailu, A. Evaluation of crude hydatid cyst fluid antigens for the serological diagnosis of hydatidosis in cattle. J. Helminthol. 2011, 85, 100–108. [Google Scholar] [CrossRef]
- Aboelsoued, D.; Hendawy, S.; Abdullah, H.H.A.M.; Megeed, K.N.A.; El Hakim, A.E.; Hassan, S.E.; Toaleb, N.I. Diagnosis of cryptosporidiosis using affinity-purified antigen. Egypt. J. Vet. Sci. 2022, 53, 459–473. [Google Scholar] [CrossRef]
- Zhang, W.; Li, J.; Lin, R.; Wen, H.; McManus, D.P. Immunology and Serological Diagnosis of Echinococcosis, Serological Diagnosis of Certain Human, Animal and Plant Diseases; Al-Moslih, M., Ed.; InTech: Singapore, 2012; pp. 8–150. [Google Scholar]
- Rashed, S.; Nasr, M.; Shalash, I.; Ali, N.; Kishik, S.; El-Ghanam, A. Echinococcus granulosus protoscolex antigen used in serodiagnosis of hydatidosis by nano-gold dot-ELISA. Parasitol. United J. 2019, 12, 110–115. [Google Scholar] [CrossRef]
- EL-Baz, M.A. Some Studies on Diagnosis of Hydatidosis in Slaughtered Animals. Master’s Thesis, Faculty of Veterinary Medicine Cairo University, Giza, Egypt, 1997. [Google Scholar]
- Abo-Aziza, F.A.M.; Hendawy, S.H.M.; Oda, S.S.; Aboelsoued, D.; El Shanawany, E.E. Cell-mediated and humoral immune profile to hydatidosis among naturally infected farm animals. Vet. World 2020, 13, 214–221. [Google Scholar] [CrossRef] [Green Version]
- Garedaghi, Y. Prevalence and fertility of hydatid cyst in slaughtered farm animals of Tabriz city, Iran. Life Sci. J. 2013, 10, 190–193. [Google Scholar]
- Doiz, O.; Benito, R.; Sbihi, Y.; Osuna, A.; Clavel, A.; Gomez-Lus, R. Western blot applied to the diagnosis and post-treatment monitoring of human hydatidosis. Diag. Microbiol. Infect. Dis. 2011, 41, 139–142. [Google Scholar] [CrossRef] [PubMed]
- Aboelsoued, D.; Abdullah, H.H.A.M.; Abdel Megeed, K.N.; Hassan, S.E.; Toaleb, N.I. Evaluation of a vaccine candidate isolated from Cryptosporidium parvum oocyst in mice. Vet. World 2022, 15, 2772–2784. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Blood Samples | Number |
|---|---|
| CE positive camel blood samples collected at PM inspection during several visits to abattoirs (gold standard positive control sera) | 273 |
| CE negative camel sera collected from healthy young camels, proposed to be free of cysts as confirmed by PCR, and other parasitic infections veterinary and PM inspection, and fecal examination (gold standard negative control sera) | 57 |
| Blood samples collected from slaughtered camels infected with Fasciola gigantica (Fascioliasis) in livers | 8 |
| Blood samples collected from camels infested with Hyalomma dromedarii ticks (Ticks are abundant on camels) | 9 |
| Samples collected from camels infested with Cephalopina titillator larvae at PM inspection (Myiasis, after examination of slaughtered camels’ skulls) | 20 |
| Samples from infected camels with Coccidiosis (Eimeriosis) caused by parasites of the genus Eimeria (as proved by fecal examination) | 9 |
| Infected camels’ sera with toxoplasmosis (positive anti-Toxoplasma antibodies detection in sera of camels by ELISA) | 10 |
| Blood samples from infected camels with Cryptosporidium sp. (as proved by fecal examination) | 10 |
| Blood samples from examined camels non infected with CE or other parasites | 191 |
| Total number of samples | 587 |
| Animals Examined | Infected Organs Examined | Number of Hydatid Cysts/ Examined (%) | Number of Fertile Cysts (%) | Number of Sterile Cysts | Number of Calcified Cysts (%) | Viability of Protoscoleces in Fertile Cysts (Mean ± SD) |
|---|---|---|---|---|---|---|
| Camels | Lungs | 164/273 (60%) | 137 (83.5%) | 18 (10.9%) | 9 (5.5%) | 55.23 ± 6.57 |
| Liver | 109/273 (39.9%) | 99 (90.8%) | 6 (5.5%) | 4 (3.7%) | 67.85 ± 12.78 |
| Group (No. of Camel) | PM Diagnosis | Sandwich ELISA | ||||
|---|---|---|---|---|---|---|
| Positive Number | Negative Number | Positive Serum Sample | Negative Serum Samples | |||
| No. | Optical Density Mean ± SD | No. | Optical Density Mean ± SD | |||
| Positive control sera infected with E. granulosus (n = 273) | 273 | 0 | 270 | 1.055 ± 0.21 | 3 | 0.254 ± 0.03 |
| Negative control for E. granulosus; healthy sera (n = 57) | 0 | 57 | 0 | 0 | 57 | 0.138 ± 0.020 |
| Camel Fascioliasis sera (n = 8) | 0 | 8 | 3 | 0.345 ± 0.013 | 5 | 0.237 ± 0.015 |
| Camel infested with H. dromedarii ticks (n = 9) | 0 | 9 | 0 | 0 | 9 | 0.157 ± 0.017 |
| Camel Myiasis (n = 20) | 0 | 20 | 3 | 0.358 ± 0.021 | 17 | 0.218 ± 0.022 |
| Camels with Coccidiosis (n = 9) | 0 | 9 | 0 | 0 | 9 | 0.189 ± 0.031 |
| Camel Toxoplasmosis (n = 10) | 0 | 10 | 0 | 0 | 10 | 0.197 ± 0.013 |
| Camel Cryptosporidiosis (n = 10) | 0 | 10 | 0 | 0 | 8 | 0.246 ± 0.015 |
| Camels non infected with CE or other parasites (191) | 0 | 191 | 10 | 0.699 ± 0.21 | 181 | 0.248 ± 0.023 |
| Sandwich ELISA in Detection of Echinococcus GlAg | Sensitivity% | Specificity% | AUC/p | Diagnostic Efficacy | Accuracy |
| 98.9% | 94.9% | 0.99/<0.001 | 96.8% | 96.8% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Toaleb, N.I.; Aboelsoued, D.; Abdel Megeed, K.N.; Hekal, S.H.A. A Novel Designed Sandwich ELISA for the Detection of Echinococcus granulosus Antigen in Camels for Diagnosis of Cystic Echinococcosis. Trop. Med. Infect. Dis. 2023, 8, 400. https://doi.org/10.3390/tropicalmed8080400
Toaleb NI, Aboelsoued D, Abdel Megeed KN, Hekal SHA. A Novel Designed Sandwich ELISA for the Detection of Echinococcus granulosus Antigen in Camels for Diagnosis of Cystic Echinococcosis. Tropical Medicine and Infectious Disease. 2023; 8(8):400. https://doi.org/10.3390/tropicalmed8080400
Chicago/Turabian StyleToaleb, Nagwa I., Dina Aboelsoued, Kadria N. Abdel Megeed, and Sahar Hussein Abdalla Hekal. 2023. "A Novel Designed Sandwich ELISA for the Detection of Echinococcus granulosus Antigen in Camels for Diagnosis of Cystic Echinococcosis" Tropical Medicine and Infectious Disease 8, no. 8: 400. https://doi.org/10.3390/tropicalmed8080400