
Leishmania infantum Axenic Amastigotes Induce Human Neutrophil Extracellular Traps and Resist NET-Mediated Killing
 , , and
, , and 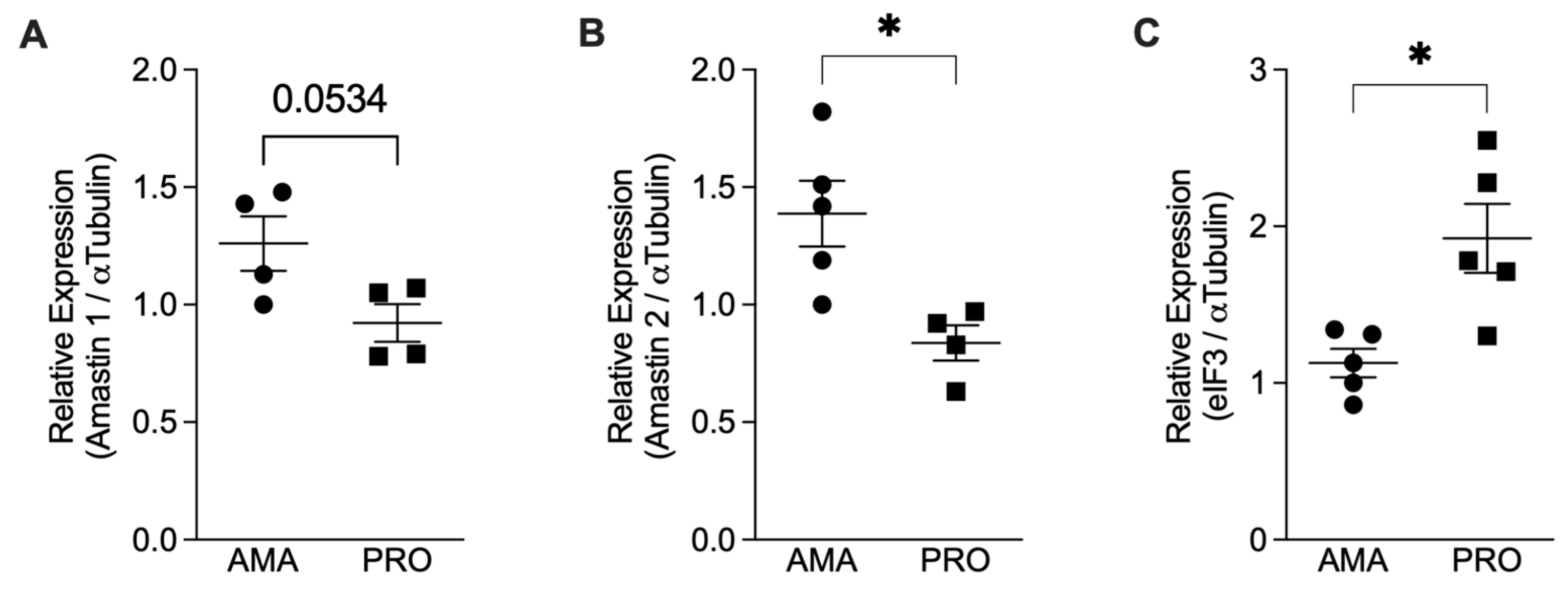{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation of Human Neutrophils
2.2. Parasite Culture
2.3. Axenic Amastigote Characterization
2.4. NET Induction Assay
2.5. Immunofluorescence
2.6. Amastigotes Labeling with Carboxyfluorescein Succinimidyl Ester (CFSE)
2.7. ROS Production
2.8. Production of NETs-Enriched Supernatants
2.9. NETs’ Toxicity to Amastigotes
2.10. Statistical Analysis
3. Results
3.1. Axenic Amastigotes Characterization
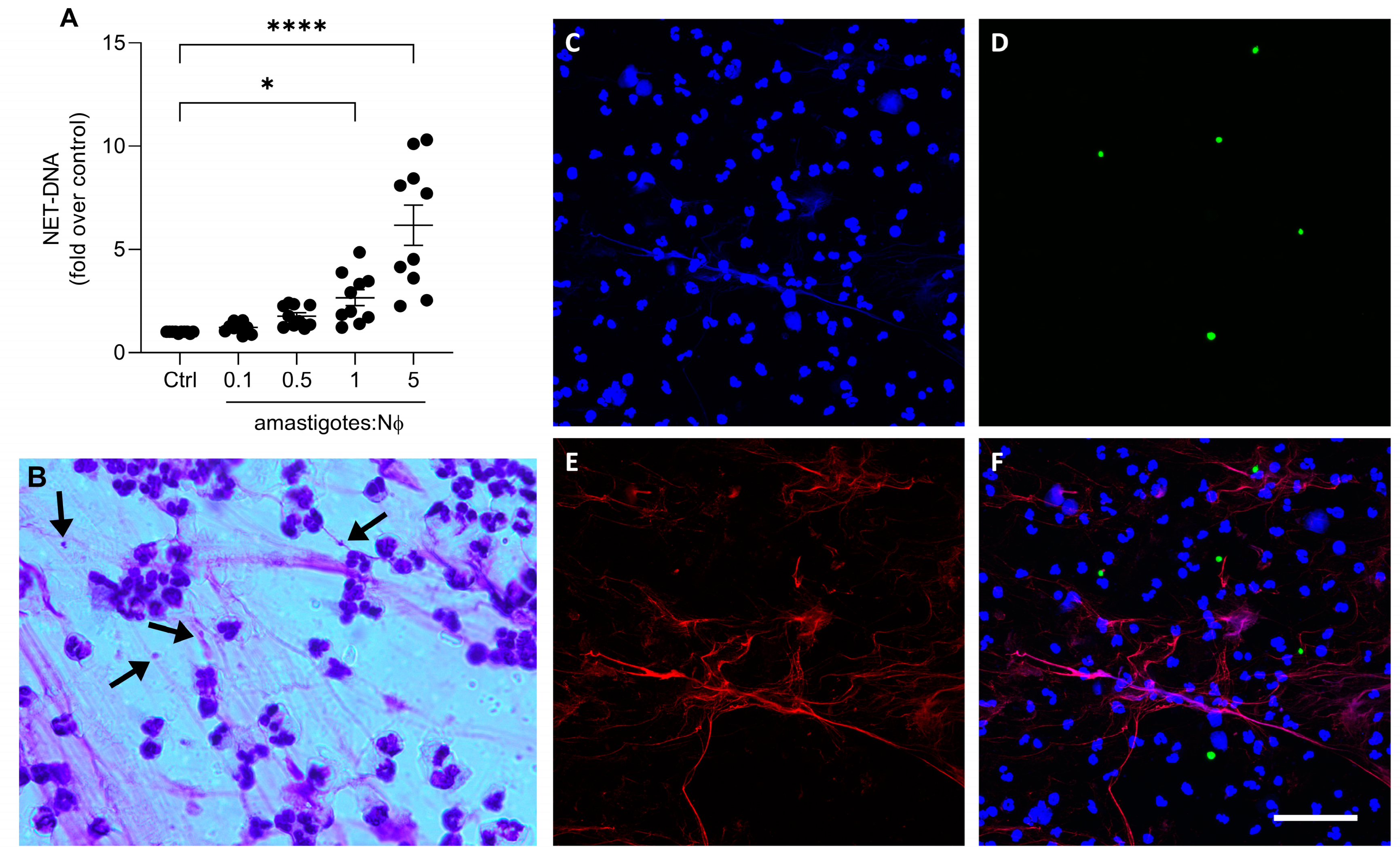
3.2. Axenic Amastigotes Induce NET Release in Human Neutrophils
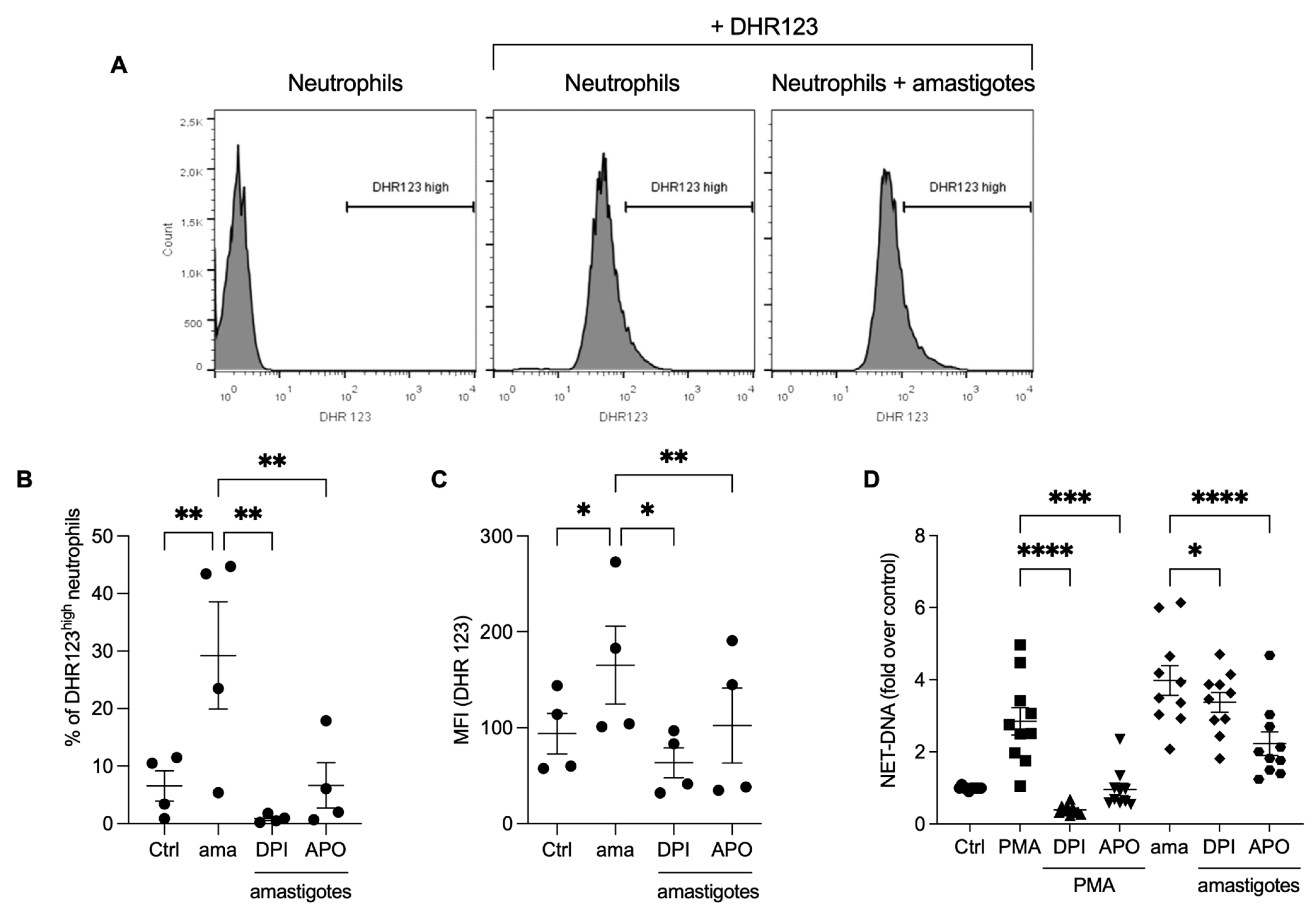
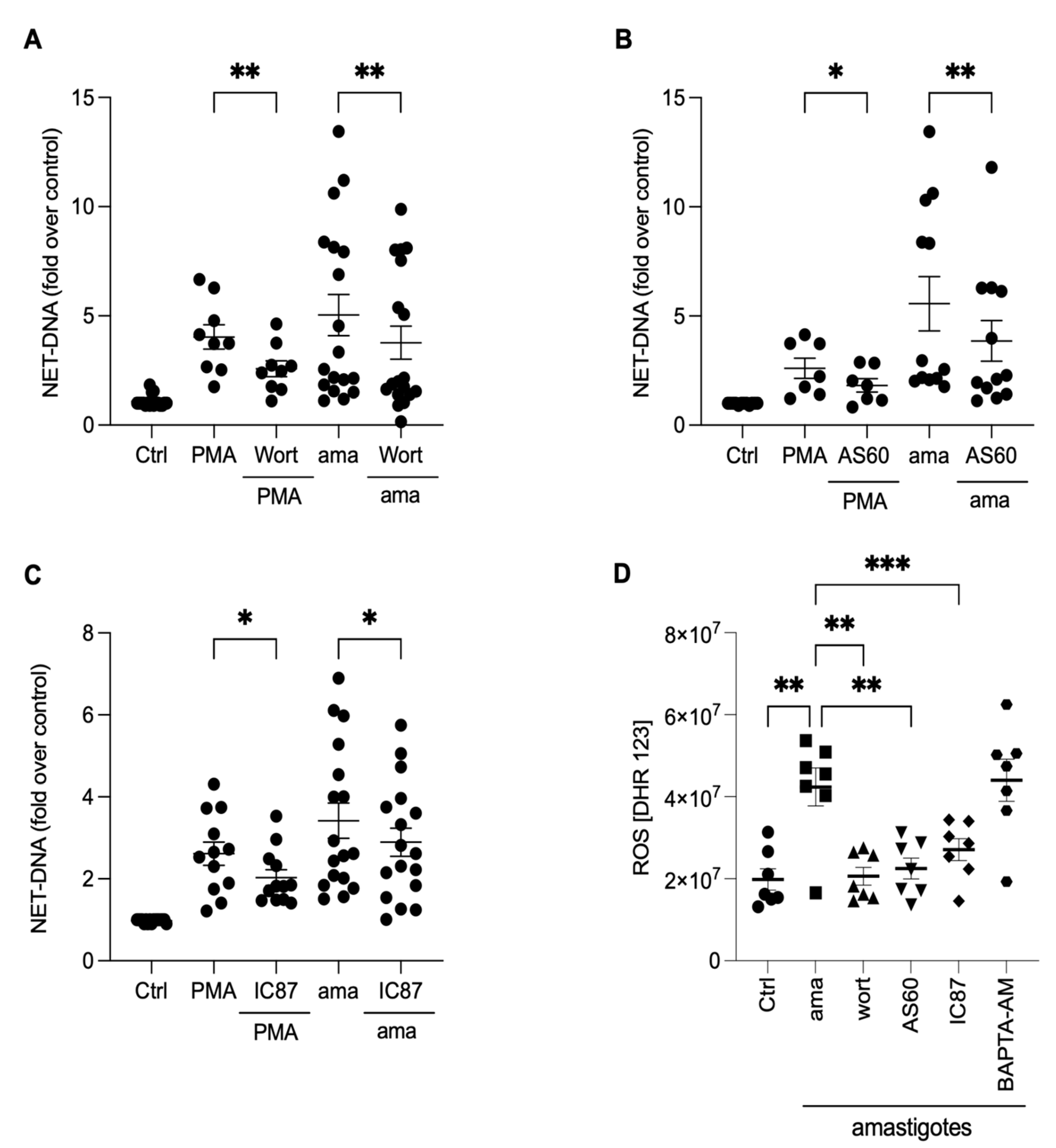
3.3. Reactive Oxygen Species (ROS) Contribute to the Release of NETs Induced by Amastigotes
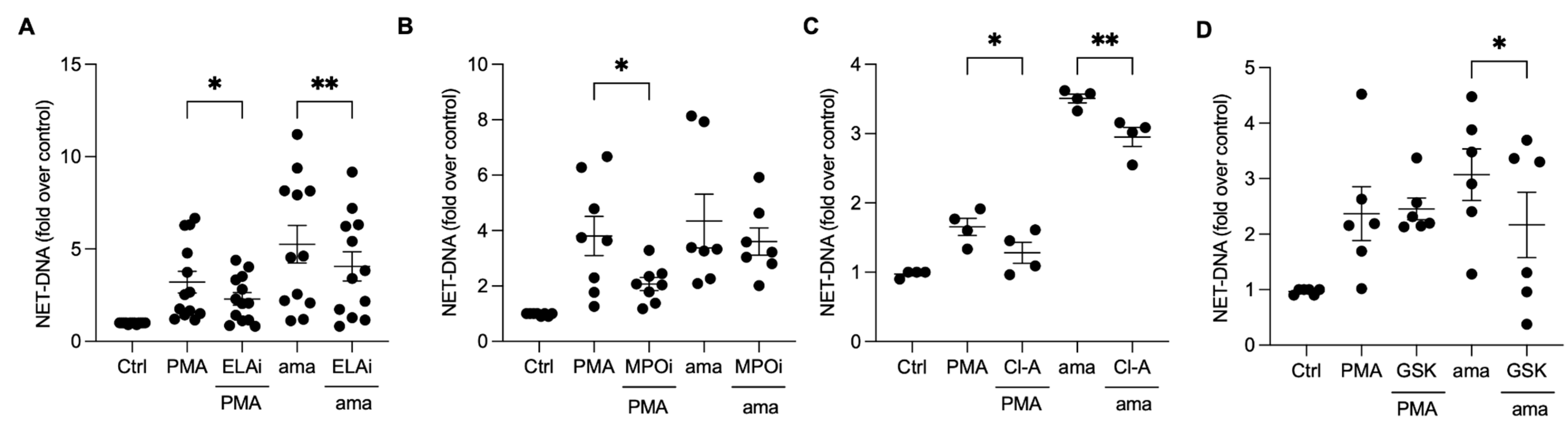
3.4. Chromatin Decondensation Promoted by Neutrophil Elastase and Peptidyl Arginine Deiminases (PAD) Contribute to Amastigotes-Induced NET Release
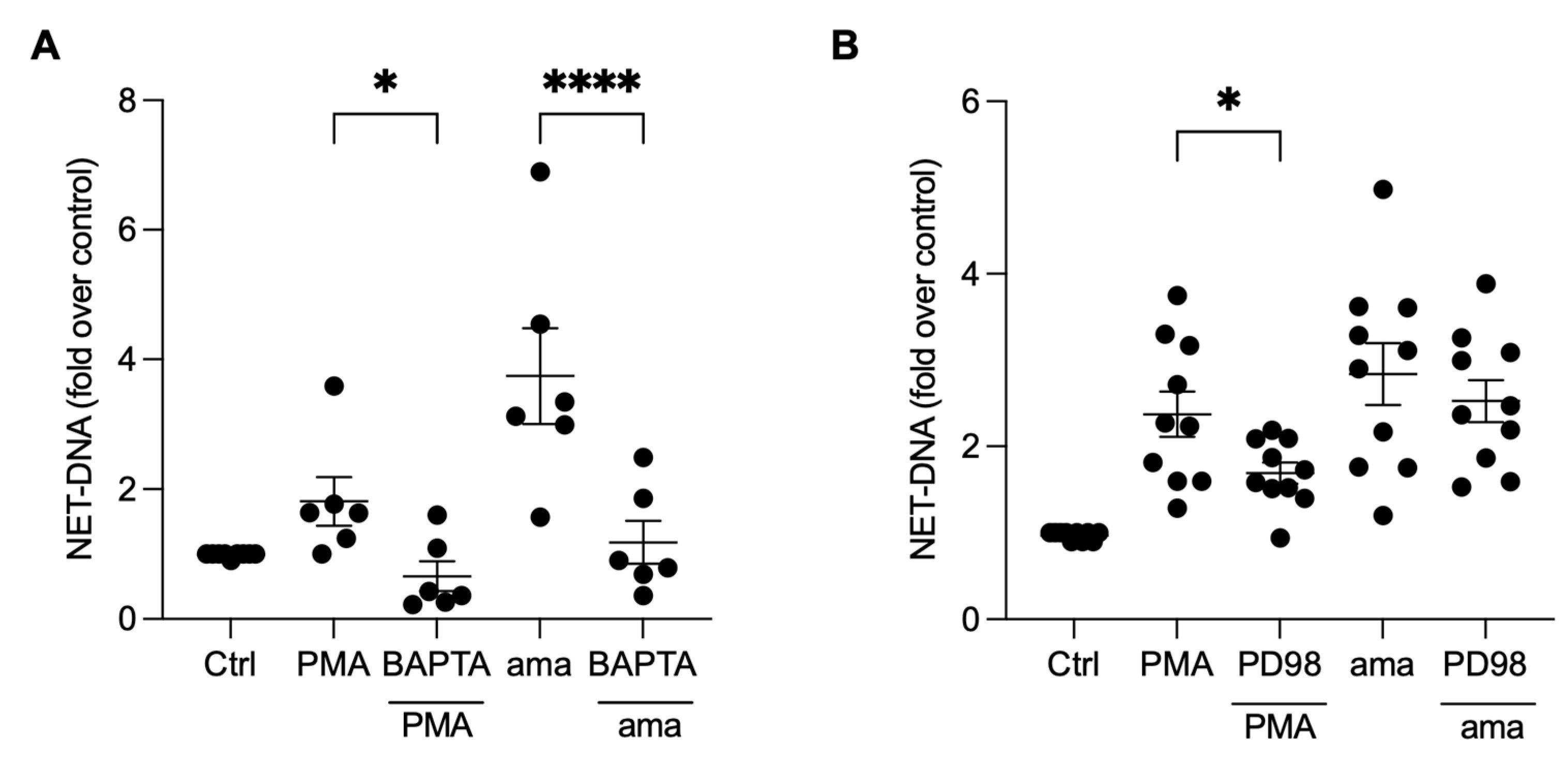
3.5. NET Generation by Amastigotes-Stimulated Neutrophils Requires Calcium, but Not ERK Signaling
3.6. PI3K Signaling Controls ROS Production and NET Extrusion by Amastigote-Stimulated Neutrophils
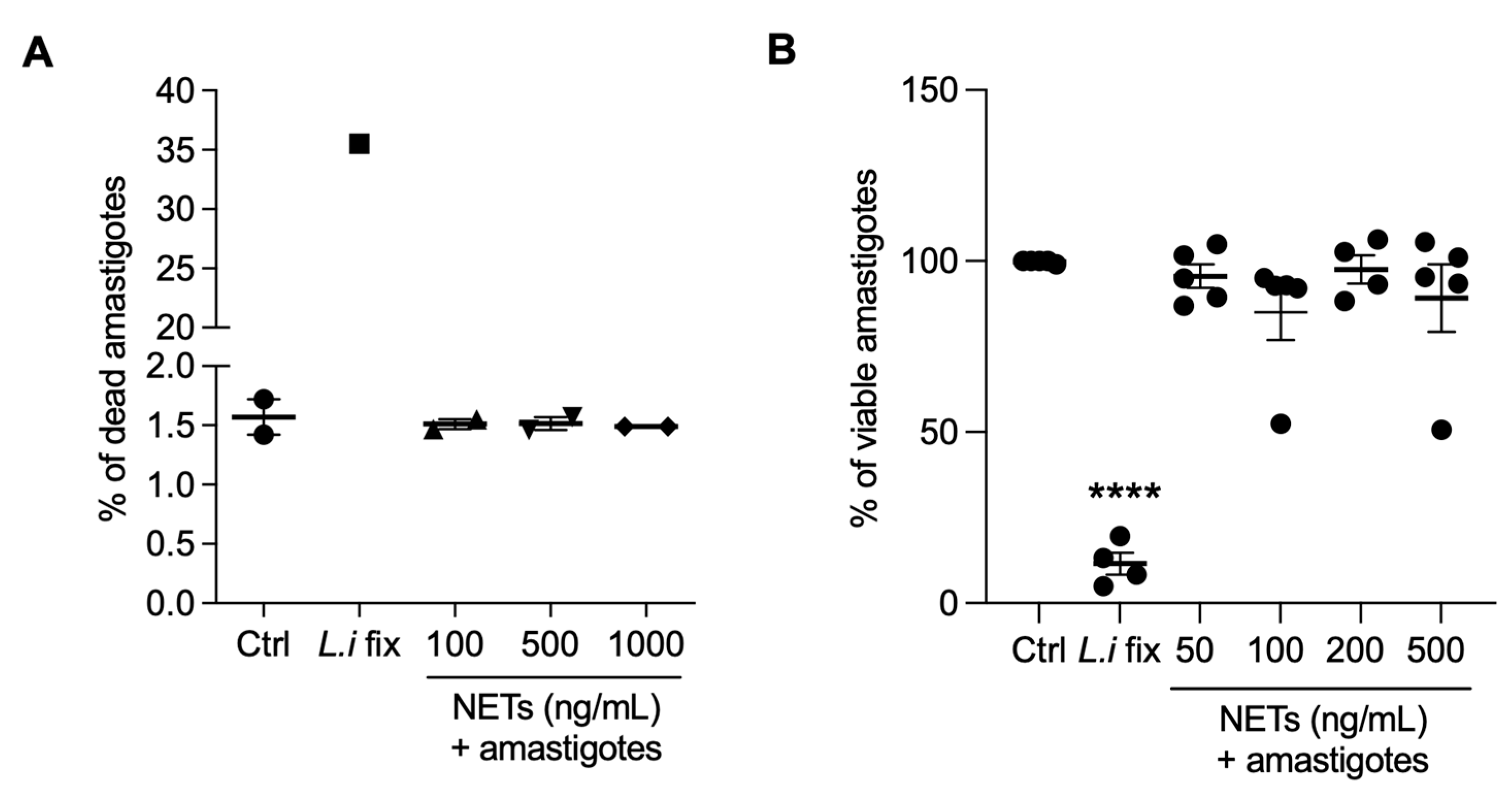
3.7. NETs Are not Toxic to Axenic Amastigotes of L. infantum
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Leishmaniasis. Available online: https://www.who.int/news-room/fact-sheets/detail/leishmaniasis (accessed on 8 May 2023).
- Fievez, A.M.d.C.; Silva-Freitas, M.L.; Sousa, A.d.Q.; Santos-Oliveira, J.R.; Da-Cruz, A.M. Lower Levels of Leptin Are Associated with Severity Parameters in Visceral Leishmaniasis Patients. PLoS ONE 2019, 14, e0214413. [Google Scholar] [CrossRef]
- Serafim, T.D.; Coutinho-Abreu, I.V.; Dey, R.; Kissinger, R.; Valenzuela, J.G.; Oliveira, F.; Kamhawi, S. Leishmaniasis: The Act of Transmission. Trends Parasitol. 2021, 37, 976–987. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, M.; Hakkim, A.; Brinkmann, V.; Siler, U.; Seger, R.A.; Zychlinsky, A.; Reichenbach, J. Restoration of NET Formation by Gene Therapy in CGD Controls Aspergillosis. Blood 2009, 114, 2619–2622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bianchi, M.; Niemiec, M.J.; Siler, U.; Urban, C.F.; Reichenbach, J. Restoration of Anti-Aspergillus Defense by Neutrophil Extracellular Traps in Human Chronic Granulomatous Disease after Gene Therapy Is Calprotectin-Dependent. J. Allergy Clin. Immunol. 2011, 127, 1243–1252.e7. [Google Scholar] [CrossRef] [PubMed]
- Brinkmann, V.; Reichard, U.; Goosmann, C.; Fauler, B.; Uhlemann, Y.; Weiss, D.S.; Weinrauch, Y.; Zychlinsky, A. Neutrophil Extracellular Traps Kill Bacteria. Science 2004, 303, 1532–1535. [Google Scholar] [CrossRef] [PubMed]
- de Jesus Gonzalez-Contreras, F.; Zarate, X. Neutrophil Extracellular Traps: Modulation Mechanisms by Pathogens. Cell Immunol. 2022, 382, 104640. [Google Scholar] [CrossRef]
- Hakkim, A.; Fürnrohr, B.G.; Amann, K.; Laube, B.; Abed, U.A.; Brinkmann, V.; Herrmann, M.; Voll, R.E.; Zychlinsky, A. Impairment of Neutrophil Extracellular Trap Degradation Is Associated with Lupus Nephritis. Proc. Natl. Acad. Sci. USA 2010, 107, 9813–9818. [Google Scholar] [CrossRef] [Green Version]
- Cahilog, Z.; Zhao, H.; Wu, L.; Alam, A.; Eguchi, S.; Weng, H.; Ma, D. The Role of Neutrophil NETosis in Organ Injury: Novel Inflammatory Cell Death Mechanisms. Inflammation 2020, 43, 2021–2032. [Google Scholar] [CrossRef]
- Oualha, R.; Barhoumi, M.; Marzouki, S.; Harigua-Souiai, E.; Ben Ahmed, M.; Guizani, I. Infection of Human Neutrophils With Leishmania Infantum or Leishmania Major Strains Triggers Activation and Differential Cytokines Release. Front. Cell. Infect. Microbiol. 2019, 9, 153. [Google Scholar] [CrossRef] [Green Version]
- Guimarães-Costa, A.B.; Nascimento, M.T.C.; Froment, G.S.; Soares, R.P.P.; Morgado, F.N.; Conceição-Silva, F.; Saraiva, E.M. Leishmania Amazonensis Promastigotes Induce and Are Killed by Neutrophil Extracellular Traps. Proc. Natl. Acad. Sci. USA 2009, 106, 6748–6753. [Google Scholar] [CrossRef] [Green Version]
- Guimarães-Costa, A.B.; DeSouza-Vieira, T.S.; Paletta-Silva, R.; Freitas-Mesquita, A.L.; Meyer-Fernandes, J.R.; Saraiva, E.M. 3′-Nucleotidase/Nuclease Activity Allows Leishmania Parasites To Escape Killing by Neutrophil Extracellular Traps. Infect. Immun. 2014, 82, 1732–1740. [Google Scholar] [CrossRef] [Green Version]
- Gabriel, C.; McMaster, W.R.; Girard, D.; Descoteaux, A. Leishmania Donovani Promastigotes Evade the Antimicrobial Activity of Neutrophil Extracellular Traps. J. Immunol. 2010, 185, 4319–4327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hurrell, B.P.; Schuster, S.; Grün, E.; Coutaz, M.; Williams, R.A.; Held, W.; Malissen, B.; Malissen, M.; Yousefi, S.; Simon, H.-U.; et al. Rapid Sequestration of Leishmania Mexicana by Neutrophils Contributes to the Development of Chronic Lesion. PLoS Pathog. 2015, 11, e1004929. [Google Scholar] [CrossRef] [PubMed]
- Wardini, A.B.; Pinto-da-Silva, L.H.; Nadaes, N.R.; Nascimento, M.T.; Roatt, B.M.; Reis, A.B.; Viana, K.F.; Giunchetti, R.C.; Saraiva, E.M. Neutrophil Properties in Healthy and Leishmania Infantum-Naturally Infected Dogs. Sci. Rep. 2019, 9, 6247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rochael, N.C.; Guimarães-Costa, A.B.; Nascimento, M.T.C.; DeSouza-Vieira, T.S.; Oliveira, M.P.; Garcia E Souza, L.F.; Oliveira, M.F.; Saraiva, E.M. Classical ROS-Dependent and Early/Rapid ROS-Independent Release of Neutrophil Extracellular Traps Triggered by Leishmania Parasites. Sci. Rep. 2015, 5, 18302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeSouza-Vieira, T.; Guimarães-Costa, A.; Rochael, N.C.; Lira, M.N.; Nascimento, M.T.; Lima-Gomez, P.d.S.; Mariante, R.M.; Persechini, P.M.; Saraiva, E.M. Neutrophil Extracellular Traps Release Induced by Leishmania: Role of PI3Kγ, ERK, PI3Kσ, PKC, and [Ca2+]. J. Leukoc. Biol. 2016, 100, 801–810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morgado, F.N.; Nascimento, M.T.C.; Saraiva, E.M.; Oliveira-Ribeiro, C.D.; Madeira, M.D.F.; Costa-Santos, M.D.; Vasconcellos, E.C.F.; Pimentel, M.I.F.; Rosandiski Lyra, M.; Schubach, A.D.O.; et al. Are Neutrophil Extracellular Traps Playing a Role in the Parasite Control in Active American Tegumentary Leishmaniasis Lesions? PLoS ONE 2015, 10, e0133063. [Google Scholar] [CrossRef] [Green Version]
- Gardinassi, L.G.; DeSouza-Vieira, T.S.; da Silva, N.O.; Garcia, G.R.; Borges, V.M.; Campos, R.N.S.; de Almeida, R.P.; de Miranda Santos, I.K.F.; Saraiva, E.M. Molecular Signatures of Neutrophil Extracellular Traps in Human Visceral Leishmaniasis. Parasites Vectors 2017, 10, 285. [Google Scholar] [CrossRef]
- Carlsen, E.D.; Hay, C.; Henard, C.A.; Popov, V.; Garg, N.J.; Soong, L. Leishmania Amazonensis Amastigotes Trigger Neutrophil Activation but Resist Neutrophil Microbicidal Mechanisms. Infect. Immun. 2013, 81, 3966–3974. [Google Scholar] [CrossRef] [Green Version]
- Rocco-Machado, N.; Cosentino-Gomes, D.; Nascimento, M.T.; Paes-Vieira, L.; Khan, Y.A.; Mittra, B.; Andrews, N.W.; Meyer-Fernandes, J.R. Leishmania Amazonensis Ferric Iron Reductase (LFR1) Is a Bifunctional Enzyme: Unveiling a NADPH Oxidase Activity. Free. Radic. Biol. Med. 2019, 143, 341–353. [Google Scholar] [CrossRef]
- Meleppattu, S.; Kamus-Elimeleh, D.; Zinoviev, A.; Cohen-Mor, S.; Orr, I.; Shapira, M. The EIF3 Complex of Leishmania —Subunit Composition and Mode of Recruitment to Different Cap-Binding Complexes. Nucleic Acids Res. 2015, 43, 6222–6235. [Google Scholar] [CrossRef] [Green Version]
- Gupta, A.K.; Giaglis, S.; Hasler, P.; Hahn, S. Efficient Neutrophil Extracellular Trap Induction Requires Mobilization of Both Intracellular and Extracellular Calcium Pools and Is Modulated by Cyclosporine A. PLoS ONE 2014, 9, e97088. [Google Scholar] [CrossRef] [Green Version]
- Hawkins, P.T.; Davidson, K.; Stephens, L.R. The Role of PI3Ks in the Regulation of the Neutrophil NADPH Oxidase. Biochem. Soc. Symp. 2007, 74, 59. [Google Scholar] [CrossRef]
- Wardini, A.B.; Guimaraes-Costa, A.B.; Nascimento, M.T.C.; Nadaes, N.R.; Danelli, M.G.M.; Mazur, C.; Benjamim, C.F.; Saraiva, E.M.; Pinto-da-Silva, L.H. Characterization of Neutrophil Extracellular Traps in Cats Naturally Infected with Feline Leukemia Virus. J. Gen. Virol. 2010, 91, 259–264. [Google Scholar] [CrossRef]
- Hurrell, B.P.; Regli, I.B.; Tacchini-Cottier, F. Different Leishmania Species Drive Distinct Neutrophil Functions. Trends Parasitol. 2016, 32, 392–401. [Google Scholar] [CrossRef] [PubMed]
- Wei, F.; Gong, W.; Wang, J.; Yang, Y.; Liu, J.; Wang, Y.; Cao, J. Role of the Lipoxin A4 Receptor in the Development of Neutrophil Extracellular Traps in Leishmania Infantum Infection. Parasites Vectors 2019, 12, 275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, A.V.A.; Souza, T.L.; Figueiredo, F.B.; Mendes, A.A.V.; Ferreira, L.C.; Filgueira, C.P.B.; Cuervo, P.; Porrozzi, R.; Menezes, R.C.; Morgado, F.N. Detection of Amastigotes and Histopathological Alterations in the Thymus of Leishmania Infantum -infected Dogs. Immun. Inflamm. Dis. 2020, 8, 127–139. [Google Scholar] [CrossRef] [Green Version]
- Kost, W.D.O.; Pereira, S.A.; Figueiredo, F.B.; Mendes Junior, A.A.V.; Madeira, M.D.F.; Miranda, L.D.F.C.; De Oliveira, R.D.V.C.; Ferreira, L.C.; Morgado, F.N.; Menezes, R.C. Frequency of Detection and Load of Amastigotes in the Pancreas of Leishmania Infantum-Seropositive Dogs: Clinical Signs and Histological Changes. Parasites Vectors 2021, 14, 321. [Google Scholar] [CrossRef] [PubMed]
- Rochette, A.; McNicoll, F.; Girard, J.; Breton, M.; Leblanc, É.; Bergeron, M.G.; Papadopoulou, B. Characterization and Developmental Gene Regulation of a Large Gene Family Encoding Amastin Surface Proteins in Leishmania spp. Mol. Biochem. Parasitol. 2005, 140, 205–220. [Google Scholar] [CrossRef]
- He, J.; Huang, F.; Li, J.; Chen, Q.; Chen, D.; Chen, J. Bioinformatics Analysis of Four Proteins of Leishmania Donovani to Guide Epitopes Vaccine Design and Drug Targets Selection. Acta Trop. 2019, 191, 50–59. [Google Scholar] [CrossRef]
- Coelho, V.T.S.; Oliveira, J.S.; Valadares, D.G.; Chávez-Fumagalli, M.A.; Duarte, M.C.; Lage, P.S.; Soto, M.; Santoro, M.M.; Tavares, C.A.P.; Fernandes, A.P.; et al. Identification of Proteins in Promastigote and Amastigote-like Leishmania Using an Immunoproteomic Approach. PLoS Negl. Trop. Dis. 2012, 6, e1430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuchs, T.A.; Abed, U.; Goosmann, C.; Hurwitz, R.; Schulze, I.; Wahn, V.; Weinrauch, Y.; Brinkmann, V.; Zychlinsky, A. Novel Cell Death Program Leads to Neutrophil Extracellular Traps. J. Cell Biol. 2007, 176, 231–241. [Google Scholar] [CrossRef] [Green Version]
- Papayannopoulos, V.; Metzler, K.D.; Hakkim, A.; Zychlinsky, A. Neutrophil Elastase and Myeloperoxidase Regulate the Formation of Neutrophil Extracellular Traps. J. Cell Biol. 2010, 191, 677–691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leshner, M.; Wang, S.; Lewis, C.; Zheng, H.; Chen, X.A.; Santy, L.; Wang, Y. PAD4 Mediated Histone Hypercitrullination Induces Heterochromatin Decondensation and Chromatin Unfolding to Form Neutrophil Extracellular Trap-like Structures. Front. Immunol. 2012, 3, 307. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Li, M.; Stadler, S.; Correll, S.; Li, P.; Wang, D.; Hayama, R.; Leonelli, L.; Han, H.; Grigoryev, S.A.; et al. Histone Hypercitrullination Mediates Chromatin Decondensation and Neutrophil Extracellular Trap Formation. J. Cell Biol. 2009, 184, 205–213. [Google Scholar] [CrossRef] [Green Version]
- Kenny, E.F.; Herzig, A.; Krüger, R.; Muth, A.; Mondal, S.; Thompson, P.R.; Brinkmann, V.; Bernuth, H.V.; Zychlinsky, A. Diverse Stimuli Engage Different Neutrophil Extracellular Trap Pathways. eLife 2017, 6, e24437. [Google Scholar] [CrossRef] [PubMed]
- Metzler, K.D.; Fuchs, T.A.; Nauseef, W.M.; Reumaux, D.; Roesler, J.; Schulze, I.; Wahn, V.; Papayannopoulos, V.; Zychlinsky, A. Myeloperoxidase Is Required for Neutrophil Extracellular Trap Formation: Implications for Innate Immunity. Blood 2011, 117, 953–959. [Google Scholar] [CrossRef] [Green Version]
- Metzler, K.D.; Goosmann, C.; Lubojemska, A.; Zychlinsky, A.; Papayannopoulos, V. A Myeloperoxidase-Containing Complex Regulates Neutrophil Elastase Release and Actin Dynamics during NETosis. Cell Rep. 2014, 8, 883–896. [Google Scholar] [CrossRef] [Green Version]
- Hamam, H.J.; Palaniyar, N. Post-Translational Modifications in NETosis and NETs-Mediated Diseases. Biomolecules 2019, 9, 369. [Google Scholar] [CrossRef] [Green Version]
- Hamam, H.; Khan, M.; Palaniyar, N. Histone Acetylation Promotes Neutrophil Extracellular Trap Formation. Biomolecules 2019, 9, 32. [Google Scholar] [CrossRef] [Green Version]
- Hamam, H.J.; Palaniyar, N. Histone Deacetylase Inhibitors Dose-Dependently Switch Neutrophil Death from NETosis to Apoptosis. Biomolecules 2019, 9, 184. [Google Scholar] [CrossRef] [Green Version]
- Poli, V.; Pui-Yan Ma, V.; Di Gioia, M.; Broggi, A.; Benamar, M.; Chen, Q.; Mazitschek, R.; Haggarty, S.J.; Chatila, T.A.; Karp, J.M.; et al. Zinc-Dependent Histone Deacetylases Drive Neutrophil Extracellular Trap Formation and Potentiate Local and Systemic Inflammation. iScience 2021, 24, 103256. [Google Scholar] [CrossRef] [PubMed]
- Behnen, M.; Leschczyk, C.; Möller, S.; Batel, T.; Klinger, M.; Solbach, W.; Laskay, T. Immobilized Immune Complexes Induce Neutrophil Extracellular Trap Release by Human Neutrophil Granulocytes via FcγRIIIB and Mac-1. J. Immunol. 2014, 193, 1954–1965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Der Linden, M.; Westerlaken, G.H.A.; Van Der Vlist, M.; Van Montfrans, J.; Meyaard, L. Differential Signalling and Kinetics of Neutrophil Extracellular Trap Release Revealed by Quantitative Live Imaging. Sci. Rep. 2017, 7, 6529. [Google Scholar] [CrossRef]
- Tatsiy, O.; McDonald, P.P. Physiological Stimuli Induce PAD4-Dependent, ROS-Independent NETosis, With Early and Late Events Controlled by Discrete Signaling Pathways. Front. Immunol. 2018, 9, 2036. [Google Scholar] [CrossRef] [Green Version]
- De Assis, R.R.; Ibraim, I.C.; Nogueira, P.M.; Soares, R.P.; Turco, S.J. Glycoconjugates in New World Species of Leishmania: Polymorphisms in Lipophosphoglycan and Glycoinositolphospholipids and Interaction with Hosts. Biochim. et Biophys. Acta Gen. Subj. 2012, 1820, 1354–1365. [Google Scholar] [CrossRef]
- Hann, J.; Bueb, J.-L.; Tolle, F.; Bréchard, S. Calcium Signaling and Regulation of Neutrophil Functions: Still a Long Way to Go. J. Leukoc. Biol. 2020, 107, 285–297. [Google Scholar] [CrossRef]
- Khazen, R.; Corre, B.; Garcia, Z.; Lemaître, F.; Bachellier-Bassi, S.; d’Enfert, C.; Bousso, P. Spatiotemporal Dynamics of Calcium Signals during Neutrophil Cluster Formation. Proc. Natl. Acad. Sci. USA 2022, 119, e2203855119. [Google Scholar] [CrossRef]
- Parker, H.; Dragunow, M.; Hampton, M.B.; Kettle, A.J.; Winterbourn, C.C. Requirements for NADPH Oxidase and Myeloperoxidase in Neutrophil Extracellular Trap Formation Differ Depending on the Stimulus. J. Leukoc. Biol. 2012, 92, 841–849. [Google Scholar] [CrossRef]
- Douda, D.N.; Khan, M.A.; Grasemann, H.; Palaniyar, N. SK3 Channel and Mitochondrial ROS Mediate NADPH Oxidase-Independent NETosis Induced by Calcium Influx. Proc. Natl. Acad. Sci. USA 2015, 112, 2817–2822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, C.; Liu, C.; Chai, Z. Role of the PADI family in inflammatory autoimmune diseases and cancers: A systematic review. Front. Immunol. 2023, 14, 1115794. [Google Scholar] [CrossRef]
- Urban, C.F.; Reichard, U.; Brinkmann, V.; Zychlinsky, A. Neutrophil Extracellular Traps Capture and Kill Candida Albicans Yeast and Hyphal Forms. Cell Microbiol. 2006, 8, 668–676. [Google Scholar] [CrossRef] [PubMed]
- Saitoh, T.; Komano, J.; Saitoh, Y.; Misawa, T.; Takahama, M.; Kozaki, T.; Uehata, T.; Iwasaki, H.; Omori, H.; Yamaoka, S.; et al. Neutrophil Extracellular Traps Mediate a Host Defense Response to Human Immunodeficiency Virus-1. Cell Host Microbe 2012, 12, 109–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chagas, A.C.; Oliveira, F.; Debrabant, A.; Valenzuela, J.G.; Ribeiro, J.M.C.; Calvo, E. Lundep, a Sand Fly Salivary Endonuclease Increases Leishmania Parasite Survival in Neutrophils and Inhibits XIIa Contact Activation in Human Plasma. PLoS Pathog. 2014, 10, e1003923. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chen, Y.; Xin, L.; Beverley, S.M.; Carlsen, E.D.; Popov, V.; Chang, K.-P.; Wang, M.; Soong, L. Differential Microbicidal Effects of Human Histone Proteins H2A and H2B on Leishmania Promastigotes and Amastigotes. Infect. Immun. 2011, 79, 1124–1133. [Google Scholar] [CrossRef] [PubMed] [Green Version]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oliveira, T.K.F.; Oliveira-Silva, J.; Linhares-Lacerda, L.; da Silva Fraga-Junior, V.; Benjamim, C.F.; Guimaraes-Costa, A.B.; Saraiva, E.M. Leishmania infantum Axenic Amastigotes Induce Human Neutrophil Extracellular Traps and Resist NET-Mediated Killing. Trop. Med. Infect. Dis. 2023, 8, 336. https://doi.org/10.3390/tropicalmed8070336
Oliveira TKF, Oliveira-Silva J, Linhares-Lacerda L, da Silva Fraga-Junior V, Benjamim CF, Guimaraes-Costa AB, Saraiva EM. Leishmania infantum Axenic Amastigotes Induce Human Neutrophil Extracellular Traps and Resist NET-Mediated Killing. Tropical Medicine and Infectious Disease. 2023; 8(7):336. https://doi.org/10.3390/tropicalmed8070336
Chicago/Turabian StyleOliveira, Thamara K. F., Jullyanna Oliveira-Silva, Leandra Linhares-Lacerda, Vanderlei da Silva Fraga-Junior, Claudia F. Benjamim, Anderson B. Guimaraes-Costa, and Elvira M. Saraiva. 2023. "Leishmania infantum Axenic Amastigotes Induce Human Neutrophil Extracellular Traps and Resist NET-Mediated Killing" Tropical Medicine and Infectious Disease 8, no. 7: 336. https://doi.org/10.3390/tropicalmed8070336