
Pathogenic Leptospira Species in Bats: Molecular Detection in a Colombian Cave

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
3. Results
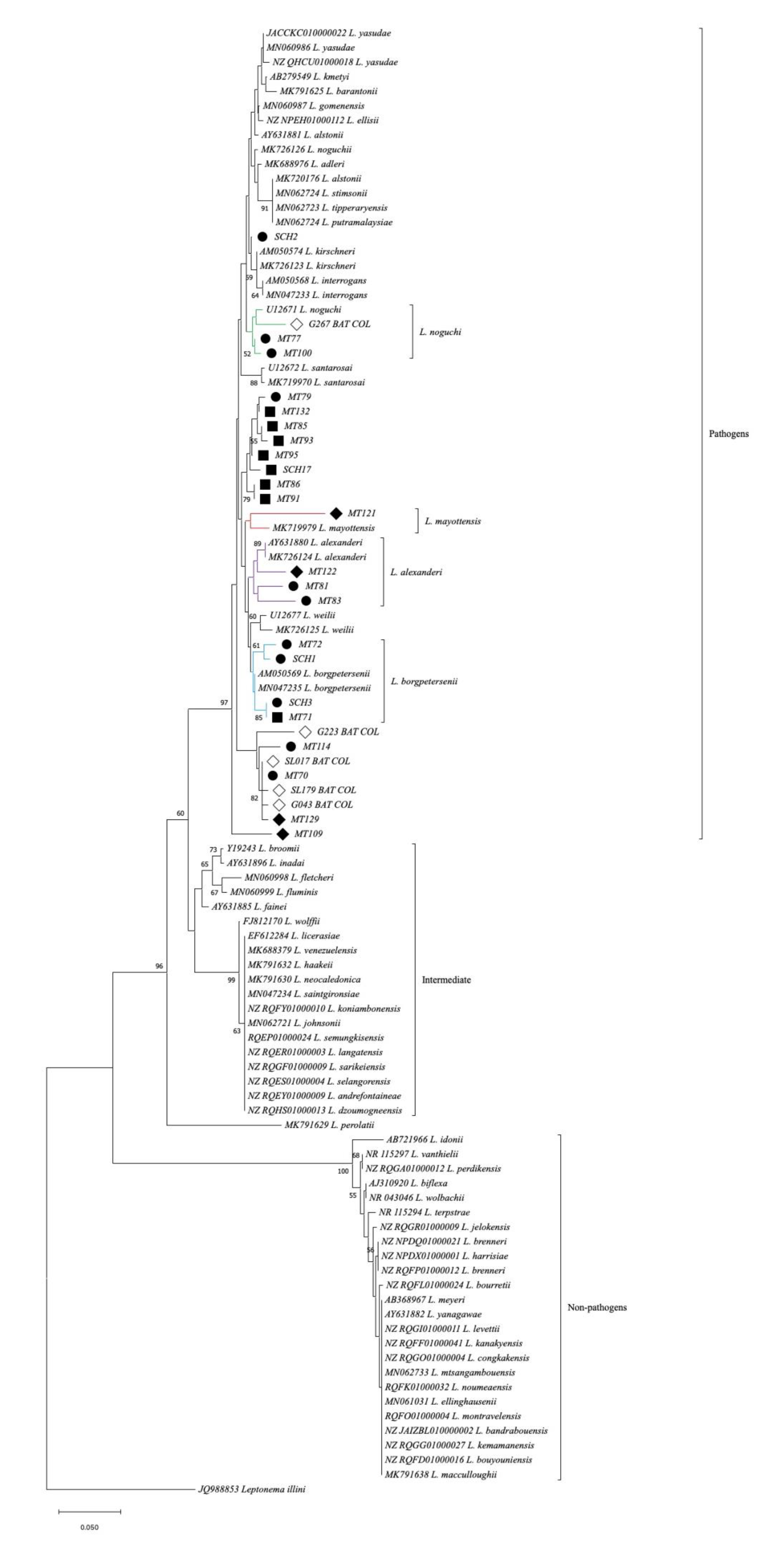
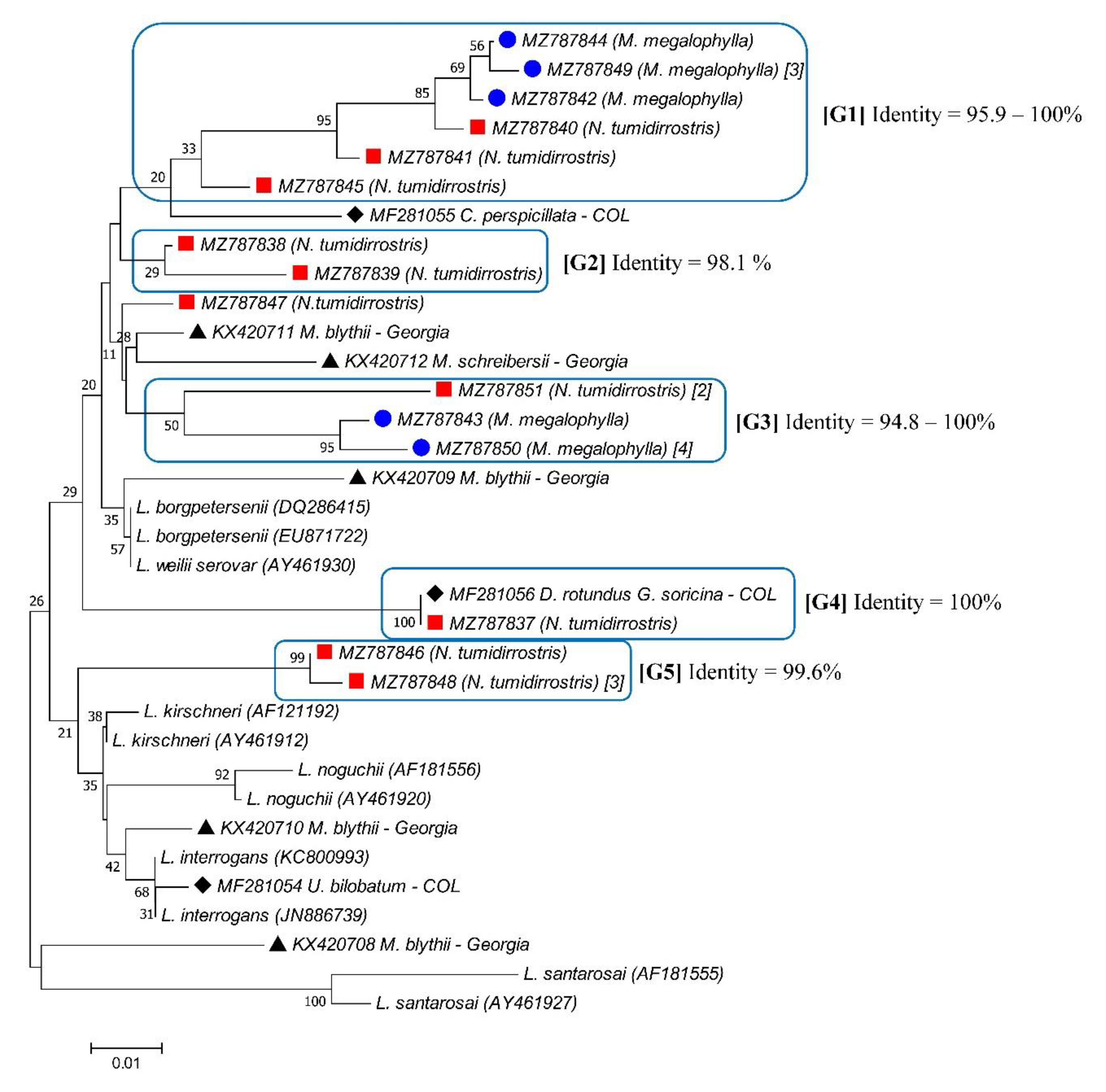
3.1. Detection of Leptospira spp.
3.2. Detection of Pathogenic Leptospira spp.
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Adler, B.; de la Peña Moctezuma, A. Leptospira and leptospirosis. Vet. Microbiol. 2010, 140, 287–296. [Google Scholar] [CrossRef]
- Picardeau, M. Leptospira. In Bergey’s Manual of Systematics of Archaea and Bacteria; Trujillo, M.E., Dedysh, S., DeVos, P., Hedlund, B., Kämpfer, P., Rainey, F.A., Whitman, W.B., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2017. [Google Scholar] [CrossRef]
- Karpagam, K.B.; Ganesh, B. Leptospirosis: A neglected tropical zoonotic infection of public health importance—An updated review. Eur. J. Clin. Microbiol. Infect. Dis. 2020, 39, 835–846. [Google Scholar] [CrossRef] [PubMed]
- Haake, D.A.; Levett, P.N. Leptospirosis in Humans. In Leptospira and Leptospirosis; Adler, B., Ed.; Springer: Berlin/Heidelberg, Germany, 2015; pp. 65–97. [Google Scholar] [CrossRef] [Green Version]
- Costa, F.; Hagan, J.E.; Calcagno, J.; Kane, M.; Torgerson, P.; Martinez-Silveira, M.S.; Stein, C.; Abela-Ridder, B.; Ko, A.I. Global Morbidity and Mortality of Leptospirosis: A Systematic Review. PLoS Negl. Trop. Dis. 2015, 9, e0003898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellis, W.A. Animal Leptospirosis. In Leptospira and Leptospirosis; Adler, B., Ed.; Springer: Berlin/Heidelberg, Germany, 2015; pp. 99–137. [Google Scholar] [CrossRef]
- Hassell, J.M.; Begon, M.; Ward, M.J.; Fèvre, E.M. Urbanization and Disease Emergence: Dynamics at the Wildlife-Livestock-Human Interface. Trends Ecol. Evol. 2017, 32, 55–67. [Google Scholar] [CrossRef] [Green Version]
- Kasso, M.; Balakrishnan, M. Ecological and Economic Importance of Bats (Order Chiroptera). Hindawi Publ. Corp. 2013, 2013, 187415. [Google Scholar] [CrossRef] [Green Version]
- Kunz, T.H.; Braun de Torrez, E.; Bauer, D.; Lobova, T.; Fleming, T.H. Ecosystem services provided by bats. Ann. N. Y. Acad. Sci. 2011, 1223, 1–38. [Google Scholar] [CrossRef] [PubMed]
- Kerth, G. Causes and Consequences of Sociality in Bats. BioScience 2008, 58, 737–746. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, A.; Baker, M.L.; Kulcsar, K.; Misra, V.; Plowright, R.; Mossman, K. Novel Insights Into Immune Systems of Bats. Front. Immunol. 2020, 11, 26. [Google Scholar] [CrossRef]
- Han, H.J.; Wen, H.L.; Zhou, C.M.; Chen, F.F.; Luo, L.M.; Liu, J.W.; Yu, X.J. Bats as reservoirs of severe emerging infectious diseases. Virus Res. 2015, 205, 1–6. [Google Scholar] [CrossRef]
- Irving, A.T.; Ahn, M.; Goh, G.; Anderson, D.E.; Wang, L.-F. Lessons from the host defences of bats, a unique viral reservoir. Nature 2021, 589, 363–370. [Google Scholar] [CrossRef]
- Mühldorfer, K. Bats and bacterial pathogens: A review. Zoonoses Public Health 2013, 60, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Kingston, T.; Aguirre, L.; Armstrong, K.; Mies, R.; Racey, P.; Rodríguez-Herrera, B.; Waldien, D. Networking Networks for Global Bat Conservation. In Bats in the Anthropocene: Conservation of Bats in a Changing World; Voigt, C., Kingston, T., Eds.; Springer: Cham, Switzerland. [CrossRef] [Green Version]
- Aguirre, L.F.; Nassar, J.M.; Barquez, R.M.; Medellín, R.A.; Navarro, L.; Rodríguez-Durán, A. De esfuerzos locales a una iniciativa regional: La Red Latinoamericana y del Caribe para la Conservación de los Murciélagos (RELCOM). Ecol. Boliv. 2014, 49, 45–50. [Google Scholar]
- Ramírez-Chaves, H.E.; Suárez Castro, A.F.; Morales-Martínez, D.M.; Rodríguez-Posada, M.E.; Zurc, D.; Concha Osbahr, D.C.; Trujillo, A.; Noguera Urbano, E.A.; Pantoja Peña, G.E.; González Maya, J.F.; et al. Mamíferos de Colombia. Sociedad Colombiana de Mastozoología. Dataset/Checklist 2021, 1, 12. [Google Scholar] [CrossRef]
- Peñuela, M.; Perez-Torres, J. Environmental and spatial characteristics that affect roost use by Seba’s short-tailed bat (Carollia Perspicillata) in a Colombian cave. J. Cave Karst Stud. 2015, 77, 160–164. [Google Scholar] [CrossRef]
- Pérez-Torres, J.; Martínez-Medina, D.; Peñuela, M.; Rios-Blanco, C.; Estrada-Villegas, S.; Martínez-Luque, L. Macaregua: The cave with the highest bat richness in Colombia. Check List 2015, 11, 1616. [Google Scholar] [CrossRef]
- Victoria, R.J.; Iriarte, L.J.; Sampedro, A.C. Presence of Leptospire spp. in urban bats from Sincelejo, Sucre, Colombia. Int. J. PharmTech Res. 2018, 11, 218–225. [Google Scholar] [CrossRef]
- Mateus, J.; Gómez, N.; Herrera-Sepúlveda, M.T.; Hidalgo, M.; Pérez-Torres, J.; Cuervo, C. Bats are a potential reservoir of pathogenic Leptospira species in Colombia. J. Infect. Dev. Ctries. 2019, 13, 278–283. [Google Scholar] [CrossRef] [Green Version]
- Monroy, F.P.; Solari, S.; Lopez, J.Á.; Agudelo-Flórez, P.; Peláez Sánchez, R.G. High Diversity of Leptospira Species Infecting Bats Captured in the Urabá Region (Antioquia-Colombia). Microorganisms 2021, 9, 1897. [Google Scholar] [CrossRef]
- Muñoz-Leal, S.; Faccini-Martínez, Á.A.; Pérez-Torres, J.; Chala-Quintero, S.M.; Herrera-Sepúlveda, M.T.; Cuervo, C.; Labruna, M.B. Novel Borrelia genotypes in bats from the Macaregua Cave, Colombia. Zoonoses Public Health 2021, 68, 12–18. [Google Scholar] [CrossRef]
- Schlegel, M.; Ali, H.S.; Stieger, N.; Groschup, M.H.; Wolf, R.; Ulrich, R.G. Molecular identification of small mammal species using novel cytochrome B gene-derived degenerated primers. Biochem. Genet. 2012, 50, 440–447. [Google Scholar] [CrossRef]
- Bessa, T.A.; Spichler, A.; Chapola, E.G.; Husch, A.C.; de Almeida, M.F.; Sodré, M.M.; Savani, E.S.; Sacramento, D.R.; Vinetz, J.M. The contribution of bats to leptospirosis transmission in Sao Paulo City, Brazil. Am. J. Trop. Med. Hyg. 2010, 82, 315–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levett, P.N.; Morey, R.E.; Galloway, R.L.; Turner, D.E.; Steigerwalt, A.G.; Mayer, L.W. Detection of pathogenic leptospires by real-time quantitative PCR. J. Med. Microbiol. 2005, 54, 45–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.W.; Wu, M.S.; Pan, M.J.; Hsieh, W.J.; Vandewalle, A.; Huang, C.C. The Leptospira outer membrane protein LipL32 induces tubulointerstitial nephritis-mediated gene expression in mouse proximal tubule cells. J. Am. Soc. Nephrol. 2002, 13, 2037–2045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef] [PubMed]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Bai, Y.; Urushadze, L.; Osikowicz, L.; McKee, C.; Kuzmin, I.; Kandaurov, A.; Babuadze, G.; Natradze, I.; Imnadze, P.; Kosoy, M. Molecular Survey of Bacterial Zoonotic Agents in Bats from the Country of Georgia (Caucasus). PLoS ONE. 2017, 12, e0171175. [Google Scholar] [CrossRef]
- Dietrich, M.; Mühldorfer, K.; Tortosa, P.; Markotter, W. Leptospira and Bats: Story of an Emerging Friendship. PLoS Pathog. 2015, 11, e1005176. [Google Scholar] [CrossRef]
- Matthias, M.A.; Díaz, M.M.; Campos, K.J.; Calderon, M.; Willig, M.R.; Pacheco, V.; Gotuzzo, E.; Gilman, R.H.; Vinetz, J.M. Diversity of bat-associated Leptospira in the Peruvian Amazon inferred by bayesian phylogenetic analysis of 16S ribosomal DNA sequences. Am. J. Trop. Med. Hyg. 2005, 73, 964–974. [Google Scholar] [CrossRef]
- Haake, D.A.; Chao, G.; Zuerner, R.L.; Barnett, J.K.; Barnett, D.; Mazel, M.; Matsunaga, J.; Levett, P.N.; Bolin, C.A. The leptospiral major outer membrane protein LipL32 is a lipoprotein expressed during mammalian infection. Infect. Immun. 2000, 68, 2276–2285. [Google Scholar] [CrossRef] [Green Version]
- Seidlova, V.; Nemcova, M.; Pikula, J.; Bartonička, T.; Ghazaryan, A.; Heger, T.; Kokurewicz, T.; Orlov, O.L.; Patra, S.; Piacek, V.; et al. Urinary shedding of leptospires in palearctic bats. Transbound. Emerg. Dis. 2021, 68, 3089–3095. [Google Scholar] [CrossRef] [PubMed]
- Ullmann, L.S.; Langoni, H. Interactions between environment, wild animals and human leptospirosis. J. Venom. Anim. Toxins Incl. Trop. Dis. 2011, 17, 119–129. [Google Scholar] [CrossRef] [Green Version]
- Dietrich, M.; Wilkinson, D.A.; Benlali, A.; Lagadec, E.; Ramasindrazana, B.; Dellagi, K.; Tortosa, P. Leptospira and paramyxovirus infection dynamics in a bat maternity enlightens pathogen maintenance in wildlife. Environ. Microbiol. 2015, 17, 4280–4289. [Google Scholar] [CrossRef] [PubMed]
- Gili, F.; Newson, S.E.; Gillings, S.; Chamberlain, D.E.; Border, J.A. Bats in urbanising landscapes: Habitat selection and recommendations for a sustainable future. Biol. Conserv. 2020, 241, 108343. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Target | Gene | Primer Name | Primer Sequence 5′–3′ |
|---|---|---|---|
| Mammal | cytB | CytB Uni-F CytB Uni-R | TCATCMTGATGAAAYTTYGG ACTGGYTGDCCBCCRATTCA |
| Leptospira spp. | 16S rRNA | Lep1 Lep2 | GGCGGCGCGTCTTAAACATG TTCCCCCCATTGAGCAAGATT |
| Pathogenic Leptospira spp. | LipL32 | LipL32-270-F LipL32-692-R | CGCTGAAATGGGAGTTCGTATGATT CCAACAGATGCAACGAAAGATCCTTT |
| Family | Bat Species | Feeding Habits | Samples (n) | Leptospira spp. Detection | Frequency 2 (%) | |
|---|---|---|---|---|---|---|
| 16S rRNA 1 | LipL32 1 | |||||
| Phyllostomidae | Carollia perspicillata | Frugivorous | 30 | 11 | 0 | 36.7 |
| Mormoopidae | Mormoops megalophylla | Insectivorous | 30 | 17 | 14 | 56.7 |
| Natalidae | Natalus tumidirostris | Insectivorous | 25 | 16 | 14 | 64.0 |
| Total | 85 | 44 | 28 | 51.8 | ||
| Bats Samples(Code) | Bats Species | Leptospira Species 1 | Sequence Reference (GenBank Number) | Identity 2 (%) | Frequency of Infection (%) |
|---|---|---|---|---|---|
| MT100 | Natalus tumidirostris | Leptospira noguchii | U12671 | 85.56–99.00 | 9.09 |
| MT77 | |||||
| MT84 | |||||
| MT134 | |||||
| MT122 | Carollia perspicillata | Leptospira alexanderi | AY631880 MK726124 | 95.4–96.7 | 6.81 |
| MT83 | Natalus tumidirostris | ||||
| MT81 | |||||
| SCH3 | Natalus tumidirostris | Leptospira borgpetersenii | AM50569 MN047235 | 89.0–99.3 | 9.09 |
| MT71 MT72 SCH1 | |||||
| MT109 | Mormoops megalophylla | Leptospira mayottensis | MK719979 | 95.6 | 2.27 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva-Ramos, C.R.; Chala-Quintero, S.M.; Faccini-Martínez, Á.A.; Hidalgo, M.; Pulido-Villamarín, A.d.P.; Pérez-Torres, J.; Cuervo, C. Pathogenic Leptospira Species in Bats: Molecular Detection in a Colombian Cave. Trop. Med. Infect. Dis. 2022, 7, 84. https://doi.org/10.3390/tropicalmed7060084
Silva-Ramos CR, Chala-Quintero SM, Faccini-Martínez ÁA, Hidalgo M, Pulido-Villamarín AdP, Pérez-Torres J, Cuervo C. Pathogenic Leptospira Species in Bats: Molecular Detection in a Colombian Cave. Tropical Medicine and Infectious Disease. 2022; 7(6):84. https://doi.org/10.3390/tropicalmed7060084
Chicago/Turabian StyleSilva-Ramos, Carlos Ramiro, Sandra M. Chala-Quintero, Álvaro A. Faccini-Martínez, Marylin Hidalgo, Adriana del Pilar Pulido-Villamarín, Jairo Pérez-Torres, and Claudia Cuervo. 2022. "Pathogenic Leptospira Species in Bats: Molecular Detection in a Colombian Cave" Tropical Medicine and Infectious Disease 7, no. 6: 84. https://doi.org/10.3390/tropicalmed7060084