
The “Big Six”: Hidden Emerging Foodborne Bacterial Pathogens

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Characteristics of Diseases Caused by Non-O157 STEC Infections
3. Major Virulence Factors
3.1. Shiga toxin (Stx)
3.2. Enterocyte Effacement (LEE) Pathogenicity Island
3.3. Plasmid-Encoded Enterohemolysin
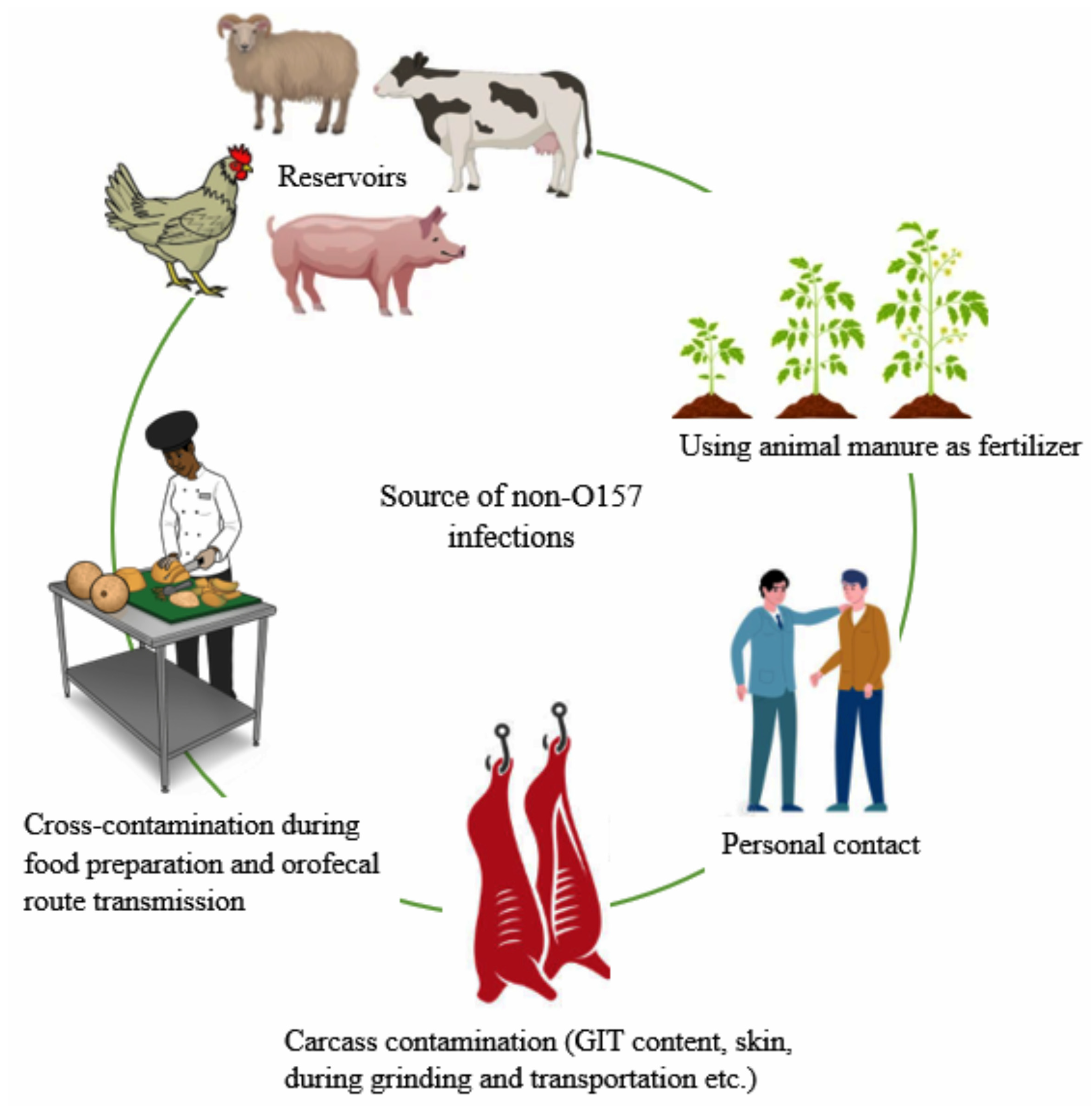
4. Reservoir, Source, and Transmission of Non-O157 E. coli
5. Six Serogroups of Non-O157 Shiga toxin-producing E. coli
5.1. E. coli Serogroup O26
5.2. E. coli Serogroup O45
5.3. E. coli Serogroup O103
5.4. E. coli Serogroup O111
5.5. E. coli Serogroup O121
5.6. E. coli Serogroup O145
6. Outbreaks Caused by Non-O157:H7 Infections
7. Diagnostic Approaches
7.1. Cultural Methods
7.2. Immunological Methods
7.3. Molecular Methods
7.4. Flow Cytometry
7.5. Metagenomics
8. Prevention and Control of Non-O157:H7 E. coli Infections
9. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, H.; Liu, Y.; Yang, L.; Wu, X.; Wu, Y.; Shao, B. Prevalence of Escherichia coli and Antibiotic Resistance in Animal-Derived Food Samples—Six Districts, Beijing, China, 2020. China CDC Wkly. 2021, 3, 999. [Google Scholar] [CrossRef] [PubMed]
- Sarowska, J.; Futoma-Koloch, B.; Jama-Kmiecik, A.; Frej-Madrzak, M.; Ksiazczyk, M.; Bugla-PLoSkonska, G.; Choroszy-Krol, I. Virulence factors, prevalence and potential transmission of extraintestinal pathogenic Escherichia coli isolated from different sources: Recent reports. Gut Pathog. 2019, 11, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teng, L.-J.; Hsueh, P.-R.; Liaw, S.-J.; Ho, S.-W.; Tsai, J.-C. Genetic detection of diarrheagenic Escherichia coli isolated from children with sporadic diarrhea. J. Microbiol. Immunol. Infect. 2004, 37, 327–334. [Google Scholar] [PubMed]
- Nguyen, T.V.; Le Van, P.; Le Huy, C.; Gia, K.N.; Weintraub, A. Detection and characterization of diarrheagenic Escherichia coli from young children in Hanoi, Vietnam. J. Clin. Microbiol. 2005, 43, 755–760. [Google Scholar] [CrossRef] [Green Version]
- Campos, L.C.; Franzolin, M.R.; Trabulsi, L.R. Diarrheagenic Escherichia coli categories among the traditional enteropathogenic E. coli O serogroups: A review. Mem. Do Inst. Oswaldo Cruz 2004, 99, 545–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karch, H.; Meyer, T.; Rüssmann, H.; Heesemann, J. Frequent loss of Shiga-like toxin genes in clinical isolates of Escherichia coli upon subcultivation. Infect. Immun. 1992, 60, 3464–3467. [Google Scholar] [CrossRef] [Green Version]
- Kaper, J.B.; Nataro, J.P.; Mobley, H.L. Pathogenic Escherichia coli. Nat. Rev. Microbiol. 2004, 2, 123–140. [Google Scholar] [CrossRef] [PubMed]
- Santos, F.F.; Yamamoto, D.; Abe, C.M.; Bryant, J.A.; Hernandes, R.T.; Kitamura, F.C.; Castro, F.S.; Valiatti, T.B.; Piazza, R.M.; Elias, W.P. The Type III Secretion System (T3SS)-translocon of atypical enteropathogenic Escherichia coli (aEPEC) can mediate adherence. Front. Microbiol. 2019, 10, 1527. [Google Scholar] [CrossRef] [Green Version]
- Trabulsi, L.R.; Keller, R.; Gomes, T.A.T. Typical and Atypical Enteropathogenic Escherichia coli. Emerg. Infect. Dis. 2002, 8, 508. [Google Scholar] [CrossRef]
- CDC. E. coli—General Information- What are Shiga Toxin-Producing E. coli (STEC). Available online: https://www.cdc.gov/ecoli/general/index.html (accessed on 21 August 2017).
- WHO. Zoonotic Non-0157 Shiga Toxin-Producing Escherichia coli (STEC): Report of a WHO Scientific Working Group Meeting, Berlin, Germany 23–26 June 1998; World Health Organization: Geneva, Switzerland.
- CDC. Escherichia coli (E. coli). Available online: https://www.cdc.gov/ecoli/pdfs/cdc-e.-coli-factsheet.pdf (accessed on 31 May 2017).
- Selim, S.A.; Ahmed, S.F.; Aziz, M.H.A.; Zakaria, A.M.; Klena, J.D.; Pangallo, D. Prevalence and characterization of Shiga-toxin O157: H7 and non-O157: H7 enterohemorrhagic Escherichia coli isolated from different sources. Biotechnol. Biotechnol. Equip. 2013, 27, 3834–3842. [Google Scholar] [CrossRef]
- Sahar, M.E.-A.; Salwa, F.A.; Samy, A.S.; Mohamed, H.A.A.; Amira, M.Z.; John, D.K. Prevalence and characterization of Shiga toxin O157 and non-O157 enterohemorrhagic Escherichia coli isolated from different sources in Ismailia, Egypt. Afr. J. Microbiol. Res. 2013, 7, 2637–2645. [Google Scholar] [CrossRef]
- Lupindu, A.M.; Olsen, J.E.; Ngowi, H.A.; Msoffe, P.L.; Mtambo, M.M.; Scheutz, F.; Dalsgaard, A. Occurrence and characterization of Shiga toxin-producing Escherichia coli O157:H7 and other non-sorbitol-fermenting E. coli in cattle and humans in urban areas of Morogoro, Tanzania. Vector Borne Zoonotic Dis. 2014, 14, 503–510. [Google Scholar] [CrossRef]
- Karama, M.; Cenci-Goga, B.T.; Malahlela, M.; Smith, A.M.; Keddy, K.H.; El-Ashram, S.; Kabiru, L.M.; Kalake, A. Virulence characteristics and antimicrobial resistance profiles of shiga toxin-producing Escherichia coli isolates from humans in South Africa: 2006–2013. Toxins 2019, 11, 424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vally, H.; Hall, G.; Dyda, A.; Raupach, J.; Knope, K.; Combs, B.; Desmarchelier, P. Epidemiology of Shiga toxin producing Escherichia coli in Australia, 2000–2010. BMC Public Health 2012, 12, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brusa, V.; Restovich, V.; Galli, L.; Teitelbaum, D.; Signorini, M.; Brasesco, H.; Londero, A.; García, D.; Padola, N.L.; Superno, V. Isolation and characterization of non-O157 Shiga toxin-producing Escherichia coli from beef carcasses, cuts and trimmings of abattoirs in Argentina. PLoS ONE 2017, 12, e0183248. [Google Scholar] [CrossRef]
- Authority, E.F.S.; European Centre for Disease Prevention and Control. The European Union one health 2020 zoonoses report. EFSA J. 2021, 19, e06971. [Google Scholar]
- Carroll, K.J.; Harvey-Vince, L.; Jenkins, C.; Mohan, K.; Balasegaram, S. The epidemiology of Shiga toxin-producing Escherichia coli infections in the South East of England: November 2013–March 2017 and significance for clinical and public health. J. Med. Microbiol. 2019, 68, 930–939. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, S.J.; Adak, G.K.; Gilham, C. Contact with farming environment as a major risk factor for Shiga toxin (Vero cytotoxin)-producing Escherichia coli O157 infection in humans. Emerg. Infect. Dis. 2001, 7, 1049. [Google Scholar] [CrossRef]
- Luna, R.; Mody, R.; Griffin, P. Non-O157 Shiga toxin-producing E. coli (STEC) outbreaks, United States. Dep. Health Hum. Serv. CfDCaPC Ed. Cent. Dis. Control Prev. Atlanta 2010. Available online: http://blogs.cdc.gov/publichealthmatters/files/2010/05/nono157stec_obs_052110 (accessed on 15 November 2011).
- CDC. E. coli Outbreak Investigations. Available online: https://www.cdc.gov/ecoli/outbreaks.html (accessed on 30 December 2021).
- Pihkala, N.; Bauer, N.; Eblen, D.; Evans, P.; Johnson, R.; Webb, J.; Williams, C. Risk profile for pathogenic non-O157 Shiga toxin-producing Escherichia coli (non-O157 STEC). US Dep. Agric. Food Saf. Insp. Serv. Public Health Sci. Policy Program Dev. 2012, 1–64. [Google Scholar]
- Yang, X.; Sun, H.; Fan, R.; Fu, S.; Zhang, J.; Matussek, A.; Xiong, Y.; Bai, X. Genetic diversity of the intimin gene (eae) in non-O157 Shiga toxin-producing Escherichia coli strains in China. Sci. Rep. 2020, 10, 3275. [Google Scholar] [PubMed] [Green Version]
- Fan, R.; Shao, K.; Yang, X.; Bai, X.; Fu, S.; Sun, H.; Xu, Y.; Wang, H.; Li, Q.; Hu, B. High prevalence of non-O157 Shiga toxin-producing Escherichia coli in beef cattle detected by combining four selective agars. BMC Microbiol. 2019, 19, 213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conrad, C.C.; Stanford, K.; McAllister, T.A.; Thomas, J.; Reuter, T. Further development of sample preparation and detection methods for O157 and the top 6 non-O157 STEC serogroups in cattle feces. J. Microbiol. Methods 2014, 105, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Valilis, E.; Ramsey, A.; Sidiq, S.; DuPont, H.L. Non-O157 Shiga toxin-producing Escherichia coli—A poorly appreciated enteric pathogen: Systematic review. Int. J. Infect. Dis. 2018, 76, 82–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vishram, B.; Jenkins, C.; Greig, D.R.; Godbole, G.; Carroll, K.; Balasegaram, S.; Byrne, L. The emerging importance of Shiga toxin-producing Escherichia coli other than serogroup O157 in England. J. Med. Microbiol. 2021, 70, 001375. [Google Scholar] [CrossRef] [PubMed]
- Ethelberg, S.; Smith, B.; Torpdahl, M.; Lisby, M.; Boel, J.; Jensen, T.; Nielsen, E.M.; Mølbak, K. Outbreak of non-O157 Shiga toxin-producing Escherichia coli infection from consumption of beef sausage. Clin. Infect. Dis. 2009, 48, e78–e81. [Google Scholar] [CrossRef] [Green Version]
- Gyles, C. Shiga toxin-producing Escherichia coli: An overview. J. Anim. Sci. 2007, 85, E45–E62. [Google Scholar] [CrossRef]
- Nataro, J.P.; Kaper, J.B. Diarrheagenic Escherichia coli. Clin. Microbiol. Rev. 1998, 11, 142–201. [Google Scholar] [CrossRef] [Green Version]
- Banatvala, N.; Griffin, P.M.; Greene, K.D.; Barrett, T.J.; Bibb, W.F.; Green, J.H.; Wells, J.G. The United States national prospective hemolytic uremic syndrome study: Microbiologic, serologic, clinical, and epidemiologic findings. J. Infect. Dis. 2001, 183, 1063–1070. [Google Scholar] [CrossRef] [Green Version]
- Griffin, P. CDC perspective on non-O157 Shiga toxinproducing E. coli (STEC) in the United States. In Proceedings of the Public Health Significance of Non-O157 Shiga Toxin-Producing Escherichia coli (STEC), Public Meeting, Arlington, VA, USA, 17 October 2007. [Google Scholar]
- Joseph, A.; Cointe, A.; Mariani Kurkdjian, P.; Rafat, C.; Hertig, A. Shiga toxin-associated hemolytic uremic syndrome: A narrative review. Toxins 2020, 12, 67. [Google Scholar]
- Glassman, H.; Ferrato, C.; Chui, L. Epidemiology of Non-O157 Shiga Toxin-Producing Escherichia coli in the Province of Alberta, Canada, from 2018 to 2021. Microorganisms 2022, 10, 814. [Google Scholar] [CrossRef] [PubMed]
- Konowalchuk, J.; Speirs, J.; Stavric, S. Vero response to a cytotoxin of Escherichia coli. Infect. Immun. 1977, 18, 775–779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Łoś, J.M.; Łoś, M.; Węgrzyn, G. Bacteriophages carrying Shiga toxin genes: Genomic variations, detection and potential treatment of pathogenic bacteria. Future Microbiol. 2011, 6, 909–924. [Google Scholar] [CrossRef] [PubMed]
- Macori, G.; McCarthy, S.C.; Burgess, C.M.; Fanning, S.; Duffy, G. Investigation of the causes of Shigatoxigenic Escherichia coli PCR positive and culture negative samples. Microorganisms 2020, 8, 587. [Google Scholar] [CrossRef]
- Melton-Celsa, A.R. Shiga toxin (Stx) classification, structure, and function. Microbiol. Spectr. 2014, 2, 4–6. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Tian, S.; Thaker, H.; Dong, M. Shiga toxins: An update on host factors and biomedical applications. Toxins 2021, 13, 222. [Google Scholar] [CrossRef]
- O’Brien, A.D.; LaVeck, G.D.; Thompson, M.R.; Formal, S.B. Production of Shigella dysenteriae type 1-like cytotoxin by Escherichia coli. J. Infect. Dis. 1982, 146, 763–769. [Google Scholar] [CrossRef] [Green Version]
- Bai, X.; Scheutz, F.; Dahlgren, H.M.; Hedenström, I.; Jernberg, C. Characterization of Clinical Escherichia coli Strains Producing a Novel Shiga Toxin 2 Subtype in Sweden and Denmark. Microorganisms 2021, 9, 2374. [Google Scholar] [CrossRef]
- Fuller, C.A.; Pellino, C.A.; Flagler, M.J.; Strasser, J.E.; Weiss, A.A. Shiga toxin subtypes display dramatic differences in potency. Infect. Immun. 2011, 79, 1329–1337. [Google Scholar] [CrossRef] [Green Version]
- Petro, C.D.; Trojnar, E.; Sinclair, J.; Liu, Z.-M.; Smith, M.; O’Brien, A.D.; Melton-Celsa, A. Shiga toxin type 1a (Stx1a) reduces the toxicity of the more potent Stx2a in vivo and in vitro. Infect. Immun. 2019, 87, e00787-18. [Google Scholar] [CrossRef] [Green Version]
- Furniss, R.C.D.; Clements, A. Regulation of the locus of enterocyte effacement in attaching and effacing pathogens. J. Bacteriol. 2018, 200, e00336-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montero, D.A.; Canto, F.D.; Velasco, J.; Colello, R.; Padola, N.L.; Salazar, J.C.; Martin, C.S.; Oñate, A.; Blanco, J.; Rasko, D.A. Cumulative acquisition of pathogenicity islands has shaped virulence potential and contributed to the emergence of LEE-negative Shiga toxin-producing Escherichia coli strains. Emerg. Microbes Infect. 2019, 8, 486–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lorenz, S.C.; Son, I.; Maounounen-Laasri, A.; Lin, A.; Fischer, M.; Kase, J.A. Prevalence of hemolysin genes and comparison of ehxA subtype patterns in Shiga toxin-producing Escherichia coli (STEC) and non-STEC strains from clinical, food, and animal sources. Appl. Environ. Microbiol. 2013, 79, 6301–6311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beutin, L.; Montenegro, M.A.; Orskov, I.; Orskov, F.; Prada, J.; Zimmermann, S.; Stephan, R. Close association of verotoxin (Shiga-like toxin) production with enterohemolysin production in strains of Escherichia coli. J. Clin. Microbiol. 1989, 27, 2559–2564. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, H.; Maier, E.; Karch, H.; Benz, R. Pore-Forming Properties of the Plasmid—Encoded Hemolysin of Enterohemorrhagic Escherichia coli 0157: H7. Eur. J. Biochem. 1996, 241, 594–601. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.Y.; Yoon, J.W.; Hovde, C.J. A brief overview of Escherichia coli O157: H7 and its plasmid O157. J. Microbiol. Biotechnol. 2010, 20, 5. [Google Scholar] [CrossRef] [Green Version]
- Gioia-Di Chiacchio, R.; Cunha, M.; Sturn, R.; Moreno, L.; Moreno, A.M.; Pereira, C.; Martins, F.; Franzolin, M.; Piazza, R.; Knöbl, T. Shiga toxin-producing Escherichia coli (STEC): Zoonotic risks associated with psittacine pet birds in home environments. Vet. Microbiol. 2016, 184, 27–30. [Google Scholar] [CrossRef]
- OzFoodNet. OzFoodNet: Enhancing foodborne disease surveillance across Australia: Quarterly report, 1 July to 30 September 2003. Commun. Dis. Intell. Q. Rep. 2003, 27, 504–507. [Google Scholar]
- Koohmaraie, M.; Arthur, T.; Bosilevac, J.; Guerini, M.; Shackelford, S.; Wheeler, T. Post-harvest interventions to reduce/eliminate pathogens in beef. Meat Sci. 2005, 71, 79–91. [Google Scholar] [CrossRef] [Green Version]
- Boerlin, P.; McEwen, S.A.; Boerlin-Petzold, F.; Wilson, J.B.; Johnson, R.P.; Gyles, C.L. Associations between virulence factors of Shiga toxin-producing Escherichia coli and disease in humans. J. Clin. Microbiol. 1999, 37, 497–503. [Google Scholar] [CrossRef] [Green Version]
- Persad, A.K.; Lejeune, J.T. Animal reservoirs of Shiga toxin—Producing Escherichia coli. In Enterohemorrhagic Escherichia coli and Other Shiga Toxin-Producing E. coli; ASM Press: Washington, DC, USA, 2015; pp. 211–230. [Google Scholar]
- Shahzad, A.; Ullah, F.; Irshad, H.; Ahmed, S.; Shakeela, Q.; Mian, A.H. Molecular detection of Shiga toxin-producing Escherichia coli (STEC) O157 in sheep, goats, cows and buffaloes. Mol. Biol. Rep. 2021, 48, 6113–6121. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, S.C.; Macori, G.; Duggan, G.; Burgess, C.M.; Fanning, S.; Duffy, G. Prevalence and Whole-Genome Sequence-Based Analysis of Shiga Toxin-Producing Escherichia coli Isolates from the Recto-Anal Junction of Slaughter-Age Irish Sheep. Appl. Environ. Microbiol. 2021, 87, e01384-21. [Google Scholar] [CrossRef] [PubMed]
- Karmali, M.A. Emerging public health challenges of Shiga toxin–producing Escherichia coli related to changes in the pathogen, the population, and the environment. Clin. Infect. Dis. 2017, 64, 371–376. [Google Scholar]
- Paquette, S.-J.; Stanford, K.; Thomas, J.; Reuter, T. Quantitative surveillance of shiga toxins 1 and 2, Escherichia coli O178 and O157 in feces of western-Canadian slaughter cattle enumerated by droplet digital PCR with a focus on seasonality and slaughterhouse location. PLoS ONE 2018, 13, e0195880. [Google Scholar] [CrossRef] [Green Version]
- McCabe, E.; Burgess, C.M.; Lawal, D.; Whyte, P.; Duffy, G. An investigation of shedding and super—Shedding of Shiga toxigenic Escherichia coli O157 and E. coli O26 in cattle presented for slaughter in the Republic of Ireland. Zoonoses Public Health 2019, 66, 83–91. [Google Scholar] [CrossRef] [Green Version]
- McCarthy, S.C.; Burgess, C.M.; Fanning, S.; Duffy, G. An overview of Shiga-toxin producing Escherichia coli carriage and prevalence in the ovine meat production chain. Foodborne Pathog. Dis. 2021, 18, 147–168. [Google Scholar] [CrossRef]
- Mathusa, E.C.; Chen, Y.; Enache, E.; Hontz, L. Non-O157 Shiga toxin–producing Escherichia coli in foods. J. Food Prot. 2010, 73, 1721–1736. [Google Scholar] [CrossRef]
- Foodonline. USDA Takes New Steps To Fight E. coli, Protect The Food Supply. Available online: https://www.foodonline.com/doc/usda-takes-new-steps-to-fight-e-coli-0001 (accessed on 14 September 2011).
- Liao, Y.-T.; Miller, M.F.; Loneragan, G.H.; Brooks, J.C.; Echeverry, A.; Brashears, M.M. Non-O157 Shiga toxin–producing Escherichia coli in US retail ground beef. J. Food Prot. 2014, 77, 1188–1192. [Google Scholar] [CrossRef]
- Hoshina, K.; Itagaki, A.; Seki, R.; Yamamoto, K.; Masuda, S.-i.; Muku, T.; Okada, N. Enterohemorrhagic Escherichia coli O26 outbreak caused by contaminated natural water supplied by facility owned by local community. Jpn. J. Infect. Dis. 2001, 54, 247–248. [Google Scholar]
- Peron, E.; Zaharia, A.; Zota, L.C.; Severi, E.; Mårdh, O.; Usein, C.; Bălgrădean, M.; Espinosa, L.; Jansa, J.; Scavia, G. Early findings in outbreak of haemolytic uraemic syndrome among young children caused by Shiga toxin-producing Escherichia coli, Romania, January to February 2016. Eurosurveillance 2016, 21, 30170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaros, P.; Cookson, A.L.; Campbell, D.M.; Besser, T.E.; Shringi, S.; Mackereth, G.F.; Lim, E.; Lopez, L.; Dufour, M.; Marshall, J.C. A prospective case–control and molecular epidemiological study of human cases of Shiga toxin-producing Escherichia coli in New Zealand. BMC Infect. Dis. 2013, 13, 450. [Google Scholar] [CrossRef] [Green Version]
- Alegbeleye, O.O.; Sant’Ana, A.S. Manure-borne pathogens as an important source of water contamination: An update on the dynamics of pathogen survival/transport as well as practical risk mitigation strategies. Int. J. Hyg. Environ. Health 2020, 227, 113524. [Google Scholar] [CrossRef] [PubMed]
- CDC. Control, and Prevention, Importance of culture confirmation of Shiga toxin-producing Escherichia coli infection as illustrated by outbreaks of gastroenteritis--New York and North Carolina, 2005. MMWR. Morb. Mortal. Wkly. Rep. 2006, 553, 1042–1045. [Google Scholar]
- Carbonari, C.C.; Miliwebsky, E.S.; Zolezzi, G.; Deza, N.L.; Fittipaldi, N.; Manfredi, E.; Baschkier, A.; D’Astek, B.A.; Melano, R.G.; Schesi, C. The Importance of Shiga Toxin-Producing Escherichia coli O145: NM [H28]/H28 Infections in Argentina, 1998–2020. Microorganisms 2022, 10, 582. [Google Scholar] [CrossRef] [PubMed]
- Bielaszewska, M.; Zhang, W.; Tarr, P.I.; Sonntag, A.-K.; Karch, H. Molecular profiling and phenotype analysis of Escherichia coli O26: H11 and O26: NM: Secular and geographic consistency of enterohemorrhagic and enteropathogenic isolates. J. Clin. Microbiol. 2005, 43, 4225–4228. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.-L.; Bielaszewska, M.; Bockemühl, J.; Schmidt, H.; Scheutz, F.; Karch, H. Molecular analysis of H antigens reveals that human diarrheagenic Escherichia coli O26 strains that carry the eae gene belong to the H11 clonal complex. J. Clin. Microbiol. 2000, 38, 2989–2993. [Google Scholar] [CrossRef] [Green Version]
- Jenkins, C.; Evans, J.; Chart, H.; Willshaw, G.; Frankel, G. Escherichia coli serogroup O26–a new look at an old adversary. J. Appl. Microbiol. 2008, 104, 14–25. [Google Scholar] [CrossRef]
- Anjum, M.F.; Lucchini, S.; Thompson, A.; Hinton, J.C.; Woodward, M.J. Comparative genomic indexing reveals the phylogenomics of Escherichia coli pathogens. Infect. Immun. 2003, 71, 4674–4683. [Google Scholar] [CrossRef]
- Cid, D.; Ruiz-Santa-Quiteria, J.; Marın, I.; Sanz, R.; Orden, J.; Amils, R.; De La Fuente, R. Association between intimin (eae) and EspB gene subtypes in attaching and effacing Escherichia coli strains isolated from diarrhoeic lambs and goat kids. Microbiology 2001, 147, 2341–2353. [Google Scholar] [CrossRef] [Green Version]
- Fröhlicher, E.; Krause, G.; Zweifel, C.; Beutin, L.; Stephan, R. Characterization of attaching and effacing Escherichia coli (AEEC) isolated from pigs and sheep. BMC Microbiol. 2008, 8, 144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krause, G.; Zimmermann, S.; Beutin, L. Investigation of domestic animals and pets as a reservoir for intimin-(eae) gene positive Escherichia coli types. Vet. Microbiol. 2005, 106, 87–95. [Google Scholar] [CrossRef]
- Pearce, M.; Evans, J.; McKendrick, I.; Smith, A.; Knight, H.; Mellor, D.; Woolhouse, M.; Gunn, G.; Low, J. Prevalence and virulence factors of Escherichia coli serogroups O26, O103, O111, and O145 shed by cattle in Scotland. Appl. Environ. Microbiol. 2006, 72, 653–659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanco, M.; Schumacher, S.; Tasara, T.; Zweifel, C.; Blanco, J.E.; Dahbi, G.; Blanco, J.; Stephan, R. Serotypes, intimin variants and other virulence factors of eae positive Escherichia coli strains isolated from healthy cattle in Switzerland. Identification of a new intimin variant gene (eae-η2). BMC Microbiol. 2005, 5, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, J.; Knight, H.; McKendrick, I.J.; Stevenson, H.; Barbudo, A.V.; Gunn, G.J.; Low, J.C. Prevalence of Escherichia coli O157: H7 and serogroups O26, O103, O111 and O145 in sheep presented for slaughter in Scotland. J. Med. Microbiol. 2011, 60, 653–660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoyle, D.V.; Keith, M.; Williamson, H.; Macleod, K.; Mathie, H.; Handel, I.; Currie, C.; Holmes, A.; Allison, L.; McLean, R. Prevalence and epidemiology of non-O157 Escherichia coli serogroups O26, O103, O111, and O145 and Shiga toxin gene carriage in Scottish cattle, 2014–2015. Appl. Environ. Microbiol. 2021, 87, e03142-03120. [Google Scholar] [CrossRef] [PubMed]
- Hermos, C.R.; Janineh, M.; Han, L.L.; McAdam, A.J. Shiga toxin-producing Escherichia coli in children: Diagnosis and clinical manifestations of O157: H7 and non-O157: H7 infection. J. Clin. Microbiol. 2011, 49, 955–959. [Google Scholar] [CrossRef] [Green Version]
- Pollock, K.G.; Bhojani, S.; Beattie, T.J.; Allison, L.; Hanson, M.; Locking, M.E.; Cowden, J.M. Highly virulent Escherichia coli O26, Scotland. Emerg. Infect. Dis. 2011, 17, 1777. [Google Scholar] [CrossRef]
- Bruyand, M.; Mariani-Kurkdjian, P.; Le Hello, S.; King, L.-A.; Van Cauteren, D.; Lefevre, S.; Gouali, M.; Jourdan-da Silva, N.; Mailles, A.; Donguy, M.-P. Paediatric haemolytic uraemic syndrome related to Shiga toxin-producing Escherichia coli, an overview of 10 years of surveillance in France, 2007 to 2016. Eurosurveillance 2019, 24, 1800068. [Google Scholar] [CrossRef]
- Jones, G.; Lefèvre, S.; Donguy, M.-P.; Nisavanh, A.; Terpant, G.; Fougère, E.; Vaissière, E.; Guinard, A.; Mailles, A.; de Valk, H. Outbreak of Shiga toxin-producing Escherichia coli (STEC) O26 paediatric haemolytic uraemic syndrome (HUS) cases associated with the consumption of soft raw cow’s milk cheeses, France, March to May 2019. Eurosurveillance 2019, 24, 1900305. [Google Scholar] [CrossRef] [Green Version]
- Severi, E.; Vial, F.; Peron, E.; Mardh, O.; Niskanen, T.; Takkinen, J. Community-wide outbreaks of haemolytic uraemic syndrome associated with Shiga toxin-producing Escherichia coli O26 in Italy and Romania: A new challenge for the European Union. Eurosurveillance 2016, 21, 30420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loconsole, D.; Giordano, M.; Centrone, F.; Accogli, M.; Casulli, D.; De Robertis, A.L.; Morea, A.; Quarto, M.; Parisi, A.; Scavia, G. Epidemiology of Shiga toxin-producing Escherichia coli infections in Southern Italy after implementation of symptom-based surveillance of bloody diarrhea in the pediatric population. Int. J. Environ. Res. Public Health 2020, 17, 5137. [Google Scholar] [CrossRef] [PubMed]
- Brooks, J.T.; Bergmire-Sweat, D.; Kennedy, M.; Hendricks, K.; Garcia, M.; Marengo, L.; Wells, J.; Ying, M.; Bibb, W.; Griffin, P.M. Outbreak of Shiga toxin—Producing Escherichia coli O111: H8 infections among attendees of a high school cheerleading camp. Clin. Infect. Dis. 2004, 38, 190–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaffzin, J.; Coronado, F.; Dumas, N.; Root, T.; Halse, T.; Schoonmaker-Bopp, D.; Lurie, M.; Nicholas, D.; Gerzonich, B.; Johnson, G. Public health approach to detection of non-O157 Shiga toxin-producing Escherichia coli: Summary of two outbreaks and laboratory procedures. Epidemiol. Infect. 2012, 140, 283–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luna-Gierke, R.; Griffin, P.; Gould, L.; Herman, K.; Bopp, C.; Strockbine, N.; Mody, R. Outbreaks of non-O157 Shiga toxin-producing Escherichia coli infection: USA. Epidemiol. Infect. 2014, 142, 2270–2280. [Google Scholar] [CrossRef] [PubMed]
- Cooper, K.K.; Mandrell, R.E.; Louie, J.W.; Korlach, J.; Clark, T.A.; Parker, C.T.; Huynh, S.; Chain, P.S.; Ahmed, S.; Carter, M.Q. Comparative genomics of enterohemorrhagic Escherichia coli O145: H28 demonstrates a common evolutionary lineage with Escherichia coli O157: H7. BMC Genom. 2014, 15, 17. [Google Scholar] [CrossRef] [Green Version]
- Michino, H.; Araki, K.; Minami, S.; Takaya, S.; Sakai, N.; Miyazaki, M.; Ono, A.; Yanagawa, H. Massive outbreak of Escherichia coli O157: H7 infection in schoolchildren in Sakai City, Japan, associated with consumption of white radish sprouts. Am. J. Epidemiol. 1999, 150, 787–796. [Google Scholar]
- Sekse, C.; Sunde, M.; Hopp, P.; Bruheim, T.; Cudjoe, K.S.; Kvitle, B.; Urdahl, A.M. Occurrence of potentially human-pathogenic Escherichia coli O103 in Norwegian sheep. Appl. Environ. Microbiol. 2013, 79, 7502–7509. [Google Scholar] [CrossRef] [Green Version]
- Spika, J.; Khakhria, R.; Michel, P.; Milley, D.; Wilson, J.; Waters, J. Shiga toxin-producing Escherichia coli infections in Canada. Escherichia coli O 1998, 157, 23–29. [Google Scholar]
- Sekse, C.; O’Sullivan, K.; Granum, P.E.; Rørvik, L.M.; Wasteson, Y.; Jørgensen, H.J. An outbreak of Escherichia coli O103: H25—Bacteriological investigations and genotyping of isolates from food. Int. J. Food Microbiol. 2009, 133, 259–264. [Google Scholar] [CrossRef]
- Martins, F.H.; Guth, B.E.; Piazza, R.M.; Blanco, J.; Pelayo, J.S. First description of a Shiga toxin-producing Escherichia coli O103: H2 strain isolated from sheep in Brazil. J. Infect. Dev. Ctries. 2014, 8, 126–128. [Google Scholar] [CrossRef] [PubMed]
- Miko, A.; Pries, K.; Haby, S.; Steege, K.; Albrecht, N.; Krause, G.; Beutin, L. Assessment of Shiga toxin-producing Escherichia coli isolates from wildlife meat as potential pathogens for humans. Appl. Environ. Microbiol. 2009, 75, 6462–6470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanayama, A.; Yahata, Y.; Arima, Y.; Takahashi, T.; Saitoh, T.; Kanou, K.; Kawabata, K.; Sunagawa, T.; Matsui, T.; Oishi, K. Enterohemorrhagic Escherichia coli outbreaks related to childcare facilities in Japan, 2010–2013. BMC Infect. Dis. 2015, 15, 539. [Google Scholar] [CrossRef] [PubMed]
- Mylius, M.; Dreesman, J.; Pulz, M.; Pallasch, G.; Beyrer, K.; Claußen, K.; Allerberger, F.; Fruth, A.; Lang, C.; Prager, R. Shiga toxin-producing Escherichia coli O103: H2 outbreak in Germany after school trip to Austria due to raw cow milk, 2017–The important role of international collaboration for outbreak investigations. Int. J. Med. Microbiol. 2018, 308, 539–544. [Google Scholar] [CrossRef]
- Jeon, B.-W.; Jeong, J.-M.; Won, G.-Y.; Park, H.; Eo, S.-K.; Kang, H.-Y.; Hur, J.; Lee, J.H. Prevalence and characteristics of Escherichia coli O26 and O111 from cattle in Korea. Int. J. Food Microbiol. 2006, 110, 123–126. [Google Scholar] [CrossRef]
- Kato, K.; Shimoura, R.; Nashimura, K.; Yoshifuzi, K.; Shiroshita, K.; Sakurai, N.; Kodama, H.; Kuramoto, S. Outbreak of enterohemorrhagic Escherichia coli O111 among high school participants in excursion to Korea. Jpn. J. Infect. Dis. 2005, 58, 332. [Google Scholar]
- Kauffmann, F.; Dupont, A. Escherichia Strains from Infantile Epidemic Gastro-Enterltis. Acta Pathol. et Microbiol. Scand. 1950, 27, 552–564. [Google Scholar] [CrossRef]
- Moyenuddin, M.; Wachsmuth, I.; Moseley, S.; Bopp, C.; Blake, P. Serotype, antimicrobial resistance, and adherence properties of Escherichia coli strains associated with outbreaks of diarrheal illness in children in the United States. J. Clin. Microbiol. 1989, 27, 2234–2239. [Google Scholar] [CrossRef] [Green Version]
- Regua, A.H.; Bravo, V.; Leal, M.d.C.; Lobo Leite, M. Epidemiological survey of the enteropathogenic Escherichia coli isolated from children with diarrhoea. J. Trop. Pediatr. 1990, 36, 176–179. [Google Scholar] [CrossRef]
- Viljanen, M.; Peltola, T.; Kuistila, M.; Huovinen, P.; Junnila, S.; Olkkonen, L.; Järvinen, H. Outbreak of diarrhoea due to Escherichia coli 0111: B4 in schoolchildren and adults: Association of Vi antigen-like reactivity. Lancet 1990, 336, 831–834. [Google Scholar] [CrossRef]
- Caprioli, A.; Luzzi, I.; Rosmini, F.; Resti, C.; Edefonti, A.; Perfumo, F.; Farina, C.; Goglio, A.; Gianviti, A.; Rizzoni, G. Communitywide outbreak of hemolytic-uremic syndrome associated with non-O157 verocytotoxin-producing Escherichia coli. J. Infect. Dis. 1994, 169, 208–211. [Google Scholar] [CrossRef] [PubMed]
- Paton, A.W.; Ratcliff, R.M.; Doyle, R.M.; Seymour-Murray, J.; Davos, D.; Lanser, J.A.; Paton, J.C. Molecular microbiological investigation of an outbreak of hemolytic-uremic syndrome caused by dry fermented sausage contaminated with Shiga-like toxin-producing Escherichia coli. J. Clin. Microbiol. 1996, 34, 1622–1627. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, T.A.; Barrett, N.L.; Hadler, J.L.; Salsbury, B.; Howard, R.T.; Dingman, D.W.; Brinkman, C.D.; Bibb, W.F.; Cartter, M.L. Hemolytic-uremic syndrome and Escherichia coli O121 at a lake in Connecticut, 1999. Pediatrics 2001, 108, e59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stock, K.J.; Scott, M.A.; Davis, S.F.; Pierson Iii, R.N.; Dummer, J.S. Hemorrhagic colitis due to a novel Escherichia coli serotype (O121: H19) in a transplant patient. Transpl. Int. 2001, 14, 44–47. [Google Scholar] [CrossRef] [PubMed]
- Cornu, G.; Proesmans, W.; Dediste, A.; Jacobs, F.; Van De Walle, J.; Mertens, A.; Ramet, J.; Lauwers, S. Hemolytic uremic syndrome in Belgium: Incidence and association with verocytotoxin-producing Escherichia coli infection. Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 1999, 5, 16–22. [Google Scholar] [CrossRef] [Green Version]
- Hiruta, N.; Himori, M.; Habutsu, M.; Okamura, N.; Ogawa, M.; Matsushita, S.; Kudoh, Y. Enteroinvasive Escherichia coli O121: H-isolated from travellers’ diarrhea. Kansenshogaku Zasshi. J. Jpn. Assoc. Infect. Dis. 1991, 65, 537–539. [Google Scholar] [CrossRef]
- Tarr, C.L.; Large, T.M.; Moeller, C.L.; Lacher, D.W.; Tarr, P.I.; Acheson, D.W.; Whittam, T.S. Molecular characterization of a serotype O121: H19 clone, a distinct Shiga toxin-producing clone of pathogenic Escherichia coli. Infect. Immun. 2002, 70, 6853–6859. [Google Scholar] [CrossRef] [Green Version]
- Rivero, M.A.; Passucci, J.A.; Rodriguez, E.M.; Parma, A.E. Role and clinical course of verotoxigenic Escherichia coli infections in childhood acute diarrhoea in Argentina. J. Med. Microbiol. 2010, 59, 345–352. [Google Scholar] [CrossRef] [Green Version]
- Beutin, L.; Zimmermann, S.; Gleier, K. Human infections with Shiga toxin-producing Escherichia coli other than serogroup O157 in Germany. Emerg. Infect. Dis. 1998, 4, 635. [Google Scholar] [CrossRef] [Green Version]
- De Schrijver, K.; Buvens, G.; Possé, B.; Van den Branden, D.; Oosterlynck, O.; De Zutter, L.; Eilers, K.; Piérard, D.; Dierick, K.; Van Damme-Lombaerts, R. Outbreak of verocytotoxin-producing E. coli O145 and O26 infections associated with the consumption of ice cream produced at a farm, Belgium, 2007. Eurosurveillance 2008, 13, 9–10. [Google Scholar] [CrossRef]
- Yoder, J.; Roberts, V.; Craun, G.F.; Hill, V.; Hicks, L.A.; Alexander, N.T.; Radke, V.; Calderon, R.L.; Hlavsa, M.C.; Beach, M.J. Surveillance for waterborne disease and outbreaks associated with drinking water and water not intended for drinking—United States, 2005–2006. Morb. Mortal. Wkly. Report. Surveill. Summ. 2008, 57, 39–62. [Google Scholar]
- Taylor, E.; Nguyen, T.; Machesky, K.; Koch, E.; Sotir, M.; Bohm, S.; Folster, J.; Bokanyi, R.; Kupper, A.; Bidol, S. Multistate outbreak of Escherichia coli O145 infections associated with romaine lettuce consumption, 2010. J. Food Prot. 2013, 76, 939–944. [Google Scholar] [CrossRef] [PubMed]
- Tenover, F.C.; Arbeit, R.D.; Goering, R.V.; Mickelsen, P.A.; Murray, B.E.; Persing, D.H.; Swaminathan, B. Interpreting chromosomal DNA restriction patterns produced by pulsed-field gel electrophoresis: Criteria for bacterial strain typing. J. Clin. Microbiol. 1995, 33, 2233–2239. [Google Scholar] [CrossRef] [PubMed]
- Bettelheim, K.A. The non-O157 shiga-toxigenic (verocytotoxigenic) Escherichia coli; under-rated pathogens. Crit. Rev. Microbiol. 2007, 33, 67–87. [Google Scholar] [CrossRef]
- Arthur, T.M.; Barkocy-Gallagher, G.A.; Rivera-Betancourt, M.; Koohmaraie, M. Prevalence and characterization of non-O157 Shiga toxin-producing Escherichia coli on carcasses in commercial beef cattle processing plants. Appl. Environ. Microbiol. 2002, 68, 4847–4852. [Google Scholar] [CrossRef] [Green Version]
- Rounds, J.M.; Rigdon, C.E.; Muhl, L.J.; Forstner, M.; Danzeisen, G.T.; Koziol, B.S.; Taylor, C.; Shaw, B.T.; Short, G.L.; Smith, K.E. Non-O157 Shiga toxin–producing Escherichia coli associated with venison. Emerg. Infect. Dis. 2012, 18, 279. [Google Scholar] [CrossRef]
- CDC. Outbreak of E. coli Infections Linked to Ground Beef. Available online: https://www.cdc.gov/ecoli/2019/o103-04-19/index.html (accessed on 26 February 2022).
- CDC. Multistate Outbreaks of Shiga Toxin-Producing Escherichia coli O26 Infections Linked to Chipotle Mexican Grill Restaurants (Final Update). Available online: https://www.cdc.gov/ecoli/2015/o26-11-15/index.html (accessed on 31 May 2017).
- Vogel, G. Egyptian fenugreek seeds blamed for deadly E. coli outbreak; European authorities issue recall. ScienceInsider. Uploaded 2011, 5. [Google Scholar]
- CDC. Multistate Outbreak of Human E. coli O145 Infections Linked to Shredded Romaine Lettuce from a Single Processing Facility (FINAL UPDATE). Available online: https://www.cdc.gov/ecoli/2010/shredded-romaine-5-21-10.html (accessed on 25 March 2019).
- Bradley, K.; Williams, J.; Burnsed, L.; Lytle, M.; McDermott, M.; Mody, R.; Bhattarai, A.; Mallonee, S.; Piercefield, E.; McDonald-Hamm, C. Epidemiology of a large restaurant-associated outbreak of Shiga toxin-producing Escherichia coli O111: NM. Epidemiol. Infect. 2012, 140, 1644–1654. [Google Scholar] [CrossRef] [Green Version]
- Diercke, M.; Kirchner, M.; Claussen, K.; Mayr, E.; Strotmann, I.; Frangenberg, J.; Schiffmann, A.; Bettge-Weller, G.; Arvand, M.; Uphoff, H. Transmission of shiga toxin-producing Escherichia coli O104: H4 at a family party possibly due to contamination by a food handler, Germany 2011. Epidemiol. Infect. 2014, 142, 99–106. [Google Scholar] [CrossRef]
- Ethelberg, S.; Smith, B.; Torpdahl, M.; Lisby, M.; Boel, J.; Jensen, T.; Mølbak, K. An outbreak of Verocytotoxin-producing Escherichia coli O26: H11 caused by beef sausage, Denmark 2007. Wkly. Releases 2007, 12, 3208. [Google Scholar] [CrossRef]
- CDC. Community outbreak of hemolytic uremic syndrome attributable to Escherichia coli O111: NM--South Australia 1995. MMWR Morb. Mortal. Wkly. Rep. 1995, 44, 550–551, 557. [Google Scholar]
- Banatvala, N.; Debeukelaer, M.M.; Griffin, P.M.; Barrett, T.J.; Greene, K.D.; Green, J.H.; Wells, J.G. Shiga-like toxin-producing Escherichia coli O111 and associated hemolytic-uremic syndrome: A family outbreak. Pediatr. Infect. Dis. J. 1996, 15, 1008–1011. [Google Scholar] [CrossRef] [PubMed]
- Remis, R.S.; MacDonald, K.L.; Riley, L.W.; Puhr, N.D.; Wells, J.G.; Davis, B.R.; Blake, P.A.; Cohen, M.L. Sporadic cases of hemorrhagic colitis associated with Escherichia coli 0157: H7. Ann. Intern. Med. 1984, 101, 624–626. [Google Scholar] [CrossRef] [PubMed]
- Ammon, A.; Petersen, L.R.; Karch, H. A Large Outbreak of Hemolytic Uremic Syndrome Caused by an Unusual Sorbitol-Fermenting Strain of Escherichia coli O157: H—. J. Infect. Dis. 1999, 179, 1274–1277. [Google Scholar] [CrossRef] [Green Version]
- Gunzer, F.; Böhm, H.; Rüssmann, H.; Bitzan, M.; Aleksic, S.; Karch, H. Molecular detection of sorbitol-fermenting Escherichia coli O157 in patients with hemolytic-uremic syndrome. J. Clin. Microbiol. 1992, 30, 1807–1810. [Google Scholar] [CrossRef]
- Bettelheim, K. Reliability of CHROMagar® O157 for the detection of enterohaemorrhagic Escherichia coli (EHEC) O157 but not EHEC belonging to other serogroups. J. Appl. Microbiol. 1998, 85, 425–428. [Google Scholar] [CrossRef]
- Yoshitomi, K.J.; Jinneman, K.C.; Zapata, R.; Weagant, S.D.; Fedio, W.M. Detection and Isolation of Low Levels of E. coli O157: H7 in Cilantro by Real—Time PCR, Immunomagnetic Separation, and Cultural Methods with and without an Acid Treatment. J. Food Sci. 2012, 77, M481–M489. [Google Scholar] [CrossRef]
- Ngwa, G.; Schop, R.; Weir, S.; León-Velarde, C.; Odumeru, J. Detection and enumeration of E. coli O157: H7 in water samples by culture and molecular methods. J. Microbiol. Methods 2013, 92, 164–172. [Google Scholar] [CrossRef]
- Zelyas, N.; Poon, A.; Patterson-Fortin, L.; Johnson, R.P.; Lee, W.; Chui, L. Assessment of commercial chromogenic solid media for the detection of non-O157 Shiga toxin-producing Escherichia coli (STEC). Diagn. Microbiol. Infect. Dis. 2016, 85, 302–308. [Google Scholar] [CrossRef]
- Hughes, A.C.; Patfield, S.; Rasooly, R.; He, X. Validation of a Cell-Based Assay for Detection of Active Shiga Toxins Produced by Escherichia coli in Water. Int. J. Environ. Res. Public Health 2020, 17, 7901. [Google Scholar] [CrossRef]
- Teel, L.D.; Daly, J.A.; Jerris, R.C.; Maul, D.; Svanas, G.; O’Brien, A.D.; Park, C.H. Rapid detection of Shiga toxin-producing Escherichia coli by optical immunoassay. J. Clin. Microbiol. 2007, 45, 3377–3380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chui, L.; Lee, M.-C.; Allen, R.; Bryks, A.; Haines, L.; Boras, V. Comparison between ImmunoCard STAT!® and real-time PCR as screening tools for both O157: H7 and non-O157 Shiga toxin-producing Escherichia coli in Southern Alberta, Canada. Diagn. Microbiol. Infect. Dis. 2013, 77, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Chui, L.; Patterson-Fortin, L.; Kuo, J.; Li, V.; Boras, V. Evaluation of enzyme immunoassays and real-time PCR for detecting Shiga toxin-producing Escherichia coli in Southern Alberta, Canada. J. Clin. Microbiol. 2015, 53, 1019–1023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bélanger, S.D.; Boissinot, M.; Ménard, C.; Picard, F.J.; Bergeron, M.G. Rapid detection of Shiga toxin-producing bacteria in feces by multiplex PCR with molecular beacons on the smart cycler. J. Clin. Microbiol. 2002, 40, 1436–1440. [Google Scholar] [CrossRef] [Green Version]
- Fratamico, P.M.; Bagi, L.K. Detection of Shiga toxin-producing Escherichia coli in ground beef using the GeneDisc real-time PCR system. Front. Cell. Infect. Microbiol. 2012, 2, 152. [Google Scholar] [CrossRef]
- Macori, G.; McCarthy, S.C.; Burgess, C.M.; Fanning, S.; Duffy, G. A quantitative real time PCR assay to detect and enumerate Escherichia coli O157 and O26 serogroups in sheep recto-anal swabs. J. Microbiol. Methods 2019, 165, 105703. [Google Scholar] [CrossRef]
- DebRoy, C.; Roberts, E.; Valadez, A.M.; Dudley, E.G.; Cutter, C.N. Detection of Shiga toxin–producing Escherichia coli O26, O45, O103, O111, O113, O121, O145, and O157 serogroups by multiplex polymerase chain reaction of the wzx gene of the O-antigen gene cluster. Foodborne Pathog. Dis. 2011, 8, 651–652. [Google Scholar] [CrossRef]
- Fratamico, P.M.; Bagi, L.K.; Cray Jr, W.C.; Narang, N.; Yan, X.; Medina, M.; Liu, Y. Detection by multiplex real-time polymerase chain reaction assays and isolation of Shiga toxin–producing Escherichia coli serogroups O26, O45, O103, O111, O121, and O145 in ground beef. Foodborne Pathog. Dis. 2011, 8, 601–607. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Wu, L.; Zhu, S.; Long, Y.; Hang, W.; Yan, X. Rapid, absolute, and simultaneous quantification of specific pathogenic strain and total bacterial cells using an ultrasensitive dual-color flow cytometer. Anal. Chem. 2010, 82, 1109–1116. [Google Scholar] [CrossRef]
- Hegde, N.V.; Jayarao, B.M.; DebRoy, C. Rapid detection of the top six non-O157 Shiga toxin-producing Escherichia coli O groups in ground beef by flow cytometry. J. Clin. Microbiol. 2012, 50, 2137–2139. [Google Scholar] [CrossRef] [Green Version]
- Bogaerts, B.; Nouws, S.; Verhaegen, B.; Denayer, S.; Van Braekel, J.; Winand, R.; Fu, Q.; Crombé, F.; Piérard, D.; Marchal, K. Validation strategy of a bioinformatics whole genome sequencing workflow for Shiga toxin-producing Escherichia coli using a reference collection extensively characterized with conventional methods. Microb. Genom. 2021, 7. [Google Scholar] [CrossRef] [PubMed]
- Lang, C.; Hiller, M.; Konrad, R.; Fruth, A.; Flieger, A. Whole-genome-based public health surveillance of less common Shiga toxin-producing Escherichia coli serovars and untypeable strains identifies four novel O genotypes. J. Clin. Microbiol. 2019, 57, e00768-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdalhamid, B.; Mccutchen, E.L.; Bouska, A.C.; Weiwei, Z.; Loeck, B.; Hinrichs, S.H.; Iwen, P.C. Whole genome sequencing to characterize Shiga toxin-producing Escherichia coli O26 in a public health setting. J. Infect. Public Health 2019, 12, 884–889. [Google Scholar] [CrossRef]
- Castro, V.S.; Ortega Polo, R.; Figueiredo, E.E.d.S.; Bumunange, E.W.; McAllister, T.; King, R.; Conte-Junior, C.A.; Stanford, K. Inconsistent PCR detection of Shiga toxin-producing Escherichia coli: Insights from whole genome sequence analyses. PLoS ONE 2021, 16, e0257168. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Liu, Q.; Sun, H.; Xiong, Y.; Matussek, A.; Bai, X. Genomic Characterization of Escherichia coli O8 Strains Producing Shiga Toxin 2l Subtype. Microorganisms 2022, 10, 1245. [Google Scholar] [CrossRef]
- Weinroth, M.D.; Belk, A.D.; Belk, K.E. History, development, and current status of food safety systems worldwide. Anim. Front. 2018, 8, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Soon, J.; Chadd, S.; Baines, R. Escherichia coli O157: H7 in beef cattle: On farm contamination and pre-slaughter control methods. Anim. Health Res. Rev. 2011, 12, 197–211. [Google Scholar] [CrossRef]
- Callaway, T.; Edrington, T.; Loneragan, G.; Carr, M.; Nisbet, D. Current and near-market intervention strategies for reducing Shiga Toxin-Producing Escherichia coli (STEC) shedding in cattle. Agric. Food Anal. Bacteriol. 2013, 3, 103–120. [Google Scholar]
- Guth, B.E.; Prado, V.; Rivas, M. Shiga toxin-producing Escherichia coli. Pathog. Escherichia coli Lat. Am. AG Torres 2010, 65–83. [Google Scholar]
- Puligundla, P.; Lim, S. Biocontrol Approaches against Escherichia coli O157: H7 in Foods. Foods 2022, 11, 756. [Google Scholar] [CrossRef]
- Akkina, R.C.; Payala, V.; Maganti, S.S. Tools for Rapid Detection and Control of Foodborne Microbial Pathogens. In Foodborne Pathogens—Recent Advances in Control and Detection; IntechOpen: London, UK, 2022. [Google Scholar]
- Shebs-Maurine, E.; Torres, E.; Yeh-Parker, Y.; de Mello, A. Application of MS bacteriophages on contaminated trimmings reduces Escherichia coli O157 and non-O157 in ground beef. Meat Sci. 2020, 170, 108243. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Year | Cause | Country | Sources of Infection | Outcomes | References |
|---|---|---|---|---|---|
| 2019 | O103 STEC | Multistate USA outbreak | Ground Beef | 209 cases (29 hospitalizations and 2 HUS) | [123] |
| 2015 | STEC O26 | USA | Mexican grill restaurant | 55 cases (21 hospitalizations) | [124] |
| 2015 | STEC O26 | USA | Mexican grill restaurant | Five illnesses | [124] |
| 2011 | EHEC O104:H4 | Germany | Fenugreek seeds sprouts imported from Egypt | More than 4000 people (49 dead) | [125] |
| 2010 | O145 | USA | Romaine Lettuce | 26 confirmed and 7 probable cases | [126] |
| 2012 | O111:NM | Oklahoma USA | Cross-contamination between food preparation utensils or food handlers and restaurant food | 341 cases (70 hospitalizations, and 1 death) | [127] |
| 2014 | O104:H4 | Germany | Food handler contamination | 23 cases in a family party | [128] |
| 2014 | O26 | USA | Person to person spread (n = 15) or Food (n = 17) | 38 single-etiology outbreaks (66% of the total cases) | [91] |
| 2007 | O26 | Denmark | Beef sausage | 20 cases | [129] |
| 2007 | O145, O26 | Belgium | Ice cream | 12 cases (5 HUS) | [116] |
| 1995 | O111:NM | Australia | Sausage | 158 cases (23 HUS) | [130] |
| 1999 | O121 | USA (CT) | Lake water | 11 cases | [109] |
| 1990 | O111 | USA (Ohio) | Undetermined | 5 cases | [131] |
| 1999 | O111 | USA (Texas) | Salad bar | 56 cases | [89] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alharbi, M.G.; Al-Hindi, R.R.; Esmael, A.; Alotibi, I.A.; Azhari, S.A.; Alseghayer, M.S.; Teklemariam, A.D. The “Big Six”: Hidden Emerging Foodborne Bacterial Pathogens. Trop. Med. Infect. Dis. 2022, 7, 356. https://doi.org/10.3390/tropicalmed7110356
Alharbi MG, Al-Hindi RR, Esmael A, Alotibi IA, Azhari SA, Alseghayer MS, Teklemariam AD. The “Big Six”: Hidden Emerging Foodborne Bacterial Pathogens. Tropical Medicine and Infectious Disease. 2022; 7(11):356. https://doi.org/10.3390/tropicalmed7110356
Chicago/Turabian StyleAlharbi, Mona G., Rashad R. Al-Hindi, Ahmed Esmael, Ibrahim A. Alotibi, Sheren A. Azhari, Mazen S. Alseghayer, and Addisu D. Teklemariam. 2022. "The “Big Six”: Hidden Emerging Foodborne Bacterial Pathogens" Tropical Medicine and Infectious Disease 7, no. 11: 356. https://doi.org/10.3390/tropicalmed7110356