
Updating the Insecticide Resistance Status of Aedes aegypti and Aedes albopictus in Asia: A Systematic Review and Meta-Analysis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design and Search Strategy
2.2. Study Eligibility
2.3. Data Extraction and Quality Assessment
2.4. Data Analysis
3. Results
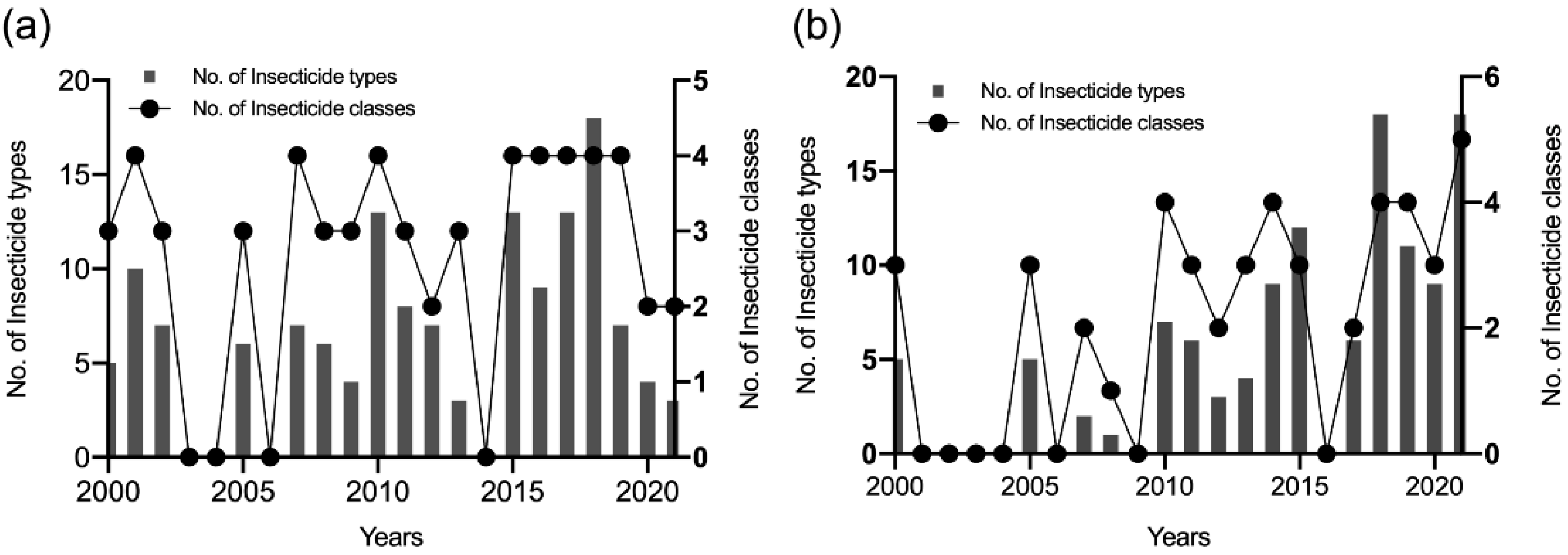
3.1. Literature Survey
3.2. Spatial Patterns of Insecticide Resistance in Asia
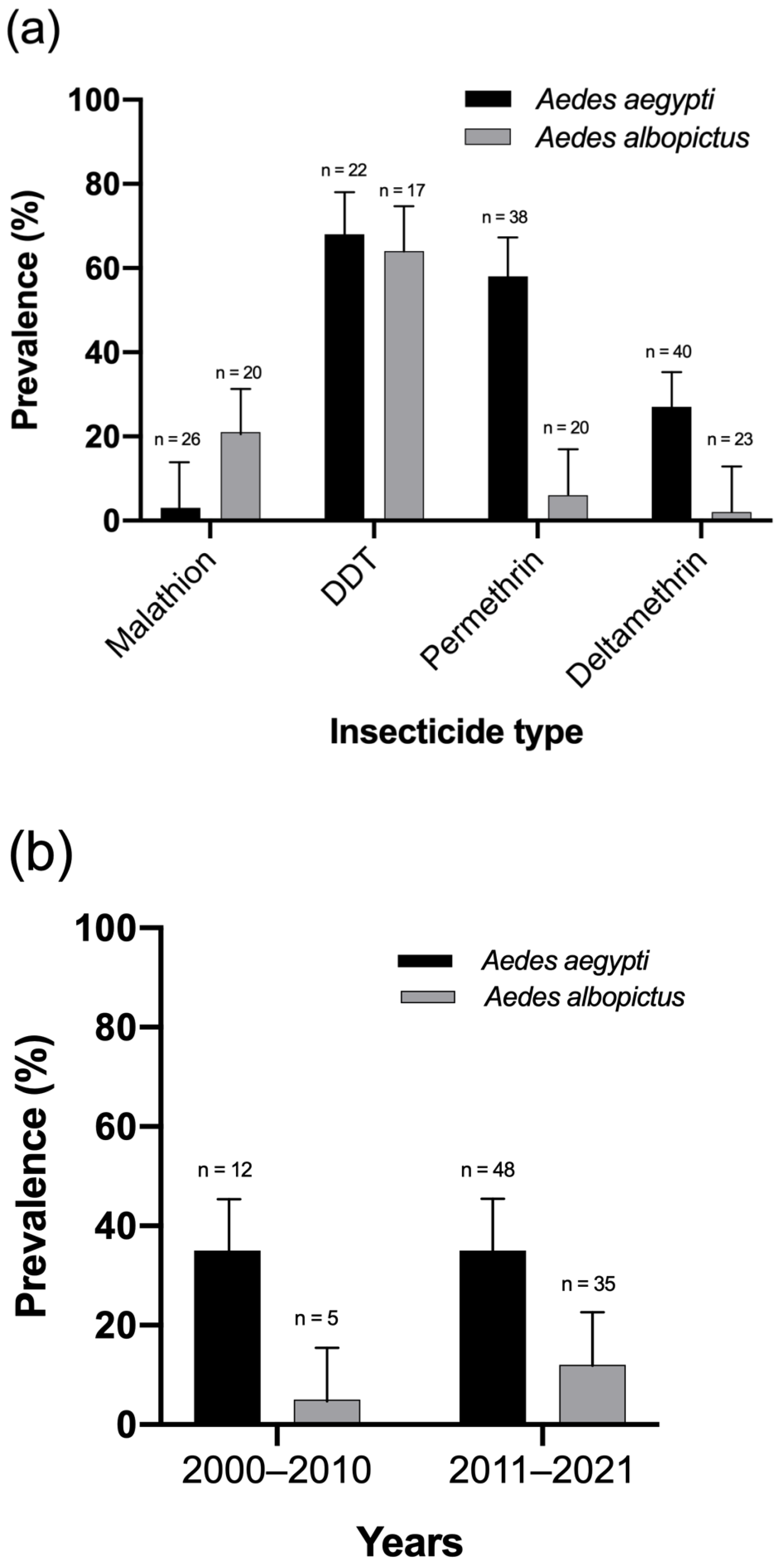
3.3. Spatial Patterns of Resistance to Malathion, DDT, Permethrin, and Deltamethrin
3.4. Prevalence of Insecticide Resistance
3.5. Heterogeneity and Publication Bias
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- World Health Organization (WHO). 2022 Vector-Borne Diseases. Available online: https://www.who.int/news-room/fact-sheets/detail/vector-borne-diseases (accessed on 4 May 2022).
- Ferguson, N.M. Challenges and opportunities in controlling mosquito-borne infections. Nature 2018, 559, 490–497. [Google Scholar] [CrossRef]
- Mordecai, E.A.; Caldwell, J.M.; Grossman, M.K.; Lippi, C.A.; Johnson, L.R.; Neira, M.; Rohr, J.S.; Ryan, S.J.; Savage, S.; Shocket, M.S.; et al. Thermal biology of mosquito-borne disease. Ecol. Lett. 2019, 22, 1690–1708. [Google Scholar] [CrossRef] [Green Version]
- Wilder-Smith, A.; Ooi, E.E.; Horstick, O.; Wills, B. Dengue. Lancet 2019, 393, 350–363. [Google Scholar] [CrossRef]
- Bhatt, S.; Gething, P.W.; Brady, O.J.; Messina, J.P.; Farlow, A.W.; Moyes, C.L.; Drake, J.M.; Brownstein, J.S.; Hoen, A.G.; Sankoh, O.; et al. The global distribution and burden of dengue. Nature 2013, 496, 504–507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stanaway, J.D.; Shepard, D.S.; Undurraga, E.A.; Halasa, Y.A.; Coffeng, L.E.; Brady, O.J.; Hay, S.I.; Bedi, N.; Bensenor, I.M.; Cast Castañeda-Orjuela, C.A.; et al. The global burden of dengue: An analysis from the global burden of disease study 2013. Lancet Infect. Dis. 2016, 16, 712–723. [Google Scholar] [CrossRef] [Green Version]
- Brady, O.J.; Gething, P.W.; Bhatt, S.; Messina, J.P.; Brownstein, J.S.; Hoen, A.G.; Moyes, C.L.; Farlow, A.W.; Scott, T.W.; Hay, S.I. Refining the global spatial limits of dengue virus transmission by evidence-based consensus. PLoS Negl. Trop. Dis. 2012, 6, e1760. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization (WHO). 2022 Dengue and Severe Dengue. Available online: https://www.who.int/news-room/fact-sheets/detail/dengue-and-severe-dengue (accessed on 4 May 2022).
- Ravi, V. Re-emergence of Chikungunya virus in India. Indian J. Med. Microbiol. 2006, 24, 83–84. [Google Scholar] [CrossRef]
- Powers, A.M.; Logue, C.H. Changing patterns of Chikungunya virus: Re-emergence of a zoonotic arbovirus. J. Gen. Virol. 2007, 88, 2363–2377. [Google Scholar] [CrossRef]
- Wimalasiri-Yapa, B.M.C.R.; Stassen, L.; Huang, X.; Hafner, L.M.; Hu, W.; Devine, G.J.; Yakob, L.; Jansen, C.C.; Faddy, H.M.; Viennet, E.; et al. Chikungunya virus in Asia–Pacific: A systematic review. Emerg. Microbes Infect. 2019, 8, 70–79. [Google Scholar] [CrossRef] [Green Version]
- Laras, K.; Sukri, N.C.; Larasati, R.P.; Bangs, M.J.; Kosim, R.; Djauzi; Wandra, J.; Master, J.; Kosasih, H.; Hartati, S.; et al. Tracking the re-emergence of epidemic Chikungunya virus in Indonesia. Trans. R. Soc. Trop. Med. Hyg. 2005, 99, 128–141. [Google Scholar] [CrossRef]
- Parola, P.; de Lamballerie, X.; Jourdan, J.; Rovery, C.; Vaillant, V.; Minodier, P.; Brouqui, P.; Flahault, A.; Raoult, D.; Charrel, R.N. Novel Chikungunya virus variant in travelers returning from Indian Ocean islands. Emerg. Infect. Dis. 2006, 12, 1493–1499. [Google Scholar] [CrossRef] [PubMed]
- Silva, L.A.; Dermody, T.S. Chikungunya virus: Epidemiology, replication, disease mechanisms, and prospective intervention strategies. J. Clin. Investig. 2017, 127, 737–749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues Faria, N.; Lourenço, J.; Marques de Cerqueira, E.; Maia de Lima, M.; Pybus, O.; Carlos Junior Alcantara, L. Epidemiology of Chikungunya virus in Bahia, brazil, 2014–2015. PLoS Curr. 2016, 8. [Google Scholar] [CrossRef] [Green Version]
- Yactayo, S.; Staples, J.E.; Millot, V.; Cibrelus, L.; Ramon-Pardo, P. Epidemiology of chikungunya in the Americas. J. Infect. Dis. 2016, 214 (Suppl. S5), S441–S445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cunha, M.S.; Costa, P.A.G.; Correa, I.A.; de Souza, M.R.M.; Calil, P.T.; da Silva, G.P.D.; Costa, S.M.; Fonseca, V.W.P.; da Costa, L.J. Chikungunya virus: An emergent arbovirus to the South American continent and a continuous threat to the world. Front. Microbiol. 2020, 11, 1297. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention (CDC). 2022 Chikungunya Virus. Available online: https://www.cdc.gov/chikungunya/index.html (accessed on 4 May 2022).
- Pan American; Health Organization (PAHO). 2021 Chikungunya: Epidemiological Alerts and Updates. Available online: https://www3.paho.org/hq/index.php?option=com_topics&view=rdmore&cid=5855&Itemid=40931&lang=en (accessed on 4 May 2022).
- World Health Organization (WHO). 2021 Chikungunya. Available online: https://www.who.int/health-topics/chikungunya#tab=tab_1 (accessed on 4 May 2022).
- World Health Organization (WHO). 2016 Zika Virus, Microcephaly and Guillain–Barre´ Syndrome; WHO: Geneva, Switzerland. Available online: http://www.who.int/emergencies/zika-virus/situation-report/1-september-2016/en/ (accessed on 4 May 2022).
- Torres, J.R.; Murillo, J.; Boffil, L. The ever changing lanscape of Zika virs infection. Learning on the fly. Int. J. Infect. Dis. 2016, 51, 123–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Europian Center for Disease Prevention and Control (ECDC). 2018 Local Transmission of Dengue Fever in France and Spain. Available online: https://www.ecdc.europa.eu/en/publications-data/rapid-risk-assessment-local-transmission-dengue-fever-france-and-spain (accessed on 4 May 2022).
- Radke, E.G.; Gregory, C.J.; Kintziger, K.W.; Sauber-Schatz, E.K.; Hunsperger, E.A.; Gallagher, G.R.; Barber, J.M.; Biggerstaff, B.J.; Stannek, D.R.; Thomasek, K.M.; et al. Dengue outbreak in Key West, Florida, USA, 2009. Emerg. Infect. Dis. 2012, 18, 135–137. [Google Scholar] [CrossRef]
- Furuya, H. Estimation of reproduction number and probable vector density of the first autochthonous dengue outbreak in Japan in the last 70 years. Environ. Health Prev. Med. 2015, 20, 466–471. [Google Scholar] [CrossRef] [Green Version]
- Pan American Health Organization (PAHO). 2015 Number of Reported Cases of CHIK Fever in the Americas by Country or 481 Territory 2013–2014, and 2015. Available online: https://www3.paho.org/hq/index.php?option=com_topics&view=rdmore&cid=5927&Itemid=358&lang=en (accessed on 4 May 2022).
- Mascarenhas, M.; Garasia, S.; Berthiaume, P.; Corrin, T.; Greig, J.; Ng, V.; Young, I.; Waddell, L. A scoping review of published literature on Chikungunya virus. PLoS ONE 2018, 13, e0207554. [Google Scholar] [CrossRef] [PubMed]
- Padbidri, V.S.; Gnaneswar, T.T. Epidemiological investigations of chikungunya epidemic at Barsi, Maharashtra state, India. J. Hyg. Epidemiol. Microbiol. Immunol. 1979, 23, 445–451. [Google Scholar] [PubMed]
- Hapuarachchi, H.C.; Bandara, K.B.A.T.; Sumanadasa, S.D.M.; Hapugoda, M.D.; Lai, Y.L.; Lee, K.S.; Tan, L.K.; Lin, R.T.P.; Ng, L.F.P.; Bucht, G.; et al. Re-emergence of Chikungunya virus in South-East Asia: Virological evidence from Sri Lanka and Singapore. J. Gen. Virol. 2010, 91, 1067–1076. [Google Scholar] [CrossRef]
- Manimunda, S.P.; Sugunan, A.P.; Rai, S.K.; Vijayachari, P.; Shriram, A.N.; Sharma, S.; Muruganandam, N.; Chaitanya, I.K.; Guruprasad, D.R.; Sudeep, A.B. Outbreak of chikungunya fever, Dakshina Kannada District, South India, 2008. Am. J. Trop. Med. Hyg. 2010, 83, 751–754. [Google Scholar] [CrossRef] [Green Version]
- Haque, F.; Rahman, M.; Banu, N.N.; Sharif, A.R.; Jubayer, S.; Shamsuzzaman, A.K.M.; Alamgir, A.S.M.; Erasmus, J.H.; Guzman, H.; Forrester, N.; et al. An epidemic of chikungunya in northwestern Bangladesh in 2011. PLoS ONE 2019, 14, e0212218. [Google Scholar] [CrossRef] [Green Version]
- Vairo, F.; Haider, N.; Kock, R.; Ntoumi, F.; Ippolito, G.; Zumla, A. Chikungunya: Epidemiology, pathogenesis, clinical features, management, and prevention. Infect. Dis. Clin. N. Am. 2019, 33, 1003–1025. [Google Scholar] [CrossRef]
- Dick, G.W.; Kitchen, S.F.; Haddow, A.J. Zika virus. I. Isolations and serological specificity. Trans. R. Soc. Trop. Med. Hyg. 1952, 46, 509–520. [Google Scholar] [CrossRef]
- Gatherer, D.; Kohl, A. Zika virus: A previously slow pandemic spreads rapidly through the Americas. J. Gen. Virol. 2016, 97, 269–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization (WHO). 2022 Zika Virus Disease Outbreak 2015–2016. Available online: https://www.who.int/emergencies/situations/zika-virus-outbreak (accessed on 4 May 2022).
- European Centre for Disease Prevention and Control (ECDC). 2016 Epidemiological Update: Outbreaks of Zika Virus and Complications Potentially Linked to the Zika Virus Infection. Available online: https://www.ecdc.europa.eu/en/news-events/epidemiological-update-outbreaks-zika-virus-and-complications-potentially-linked-23 (accessed on 4 May 2022).
- Alera, M.T.; Hermann, L.; Tac-An, I.A.; Klungthong, C.; Rutvisuttinunt, W.; Manasatienkij, W.; Villa, D.; Thaisomboosuk, B.; Velasco, J.H.; Chinnawirotpisan, P.; et al. Zika virus infection, Philippines, 2012. Emerg. Infect. Dis. 2015, 21, 722–724. [Google Scholar] [CrossRef] [PubMed]
- Quyen, N.T.H.; Kien, D.T.H.; Rabaa, M.; Tuan, N.M.; Vi, T.T.; Van Tan, L.; Hung, N.T.; Tuan, H.M.; Tram, T.V.; Da Ha, N.L.; et al. Chikungunya and Zika virus cases detected against a backdrop of endemic dengue transmission in Vietnam. Am. J. Trop. Med. Hyg. 2017, 97, 146–150. [Google Scholar] [CrossRef] [PubMed]
- Kwong, J.C.; Druce, J.D.; Leder, K. Zika virus infection acquired during brief travel to Indonesia. Am. J. Trop. Med. Hyg. 2013, 89, 516–517. [Google Scholar] [CrossRef] [Green Version]
- Leung, G.H.; Baird, R.W.; Druce, J.; Anstey, N.M. Zika virus infection in Australia following a monkey bite in Indonesia. Southeast Asian J. Trop. Med. Public Health 2015, 46, 460–464. [Google Scholar] [PubMed]
- Perkasa, A.; Yudhaputri, F.; Haryanto, S.; Hayati, R.F.; Ma’roef, C.N.; Antonjaya, U.; Yohan, B.; Myint, K.S.A.; Ledermann, P.; Rosendberg, R.; et al. Isolation of Zika virus from febrile patient, Indonesia. Emerg. Infect. Dis. 2016, 22, 924–925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tappe, D.; Nachtigall, S.; Kapaun, A.; Schnitzler, P.; Günther, S.; Schmidt-Chanasit, J. Acute Zika virus infection after travel to Malaysian Borneo, September 2014. Emerg. Infect. Dis. 2015, 21, 911–913. [Google Scholar] [CrossRef] [Green Version]
- Nitatpattana, N.; Chaiyo, K.; Rajakam, S.; Poolam, K.; Chansiprasert, K.; Pesirikan, N.; Buree, S.; Rodpai, E.; Yoksan, S. Complete genome sequence of a Zika virus strain isolated from the serum of an infected patient in Thailand in 2006. Genome Announc. 2018, 6, e00121-18. [Google Scholar] [CrossRef] [Green Version]
- European Medicines Agency. 2022 Dengvaxia. Available online: https://www.ema.europa.eu/en/medicines/human/EPAR/dengvaxia#:~:text=Dengvaxia%20is%20a%20vaccine%20used,like%20symptoms%20in%20most%20people (accessed on 4 May 2022).
- Coats, J.R. Mechanisms of toxic action and structure-activity relationships for organochlorine and synthetic pyrethroid insecticides. Environ. Health Perspect. 1990, 87, 255–262. [Google Scholar] [CrossRef]
- Davies, T.G.; Field, L.M.; Usherwood, P.N.; Williamson, M.S. DDT, pyrethrins, pyrethroids and insect sodium channels. IUBMB Life 2007, 59, 151–162. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, L.J.; Casida, J.E. Stereospecific action of pyrethroid insecticides on the gamma-aminobutyric acid receptor-ionophore complex. Science 1983, 221, 1399–1401. [Google Scholar] [CrossRef]
- Bloomquist, J.R.; Soderlund, D.M. Neurotoxic insecticides inhibit GABA-dependent chloride uptake by mouse brain vesicles. Biochem. Biophys. Res. Commun. 1985, 133, 37–43. [Google Scholar] [CrossRef]
- Fukuto, T.R. Mechanism of action of organophosphorus and carbamate insecticides. Environ. Health Perspect. 1990, 87, 245–254. [Google Scholar] [CrossRef]
- Gerba, C.P. Chapter 28 Environmental toxicology. In Environmental and Pollution Science, 3rd ed.; Brusseau, M.L., Pepper, I.L., Gerba, C.P., Eds.; Academic Press: London, UK, 2019; pp. 511–540. [Google Scholar]
- Costa, L.G. Current issues in organophosphate toxicology. Clin. Chim. Acta 2006, 366, 1–13. [Google Scholar] [CrossRef]
- Soderlund, D.M. Molecular mechanisms of pyrethroid insecticide neurotoxicity: Recent advances. Arch. Toxicol. 2012, 86, 165–181. [Google Scholar] [CrossRef]
- Breckenridge, C.B.; Holden, L.; Sturgess, N.; Weiner, M.; Sheets, L.; Sargent, D.; Soderlund, D.M.; Choi, J.S.; Symington, S.; Clark, J.M.; et al. Evidence for a separate mechanism of toxicity for the type I and the type II pyrethroid insecticides. Neurotoxicology 2009, 30 (Suppl. S1), S17–S31. [Google Scholar] [CrossRef] [PubMed]
- Gajendiran, A.; Abraham, J. An overview of pyrethroid insecticides. Front. Biol. 2018, 13, 79–90. [Google Scholar] [CrossRef]
- Singh, A.K.; Tiwari, M.N.; Prakash, O.; Singh, M.P. A current review of cypermethrin-induced neurotoxicity and nigrostriatal dopaminergic neurodegeneration. Curr. Neuropharmacol. 2012, 10, 64–71. [Google Scholar] [CrossRef] [Green Version]
- Insecticide Resistance Action Committee (IRAC). 2022 Interactive MoA Classification. Available online: https://irac-online.org/modes-of-action/ (accessed on 4 May 2022).
- Heckel, D.G. Learning the ABCs of Bt: ABC transporters and insect resistance to Bacillus thuringiensis provide clues to a crucial step in toxin mode of action. Pestic. Biochem. Physiol. 2012, 104, 103–110. [Google Scholar] [CrossRef]
- Risher, J.F.; Mink, F.L.; Stara, J.F. The toxicologic effects of the carbamate insecticide aldicarb in mammals: A review. Environ. Health Perspect. 1987, 72, 267–281. [Google Scholar] [CrossRef] [PubMed]
- Chevillon, C.; Raymond, M.; Guillemaud, T.; Lenormand, T.; Pasteur, N. Population genetics of insecticide resistance in the mosquito Culex pipiens. Biol. J. Linn. Soc. Lond. 1999, 68, 147–157. [Google Scholar] [CrossRef]
- Hemingway, J.; Hawkes, N.J.; McCarroll, L.; Ranson, H. The molecular basis of insecticide resistance in mosquitoes. Insect Biochem. Mol. Biol. 2004, 34, 653–665. [Google Scholar] [CrossRef] [PubMed]
- Liu, N. Insecticide resistance in mosquitoes: Impact, mechanisms, and research directions. Annu. Rev. Entomol. 2015, 60, 537–559. [Google Scholar] [CrossRef] [PubMed]
- Silver, K.S.; Du, Y.; Nomura, Y.; Oliveira, E.E.; Salgado, V.L.; Zhorov, B.S.; Dong, K. Voltage-gated sodium channels as insecticide targets. Adv. Insect Physiol. 2014, 46, 389–433. [Google Scholar]
- Balabanidou, V.; Grigoraki, L.; Vontas, J. Insect cuticle: A critical determinant of insecticide resistance. Curr. Opin. Insect Sci. 2018, 27, 68–74. [Google Scholar] [CrossRef]
- Kyle, J.L.; Haris, E. Global spread and persistence of dengue. Ann. Rev. Microbiolog. 2008, 62, 71–92. [Google Scholar] [CrossRef] [PubMed]
- Gubler, D.J. Dengue, urbanization and globalization: The unholy trinity of the 21st dentury. Trop. Med. Health 2011, 39 (Suppl. S4), S3–S11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Focks, D.A.; Haile, D.; Daniel, E.; Mount, G.A. Dynamic life table model fo Aedes aegypti (Diptera: Culicidae): Analysis of the literature and model development. J. Med. Entomol. 1993, 30, 1003–1017. [Google Scholar] [CrossRef] [PubMed]
- Morin, C.W.; Comrie, A.C.; Ernst, K. Climate and dengue transmission: Evidence and implications. Environ. Health Perspect 2013, 121, 1264–1272. [Google Scholar] [CrossRef] [PubMed]
- Moyes, C.L.; Vontas, J.; Martins, A.J.; Ng, L.C.; Koou, S.Y.; Dusfour, I.; Raghavendra, K.; Pinto, J.; Corbel, V.; David, J.P.; et al. Contemporary status of insecticide resistance in the major Aedes vectors of arboviruses infecting humans. PLoS Negl. Trop. Dis. 2017, 11, e0005625. [Google Scholar] [CrossRef] [PubMed]
- Haziqah-Rashid, A.; Chen, C.D.; Lau, K.W.; Low, V.L.; Sofian-Azirun, M.; Suana, I.W.; Harmonis, H.; Syahputra, E.; Razak, A.; Chin, A.C.; et al. Monitoring insecticide resistance profiles of Aedes aegypti (Diptera: Culicidae) in the Sunda Islands of Indonesia based on diagnostic doses of larvicides. J. Med. Entomol. 2019, 56, 514–518. [Google Scholar] [CrossRef]
- Mulyatno, K.C.; Yamanaka, A.; Ngadino; Konishi, E. Resistance of Aedes aegypti (L.) larvae to temephos in Surabaya, Indonesia. Southeast Asian J. Trop. Med. Public Health 2012, 43, 29–33. [Google Scholar] [PubMed]
- Rasli, R.; Lee, H.L.; Wasi Ahmad, N.; Fikri, S.F.F.; Ali, R.; Muhamed, K.A.; Hadi, A.A.; Liu, Q.Y.; Meng, F.X. Susceptibility status and resistance mechanisms in permethrin-selected, laboratory susceptible and field-collected Aedes aegypti from Malaysia. Insects 2018, 9, 43. [Google Scholar] [CrossRef] [Green Version]
- Leong, C.S.; Vythilingam, I.; Wong, M.L.; Wan Sulaiman, W.Y.; Lau, Y.L. Aedes aegypti (Linnaeus) larvae from dengue outbreak areas in Selangor showing resistance to pyrethroids but susceptible to organophosphates. Acta Trop. 2018, 185, 115–126. [Google Scholar] [CrossRef]
- Abu Hasan, H.; Jaal, Z.; Ranson, H.; McCall, P. Pyrethroid and organophosphate susceptibility status of Aedes aegypti (Linnaeus) and Aedes albopictus (Skuse) in Penang, Malaysia. Int. J. Entomol. Res. 2015, 3, 91–95. [Google Scholar]
- Stenhouse, S.A.; Plernsub, S.; Yanola, J.; Lumjuan, N.; Dantrakool, A.; Choochote, W.; Somboon, P. Detection of the V1016G mutation in the voltage-gated sodium channel gene of Aedes aegypti (Diptera: Culicidae) by allele-specific PCR assay, and its distribution and effect on deltamethrin resistance in Thailand. Parasites Vectors 2013, 6, 253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Somboon, P.; Prapanthadara, L.A.; Suwonkerd, W. Insecticide susceptibility tests of Anopheles minimus s.l., Aedes aegypti, Aedes albopictus, and Culex quinquefasciatus in northern Thailand. Southeast Asian J. Trop. Med. Public Health 2003, 34, 87–93. [Google Scholar] [PubMed]
- Jirakanjanakit, N.; Rongnoparut, P.; Saengtharatip, S.; Chareonviriyaphap, T.; Duchon, S.; Bellec, C.; Yoksan, S. Insecticide susceptible/resistance status in Aedes (stegomyia) aegypti and Aedes (stegomyia) albopictus (Diptera: Culicidae) in Thailand during 2003–2005. J. Econ. Entomol. 2007, 100, 545–550. [Google Scholar]
- Thongwat, D.; Bunchu, N. Susceptibility to temephos, permethrin and deltamethrin of Aedes aegypti (Diptera: Culicidae) from Muang district, Phitsanulok Province, Thailand. Asian Pac. J. Trop. Med. 2015, 8, 14–18. [Google Scholar] [CrossRef] [Green Version]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G.; The PRISMA Group. Preferred reporting items for systematic reviews and metaanalyses: The PRISMA statement. PLoS Med. 2009, 6, e1000097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stroup, D.F.; Berlin, J.A.; Morton, S.C.; Olkin, I.; Williamson, G.D.; Rennie, D.; Moher, D.; Becker, B.J.; Sipe, T.A.; Tacker, S.B. Meta-analysis of observational studies in epidemiology: A proposal for reporting. Meta-analysis Of Observational Studies in Epidemiology (MOOSE) group. JAMA 2000, 283, 2008–2012. [Google Scholar] [CrossRef] [PubMed]
- Hoy, D.; Brooks, P.; Woolf, A.; Blyth, F.; March, L.; Bain, C.; Baker, P.; Smith, E.; Buchbinder, R. Assessing risk of bias in prevalence studies: Modification of an existing tool and evidence of interrater agreement. J. Clin. Epidemiol. 2012, 65, 934–939. [Google Scholar] [CrossRef]
- Borenstein, M.; Hedges, L.V.; Higgins, J.P.T.; Rothstein, H.R. Introduction to meta-analysis. Chichester 2021, 77, 478–479. [Google Scholar]
- Furuya-Kanamori, L.; Barendregt, J.J.; Doi, S.A.R. A new improved graphical and quantitative method for detecting bias in meta-analysis. Int. J. Evid.-Based Healthc. 2018, 16, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Barendregt, J.J.; Doi, S.A. MetaXL User Guide; EpiGear International Pty Ltd.: Sunrise Beach, Australia, 2016; Available online: http://www.epigear.com/index_files/metaxl.html (accessed on 4 May 2022).
- Lim, J.T.; Dickens, B.S.L.; Chew, L.Z.X.; Choo, E.L.W.; Koo, J.R.; Aik, J.; Ng, L.C.; Cook, A.R. Impact of sars-cov-2 interventions on dengue transmission. PLoS Negl. Trop. Dis. 2020, 14, e0008719. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.T.; Chew, L.Z.X.; Choo, E.L.W.; Dickens, B.S.L.; Ong, J.; Aik, J.; Ng, L.C.; Cook, A.R. Increased dengue transmissions in Singapore attributable to SARS-CoV-2 social distancing measures. J. Infect. Dis. 2021, 223, 399–402. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Yang, C.; Liu, N.; Li, M.; Tong, Y.; Zeng, X.; Qiu, X. Knockdown resistance (kdr) mutations within seventeen field populations of Aedes albopictus from Beijing China: First report of a novel V1016G mutation and evolutionary origins of kdr haplotypes. Parasites Vectors 2019, 12, 180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bisset, J.; Rodríguez, M.M.; Fernández, D. Selection of insensitive acetylcholinesterase as a resistance mechanism in Aedes aegypti (Diptera: Culicidae) from Santiago de Cuba. J. Med. Entomol. 2006, 43, 1185–1189. [Google Scholar] [CrossRef] [PubMed]
- Polson, K.A.; Brogdon, W.G.; Rawlins, S.C.; Chadee, D.D. Characterization of insecticide resistance in Trinidadian strains of Aedes aegypti mosquitoes. Acta Trop. 2011, 117, 31–38. [Google Scholar] [CrossRef]
- Aktar, M.W.; SenGupta, D.; Chowdhury, A. Impact of pesticides use in agriculture: Their benefits and hazards. Interdiscip. Toxicol. 2009, 2, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaur, H.; Garg, H. Pesticides: Environmental impacts and management strategies. Pestic. Toxic Asp. 2014, 8, 187. [Google Scholar]
- Guruprasad, N.M.; Jalali, S.K.; Puttaraju, H.P. Wolbachia—A foe for mosquitoes. Asian Pac. J. Trop. Dis. 2014, 4, 78–81. [Google Scholar] [CrossRef]
- Utarini, A.; Indriani, C.; Ahmad, R.A.; Tantowijoyo, W.; Arguni, E.; Ansari, M.R.; Supriyati, E.; Wardana, D.S.; Meitika, Y.; Ernesia, I.; et al. Efficacy of Wolbachia-infected mosquito deployments for the control of dengue. N. Engl. J. Med. 2021, 384, 2177–2186. [Google Scholar] [CrossRef]
- Pinto, S.B.; Riback, T.I.S.; Sylvestre, G.; Costa, G.; Peixoto, J.; Dias, F.B.S.; Tanamas, S.K.; Simmons, C.P.; Dufault, S.M.; Ryan, P.A.; et al. Effectiveness of Wolbachia-infected mosquito deployments in reducing the incidence of dengue and other Aedes-borne diseases in Niterói, Brazil: A quasi-experimental study. PLoS Negl. Trop. Dis. 2021, 15, e0009556. [Google Scholar] [CrossRef]
- O’Neill, S.L.; Ryan, P.A.; Turley, A.P.; Wilson, G.; Retzki, K.; Iturbe-Ormaetxe, I.; Dong, Y.; Kenny, N.; Paton, C.J.; Ritchie, S.A.; et al. Scaled deployment of Wolbachia to protect the community from dengue and other Aedes transmitted arboviruses. Gates Open Res. 2018, 2, 36. [Google Scholar] [CrossRef]
- Ryan, P.A.; Turley, A.P.; Wilson, G.; Hurst, T.P.; Retzki, K.; Brown-Kenyon, J.; Hodgson, L.; Kenny, N.; Cook, H.; Montgomery, B.L.; et al. Establishment of wMel Wolbachia in Aedes aegypti mosquitoes and reduction of local dengue transmission in Cairns and surrounding locations in northern Queensland, Australia. Gates Open Res. 2019, 3, 1547. [Google Scholar] [CrossRef] [PubMed]
- Tantowijoyo, W.; Tanamas, S.K.; Nurhayati, I.; Setyawan, S.; Budiwati, N.; Fitriana, I.; Ernesia, I.; Wardana, D.S.; Supriyati, E.; Arguni, E.; et al. Aedes aegypti abundance and insecticide resistance profiles in the applying Wolbachia to eliminate dengue trial. PLoS Negl. Trop. Dis. 2022, 16, e0010284. [Google Scholar] [CrossRef] [PubMed]
- Hamid, P.H.; Prastowo, J.; Ghiffari, A.; Taubert, A.; Hermosilla, C. Aedes aegypti resistance development to commonly used insecticides in Jakarta, Indonesia. PLoS ONE 2017, 12, e0189680. [Google Scholar] [CrossRef] [PubMed]
- Hasmiwati, H.; Rusjdi, S.; Nofita, E. Detection of Ace-1 gene with insecticides resistance in Aedes aegypti populations from DHF-endemic areas in Padang, Indonesia. Biodivers. J. Biol. Divers. 2018, 19, 31–36. [Google Scholar] [CrossRef]
- Hamid, P.; Ninditya, V.; Prastowo, J.; Haryanto, A.; Taubert, A.; Hermosilla, C. Current Status of Aedes aegypti Insecticide Resistance Development from Banjarmasin, Kalimantan, Indonesia. BioMed Res. Int. 2018, 2018, 1735358. [Google Scholar] [CrossRef]
- Wuliandari, J.; Hoffmann, A.; Tantowijoyo, W.; Endersby-Harshman, N. Frequency of kdr mutations in the voltage-sensitive sodium channel (VSSC) gene in Aedes aegypti from Yogyakarta and implications for Wolbachia-infected mosquito trials. Parasites Vectors 2020, 13, 429. [Google Scholar] [CrossRef]
- Hasmiwati, H.; Supargiyono, S. Short Communication: Genotyping of kdr allele in insecticide resistant-Aedes aegypti populations from West Sumatra, Indonesia. Biodivers. J. Biol. Divers. 2018, 19, 502–508. [Google Scholar] [CrossRef]
- Srisawat, R.; Komalamisra, N.; Eshita, Y.; Zheng, M.; Ono, K.; Itoh, T.; Matsumoto, A.; Petmitr, S.; Rongsriyam, Y. Point mutations in domain II of the voltage-gated sodium channel gene in deltamethrin-resistant Aedes aegypti (Diptera: Culicidae). Appl. Entomol. Zool. 2010, 45, 275–282. [Google Scholar] [CrossRef] [Green Version]
- Yanola, J.; Somboon, P.; Walton, C.; Nachaiwieng, W.; Somwang, P.; Prapanthadara, L. High-throughput assays for detection of the F1534C mutation in the voltage-gated sodium channel gene in permethrin-resistant Aedes aegypti and the distribution of this mutation throughout Thailand. Trop. Med. Int. Health 2011, 16, 501–509. [Google Scholar] [CrossRef]
- Rajatileka, S.; Black, W.; Saavedra-Rodriguez, K.; Trongtokit, Y.; Apiwathnasorn, C.; McCall, P.; Ranson, H. Development and application of a simple colorimetric assay reveals widespread distribution of sodium channel mutations in Thai populations of Aedes aegypti. Acta Trop. 2008, 108, 54–57. [Google Scholar] [CrossRef] [PubMed]
- Saingamsook, J.; Saeung, A.; Yanola, J.; Lumjuan, N.; Walton, C.; Somboon, P. A multiplex PCR for detection of knockdown resistance mutations, V1016G and F1534C, in pyrethroid-resistant Aedes aegypti. Parasites Vectors 2017, 10, 465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leong, C.; Vythilingam, I.; Liew, J.; Wong, M.; Wan-Yusoff, W.; Lau, Y. Enzymatic and molecular characterization of insecticide resistance mechanisms in field populations of Aedes aegypti from Selangor, Malaysia. Parasites Vectors 2019, 12, 236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tangena, J.; Marcombe, S.; Thammavong, P.; Chonephetsarath, S.; Somphong, B.; Sayteng, K.; Grandadam, M.; Sutherland, I.; Lindsay, S.; Brey, P. Bionomics and insecticide resistance of the arboviral vector Aedes albopictus in northern Lao PDR. PLoS ONE 2018, 13, e0206387. [Google Scholar] [CrossRef] [Green Version]
- Kawada, H.; Higa, Y.; Komagata, O.; Kasai, S.; Tomita, T.; Thi Yen, N.; Loan, L.; Sánchez, R.; Takagi, M. Widespread Distribution of a Newly Found Point Mutation in Voltage-Gated Sodium Channel in Pyrethroid-Resistant Aedes aegypti Populations in Vietnam. PLoS Negl. Trop. Dis. 2009, 3, e527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kushwah RB, S.; Dykes, C.L.; Kapoor, N.; Adak, T.; Singh, O.P. Pyrethroid- Resistance and Presence of Two Knockdown Resistance (kdr) Mutations, F1534C and a Novel Mutation T1520I, in Indian Aedes aegypti. PLOS Negl. Trop. Dis. 2015, 9, e3332. [Google Scholar] [CrossRef]
- Saha, P.; Chatterjee, M.; Ballav, S.; Chowdhury, A.; Basu, N.; Maji, A. Prevalence of kdr mutations and insecticide susceptibility among natural population of Aedes aegypti in West Bengal. PLoS ONE 2019, 14, e0215541. [Google Scholar] [CrossRef]
- Kushwah, R.; Kaur, T.; Dykes, C.; Ravi Kumar, H.; Kapoor, N.; Singh, O. A new knockdown resistance (kdr) mutation, F1534L, in the voltage-gated sodium channel of Aedes aegypti, co-occurring with F1534C, S989P and V1016G. Parasites Vectors 2020, 13, 327. [Google Scholar] [CrossRef]
- Fernando, S.; Hapugoda, M.; Perera, R.; Saavedra-Rodriguez, K.; Black, W.; De Silva, N. First report of V1016G and S989P knockdown resistant (kdr) mutations in pyrethroid-resistant Sri Lankan Aedes aegypti mosquitoes. Parasites Vectors 2018, 11, 526. [Google Scholar] [CrossRef] [Green Version]
- Chung, H.; Cheng, I.; Chen, Y.; Lin, C.; Tomita, T.; Teng, H. Voltage-gated sodium channel intron polymorphism and four mutations comprise six haplotypes in an Aedes aegypti population in Taiwan. PLoS Negl. Trop. Dis. 2019, 13, e0007291. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zulfa, R.; Lo, W.-C.; Cheng, P.-C.; Martini, M.; Chuang, T.-W. Updating the Insecticide Resistance Status of Aedes aegypti and Aedes albopictus in Asia: A Systematic Review and Meta-Analysis. Trop. Med. Infect. Dis. 2022, 7, 306. https://doi.org/10.3390/tropicalmed7100306
Zulfa R, Lo W-C, Cheng P-C, Martini M, Chuang T-W. Updating the Insecticide Resistance Status of Aedes aegypti and Aedes albopictus in Asia: A Systematic Review and Meta-Analysis. Tropical Medicine and Infectious Disease. 2022; 7(10):306. https://doi.org/10.3390/tropicalmed7100306
Chicago/Turabian StyleZulfa, Ramdan, Wei-Cheng Lo, Po-Ching Cheng, Martini Martini, and Ting-Wu Chuang. 2022. "Updating the Insecticide Resistance Status of Aedes aegypti and Aedes albopictus in Asia: A Systematic Review and Meta-Analysis" Tropical Medicine and Infectious Disease 7, no. 10: 306. https://doi.org/10.3390/tropicalmed7100306