Role of Host and Parasite MIF Cytokines during Leishmania Infection

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Brief Overview of Immune Response to Leishmania
2. Function and Activity of Mammalian MIF
3. Role of Host MIF during Leishmaniasis
4. Overview of MIF Orthologs in Leishmania and Other Parasites
5. Targeting MIF for Therapeutic Applications
6. Summary
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Khamesipour, A.; Dowlati, Y.; Asilian, A.; Hashemi-Fesharki, R.; Javadi, A.; Noazin, S.; Modabber, F. Leishmanization: Use of an old method for evaluation of candidate vaccines against leishmaniasis. Vaccine 2005, 23, 3642–3648. [Google Scholar] [CrossRef] [PubMed]
- Peters, N.C.; Egen, J.G.; Secundino, N.; Debrabant, A.; Kimblin, N.; Kamhawi, S.; Lawyer, P.; Fay, M.P.; Germain, R.N.; Sacks, D. In vivo imaging reveals an essential role for neutrophils in leishmaniasis transmitted by sand flies. Science 2008, 321, 970–974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peters, N.C.; Sacks, D.L. The impact of vector-mediated neutrophil recruitment on cutaneous leishmaniasis. Cell. Microbiol. 2009, 11, 1290–1296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aga, E.; Katschinski, D.M.; van Zandbergen, G.; Laufs, H.; Hansen, B.; Muller, K.; Solbach, W.; Laskay, T. Inhibition of the spontaneous apoptosis of neutrophil granulocytes by the intracellular parasite Leishmania major. J. Immunol. 2002, 169, 898–905. [Google Scholar] [CrossRef] [Green Version]
- Sarkar, A.; Aga, E.; Bussmeyer, U.; Bhattacharyya, A.; Moller, S.; Hellberg, L.; Behnen, M.; Solbach, W.; Laskay, T. Infection of neutrophil granulocytes with Leishmania major activates ERK 1/2 and modulates multiple apoptotic pathways to inhibit apoptosis. Med. Microbiol. Immunol. 2013, 202, 25–35. [Google Scholar] [CrossRef]
- Kaye, P.; Scott, P. Leishmaniasis: Complexity at the host-pathogen interface. Nat. Rev. Microbiol. 2011, 9, 604–615. [Google Scholar] [CrossRef]
- De Trez, C.; Magez, S.; Akira, S.; Ryffel, B.; Carlier, Y.; Muraille, E. iNOS-producing inflammatory dendritic cells constitute the major infected cell type during the chronic Leishmania major infection phase of C57BL/6 resistant mice. PLoS Pathog. 2009, 5, e1000494. [Google Scholar] [CrossRef]
- Engwerda, C.R.; Ato, M.; Kaye, P.M. Macrophages, pathology and parasite persistence in experimental visceral leishmaniasis. Trends Parasitol. 2004, 20, 524–530. [Google Scholar] [CrossRef]
- Sacks, D.; Noben-Trauth, N. The immunology of susceptibility and resistance to Leishmania major in mice. Nat. Rev. Immunol. 2002, 2, 845–858. [Google Scholar] [CrossRef]
- Herbst, S.; Schaible, U.E.; Schneider, B.E. Interferon gamma activated macrophages kill mycobacteria by nitric oxide induced apoptosis. PLoS ONE 2011, 6, e19105. [Google Scholar] [CrossRef] [Green Version]
- Naderer, T.; McConville, M.J. The Leishmania-macrophage interaction: A metabolic perspective. Cell. Microbiol. 2008, 10, 301–308. [Google Scholar] [CrossRef]
- Varin, A.; Gordon, S. Alternative activation of macrophages: Immune function and cellular biology. Immunobiology 2009, 214, 630–641. [Google Scholar] [CrossRef]
- Soong, L. Modulation of dendritic cell function by Leishmania parasites. J. Immunol. 2008, 180, 4355–4360. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, L.P.; Pearce, E.J.; Scott, P. Functional dichotomy of dendritic cells following interaction with Leishmania braziliensis: Infected cells produce high levels of TNF-alpha, whereas bystander dendritic cells are activated to promote T cell responses. J. Immunol. 2008, 181, 6473–6480. [Google Scholar] [CrossRef] [Green Version]
- Malherbe, L.; Filippi, C.; Julia, V.; Foucras, G.; Moro, M.; Appel, H.; Wucherpfennig, K.; Guery, J.C.; Glaichenhaus, N. Selective activation and expansion of high-affinity CD4+ T cells in resistant mice upon infection with Leishmania major. Immunity 2000, 13, 771–782. [Google Scholar] [CrossRef] [Green Version]
- Akiba, H.; Miyahira, Y.; Atsuta, M.; Takeda, K.; Nohara, C.; Futagawa, T.; Matsuda, H.; Aoki, T.; Yagita, H.; Okumura, K. Critical contribution of OX40 ligand to T helper cell type 2 differentiation in experimental leishmaniasis. J. Exp. Med. 2000, 191, 375–380. [Google Scholar] [CrossRef]
- Belkaid, Y.; Hoffmann, K.F.; Mendez, S.; Kamhawi, S.; Udey, M.C.; Wynn, T.A.; Sacks, D.L. The role of interleukin (IL)-10 in the persistence of Leishmania major in the skin after healing and the therapeutic potential of anti-IL-10 receptor antibody for sterile cure. J. Exp. Med. 2001, 194, 1497–1506. [Google Scholar] [CrossRef] [Green Version]
- Belkaid, Y.; Piccirillo, C.A.; Mendez, S.; Shevach, E.M.; Sacks, D.L. CD4+CD25+ regulatory T cells control Leishmania major persistence and immunity. Nature 2002, 420, 502–507. [Google Scholar] [CrossRef]
- Zaph, C.; Uzonna, J.; Beverley, S.M.; Scott, P. Central memory T cells mediate long-term immunity to Leishmania major in the absence of persistent parasites. Nat. Med. 2004, 10, 1104–1110. [Google Scholar] [CrossRef]
- Muller, I.; Kropf, P.; Etges, R.J.; Louis, J.A. Gamma interferon response in secondary Leishmania major infection: Role of CD8+ T cells. Infect. Immun. 1993, 61, 3730–3738. [Google Scholar] [CrossRef] [Green Version]
- Colpitts, S.L.; Dalton, N.M.; Scott, P. IL-7 receptor expression provides the potential for long-term survival of both CD62Lhigh central memory T cells and Th1 effector cells during Leishmania major infection. J. Immunol. 2009, 182, 5702–5711. [Google Scholar] [CrossRef] [Green Version]
- Lees, J.R.; Farber, D.L. Generation, persistence and plasticity of CD4 T-cell memories. Immunology 2010, 130, 463–470. [Google Scholar] [CrossRef]
- Kaech, S.M.; Wherry, E.J. Heterogeneity and cell-fate decisions in effector and memory CD8+ T cell differentiation during viral infection. Immunity 2007, 27, 393–405. [Google Scholar] [CrossRef] [Green Version]
- Gradoni, L. Canine Leishmania vaccines: Still a long way to go. Vet. Parasitol. 2015, 208, 94–100. [Google Scholar] [CrossRef]
- Okwor, I.; Uzonna, J. Vaccines and vaccination strategies against human cutaneous leishmaniasis. Hum. Vaccines 2009, 5, 291–301. [Google Scholar] [CrossRef] [Green Version]
- Soong, L.; Henard, C.A.; Melby, P.C. Immunopathogenesis of non-healing American cutaneous leishmaniasis and progressive visceral leishmaniasis. Semin. Immunopathol. 2012, 34, 735–751. [Google Scholar] [CrossRef] [Green Version]
- Bacellar, O.; Faria, D.; Nascimento, M.; Cardoso, T.M.; Gollob, K.J.; Dutra, W.O.; Scott, P.; Carvalho, E.M. Interleukin 17 production among patients with American cutaneous leishmaniasis. J. Infect. Dis. 2009, 200, 75–78. [Google Scholar] [CrossRef] [Green Version]
- Nylen, S.; Gautam, S. Immunological perspectives of leishmaniasis. J. Glob. Infect. Dis. 2010, 2, 135–146. [Google Scholar] [CrossRef]
- Silveira, F.T.; Lainson, R.; De Castro Gomes, C.M.; Laurenti, M.D.; Corbett, C.E. Immunopathogenic competences of Leishmania (V.) braziliensis and L. (L.) amazonensis in American cutaneous leishmaniasis. Parasite Immunol. 2009, 31, 423–431. [Google Scholar] [CrossRef]
- Kamir, D.; Zierow, S.; Leng, L.; Cho, Y.; Diaz, Y.; Griffith, J.; McDonald, C.; Merk, M.; Mitchell, R.A.; Trent, J.; et al. A Leishmania ortholog of macrophage migration inhibitory factor modulates host macrophage responses. J. Immunol. 2008, 180, 8250–8261. [Google Scholar] [CrossRef] [Green Version]
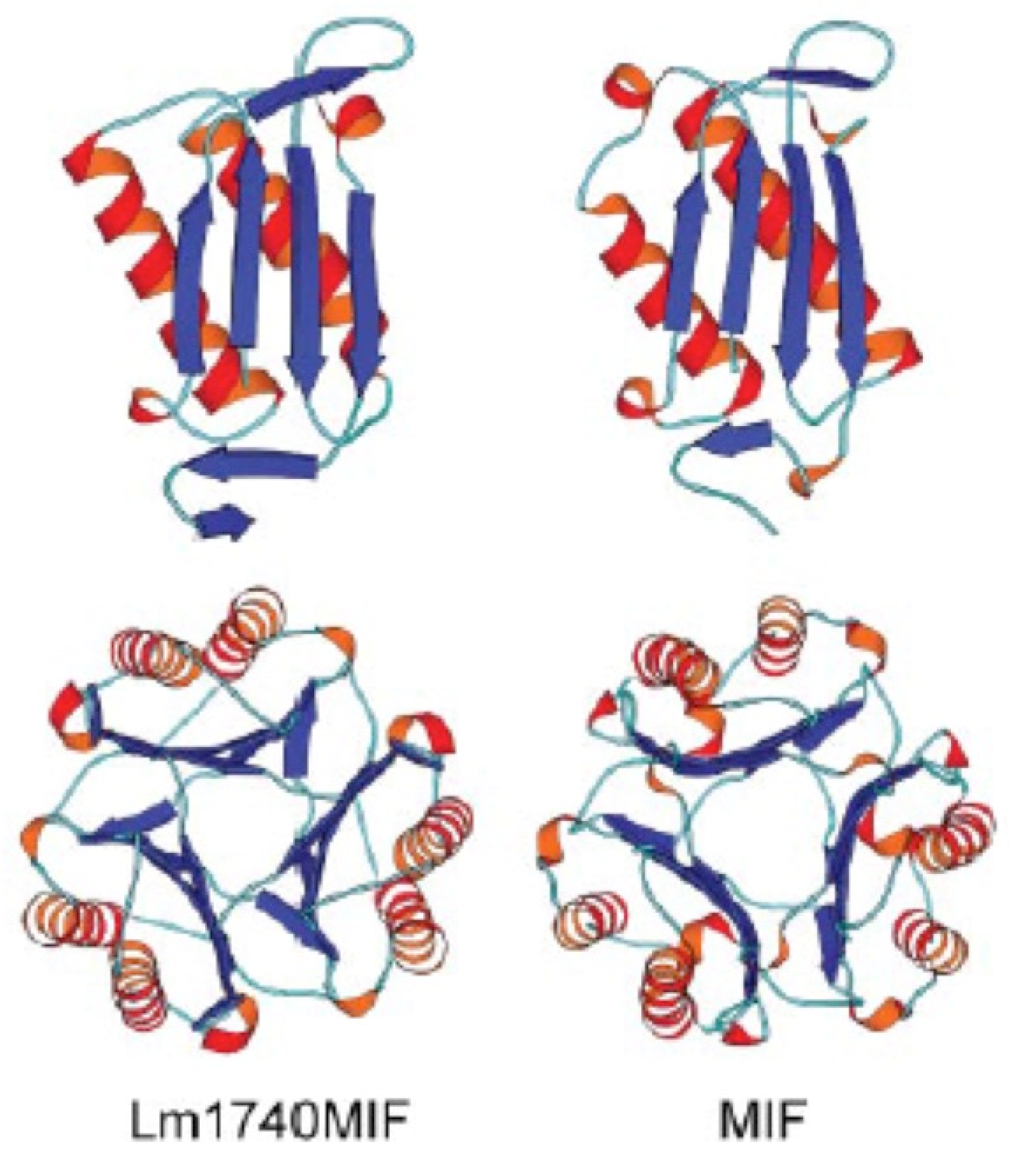
- Richardson, J.M.; Morrison, L.S.; Bland, N.D.; Bruce, S.; Coombs, G.H.; Mottram, J.C.; Walkinshaw, M.D. Structures of Leishmania major orthologues of macrophage migration inhibitory factor. Biochem. Biophys. Res. Commun. 2009, 380, 442–448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rich, A.R.; Lewis, M.R. The Nature of Allergy in Tuberculosis as Revealed by Tissue Culture Studies; Bulletin of the Johns Hopkins Hospital: Baltimore, MD, USA, 1932; pp. 115–132. [Google Scholar]
- David, J.R. Delayed hypersensitivity in vitro: Its mediation by cell-free substances formed by lymphoid cell-antigen interaction. Proc. Natl. Acad. Sci. USA 1966, 56, 72–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernhagen, J.; Calandra, T.; Mitchell, R.A.; Martin, S.B.; Tracey, K.J.; Voelter, W.; Manogue, K.R.; Cerami, A.; Bucala, R. MIF is a pituitary-derived cytokine that potentiates lethal endotoxaemia. Nature 1993, 365, 756–759. [Google Scholar] [CrossRef] [PubMed]
- Bernhagen, J.; Mitchell, R.A.; Calandra, T.; Voelter, W.; Cerami, A.; Bucala, R. Purification, bioactivity, and secondary structure analysis of mouse and human macrophage migration inhibitory factor (MIF). Biochemistry 1994, 33, 14144–14155. [Google Scholar] [CrossRef]
- Suzuki, M.; Sugimoto, H.; Nakagawa, A.; Tanaka, I.; Nishihira, J.; Sakai, M. Crystal structure of the macrophage migration inhibitory factor from rat liver. Nat. Struct. Biol. 1996, 3, 259–266. [Google Scholar] [CrossRef]
- Sun, H.W.; Bernhagen, J.; Bucala, R.; Lolis, E. Crystal structure at 2.6-A resolution of human macrophage migration inhibitory factor. Proc. Natl. Acad. Sci. USA 1996, 93, 5191–5196. [Google Scholar] [CrossRef] [Green Version]
- Baugh, J.A.; Chitnis, S.; Donnelly, S.C.; Monteiro, J.; Lin, X.; Plant, B.J.; Wolfe, F.; Gregersen, P.K.; Bucala, R. A functional promoter polymorphism in the macrophage migration inhibitory factor (MIF) gene associated with disease severity in rheumatoid arthritis. Genes Immun. 2002, 3, 170–176. [Google Scholar] [CrossRef] [Green Version]
- Merk, M.; Baugh, J.; Zierow, S.; Leng, L.; Pal, U.; Lee, S.J.; Ebert, A.D.; Mizue, Y.; Trent, J.O.; Mitchell, R.; et al. The Golgi-associated protein p115 mediates the secretion of macrophage migration inhibitory factor. J. Immunol. 2009, 182, 6896–6906. [Google Scholar] [CrossRef]
- Leng, L.; Metz, C.N.; Fang, Y.; Xu, J.; Donnelly, S.; Baugh, J.; Delohery, T.; Chen, Y.; Mitchell, R.A.; Bucala, R. MIF signal transduction initiated by binding to CD74. J. Exp. Med. 2003, 197, 1467–1476. [Google Scholar] [CrossRef] [Green Version]
- Shi, X.; Leng, L.; Wang, T.; Wang, W.; Du, X.; Li, J.; McDonald, C.; Chen, Z.; Murphy, J.W.; Lolis, E.; et al. CD44 is the signaling component of the macrophage migration inhibitory factor-CD74 receptor complex. Immunity 2006, 25, 595–606. [Google Scholar] [CrossRef] [Green Version]
- Bucala, R. MIF, MIF alleles, and prospects for therapeutic intervention in autoimmunity. J. Clin. Immunol. 2013, 33 (Suppl. 1), S72–S78. [Google Scholar] [CrossRef] [Green Version]
- Lue, H.; Thiele, M.; Franz, J.; Dahl, E.; Speckgens, S.; Leng, L.; Fingerle-Rowson, G.; Bucala, R.; Luscher, B.; Bernhagen, J. Macrophage migration inhibitory factor (MIF) promotes cell survival by activation of the Akt pathway and role for CSN5/JAB1 in the control of autocrine MIF activity. Oncogene 2007, 26, 5046–5059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernhagen, J.; Krohn, R.; Lue, H.; Gregory, J.L.; Zernecke, A.; Koenen, R.R.; Dewor, M.; Georgiev, I.; Schober, A.; Leng, L.; et al. MIF is a noncognate ligand of CXC chemokine receptors in inflammatory and atherogenic cell recruitment. Nat. Med. 2007, 13, 587–596. [Google Scholar] [CrossRef] [PubMed]
- Calandra, T.; Roger, T. Macrophage migration inhibitory factor: A regulator of innate immunity. Nat. Rev. Immunol. 2003, 3, 791–800. [Google Scholar] [CrossRef]
- Calandra, T.; Bernhagen, J.; Metz, C.N.; Spiegel, L.A.; Bacher, M.; Donnelly, T.; Cerami, A.; Bucala, R. MIF as a glucocorticoid-induced modulator of cytokine production. Nature 1995, 377, 68–71. [Google Scholar] [CrossRef]
- Bucala, R. MIF: Most Ineteresting Factor; World Scientific Publishing Company: Singapore, 2007. [Google Scholar]
- Roger, T.; David, J.; Glauser, M.P.; Calandra, T. MIF regulates innate immune responses through modulation of Toll-like receptor 4. Nature 2001, 414, 920–924. [Google Scholar] [CrossRef]
- Daun, J.M.; Cannon, J.G. Macrophage migration inhibitory factor antagonizes hydrocortisone-induced increases in cytosolic IkappaBalpha. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2000, 279, R1043–R1049. [Google Scholar] [CrossRef]
- Roger, T.; Chanson, A.L.; Knaup-Reymond, M.; Calandra, T. Macrophage migration inhibitory factor promotes innate immune responses by suppressing glucocorticoid-induced expression of mitogen-activated protein kinase phosphatase-1. Eur. J. Immunol. 2005, 35, 3405–3413. [Google Scholar] [CrossRef]
- Mitchell, R.A.; Liao, H.; Chesney, J.; Fingerle-Rowson, G.; Baugh, J.; David, J.; Bucala, R. Macrophage migration inhibitory factor (MIF) sustains macrophage proinflammatory function by inhibiting p53: Regulatory role in the innate immune response. Proc. Natl. Acad. Sci. USA 2002, 99, 345–350. [Google Scholar] [CrossRef] [Green Version]
- Xaus, J.; Comalada, M.; Valledor, A.F.; Lloberas, J.; Lopez-Soriano, F.; Argiles, J.M.; Bogdan, C.; Celada, A. LPS induces apoptosis in macrophages mostly through the autocrine production of TNF-alpha. Blood 2000, 95, 3823–3831. [Google Scholar] [CrossRef]
- Amin, M.A.; Haas, C.S.; Zhu, K.; Mansfield, P.J.; Kim, M.J.; Lackowski, N.P.; Koch, A.E. Migration inhibitory factor up-regulates vascular cell adhesion molecule-1 and intercellular adhesion molecule-1 via Src, PI3 kinase, and NFkappaB. Blood 2006, 107, 2252–2261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, I.; Bucala, R. The immunobiology of MIF: Function, genetics and prospects for precision medicine. Nat. Rev. Rheumatol. 2019, 15, 427–437. [Google Scholar] [CrossRef] [PubMed]
- Mizue, Y.; Ghani, S.; Leng, L.; McDonald, C.; Kong, P.; Baugh, J.; Lane, S.J.; Craft, J.; Nishihira, J.; Donnelly, S.C.; et al. Role for macrophage migration inhibitory factor in asthma. Proc. Natl. Acad. Sci. USA 2005, 102, 14410–14415. [Google Scholar] [CrossRef] [Green Version]
- Niino, M.; Ogata, A.; Kikuchi, S.; Tashiro, K.; Nishihira, J. Macrophage migration inhibitory factor in the cerebrospinal fluid of patients with conventional and optic-spinal forms of multiple sclerosis and neuro-Behcet’s disease. J. Neurol. Sci. 2000, 179, 127–131. [Google Scholar] [CrossRef]
- Plant, B.J.; Gallagher, C.G.; Bucala, R.; Baugh, J.A.; Chappell, S.; Morgan, L.; O’Connor, C.M.; Morgan, K.; Donnelly, S.C. Cystic fibrosis, disease severity, and a macrophage migration inhibitory factor polymorphism. Am. J. Respir. Crit. Care Med. 2005, 172, 1412–1415. [Google Scholar] [CrossRef]
- Bozza, F.A.; Gomes, R.N.; Japiassu, A.M.; Soares, M.; Castro-Faria-Neto, H.C.; Bozza, P.T.; Bozza, M.T. Macrophage migration inhibitory factor levels correlate with fatal outcome in sepsis. Shock 2004, 22, 309–313. [Google Scholar] [CrossRef]
- Calandra, T.; Echtenacher, B.; Roy, D.L.; Pugin, J.; Metz, C.N.; Hultner, L.; Heumann, D.; Mannel, D.; Bucala, R.; Glauser, M.P. Protection from septic shock by neutralization of macrophage migration inhibitory factor. Nat. Med. 2000, 6, 164–170. [Google Scholar] [CrossRef]
- Donnelly, S.C.; Haslett, C.; Reid, P.T.; Grant, I.S.; Wallace, W.A.; Metz, C.N.; Bruce, L.J.; Bucala, R. Regulatory role for macrophage migration inhibitory factor in acute respiratory distress syndrome. Nat. Med. 1997, 3, 320–323. [Google Scholar] [CrossRef]
- Murakami, H.; Akbar, S.M.; Matsui, H.; Horiike, N.; Onji, M. Macrophage migration inhibitory factor activates antigen-presenting dendritic cells and induces inflammatory cytokines in ulcerative colitis. Clin. Exp. Immunol. 2002, 128, 504–510. [Google Scholar] [CrossRef]
- Meyer-Siegler, K.L.; Vera, P.L.; Iczkowski, K.A.; Bifulco, C.; Lee, A.; Gregersen, P.K.; Leng, L.; Bucala, R. Macrophage migration inhibitory factor (MIF) gene polymorphisms are associated with increased prostate cancer incidence. Genes Immun. 2007, 8, 646–652. [Google Scholar] [CrossRef]
- Mitchell, R.A. Mechanisms and effectors of MIF-dependent promotion of tumourigenesis. Cell Signal. 2004, 16, 13–19. [Google Scholar] [CrossRef]
- Bucala, R.; Donnelly, S.C. Macrophage migration inhibitory factor: A probable link between inflammation and cancer. Immunity 2007, 26, 281–285. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; He, P.; Wang, J.; Schetter, A.; Tang, W.; Funamizu, N.; Yanaga, K.; Uwagawa, T.; Satoskar, A.R.; Gaedcke, J.; et al. A Novel MIF Signaling Pathway Drives the Malignant Character of Pancreatic Cancer by Targeting NR3C2. Cancer Res. 2016, 76, 3838–3850. [Google Scholar] [CrossRef] [Green Version]
- Jung, H.; Seong, H.A.; Ha, H. Direct interaction between NM23-H1 and macrophage migration inhibitory factor (MIF) is critical for alleviation of MIF-mediated suppression of p53 activity. J. Biol. Chem. 2008, 283, 32669–32679. [Google Scholar] [CrossRef] [Green Version]
- Das, R.; Koo, M.S.; Kim, B.H.; Jacob, S.T.; Subbian, S.; Yao, J.; Leng, L.; Levy, R.; Murchison, C.; Burman, W.J.; et al. Macrophage migration inhibitory factor (MIF) is a critical mediator of the innate immune response to Mycobacterium tuberculosis. Proc. Natl. Acad. Sci. USA 2013, 110, E2997–E3006. [Google Scholar] [CrossRef] [Green Version]
- Oddo, M.; Calandra, T.; Bucala, R.; Meylan, P.R. Macrophage migration inhibitory factor reduces the growth of virulent Mycobacterium tuberculosis in human macrophages. Infect. Immun. 2005, 73, 3783–3786. [Google Scholar] [CrossRef] [Green Version]
- Flores, M.; Saavedra, R.; Bautista, R.; Viedma, R.; Tenorio, E.P.; Leng, L.; Sanchez, Y.; Juarez, I.; Satoskar, A.A.; Shenoy, A.S.; et al. Macrophage migration inhibitory factor (MIF) is critical for the host resistance against Toxoplasma gondii. FASEB J. 2008, 22, 3661–3671. [Google Scholar] [CrossRef] [Green Version]
- Koebernick, H.; Grode, L.; David, J.R.; Rohde, W.; Rolph, M.S.; Mittrucker, H.W.; Kaufmann, S.H. Macrophage migration inhibitory factor (MIF) plays a pivotal role in immunity against Salmonella typhimurium. Proc. Natl. Acad. Sci. USA 2002, 99, 13681–13686. [Google Scholar] [CrossRef] [Green Version]
- Reyes, J.L.; Terrazas, L.I.; Espinoza, B.; Cruz-Robles, D.; Soto, V.; Rivera-Montoya, I.; Gomez-Garcia, L.; Snider, H.; Satoskar, A.R.; Rodriguez-Sosa, M. Macrophage migration inhibitory factor contributes to host defense against acute Trypanosoma cruzi infection. Infect. Immun. 2006, 74, 3170–3179. [Google Scholar] [CrossRef] [Green Version]
- Terrazas, C.A.; Juarez, I.; Terrazas, L.I.; Saavedra, R.; Calleja, E.A.; Rodriguez-Sosa, M. Toxoplasma gondii: Impaired maturation and pro-inflammatory response of dendritic cells in MIF-deficient mice favors susceptibility to infection. Exp. Parasitol. 2010, 126, 348–358. [Google Scholar] [CrossRef]
- Arjona, A.; Foellmer, H.G.; Town, T.; Leng, L.; McDonald, C.; Wang, T.; Wong, S.J.; Montgomery, R.R.; Fikrig, E.; Bucala, R. Abrogation of macrophage migration inhibitory factor decreases West Nile virus lethality by limiting viral neuroinvasion. J. Clin. Investig. 2007, 117, 3059–3066. [Google Scholar] [CrossRef] [PubMed]
- Assuncao-Miranda, I.; Amaral, F.A.; Bozza, F.A.; Fagundes, C.T.; Sousa, L.P.; Souza, D.G.; Pacheco, P.; Barbosa-Lima, G.; Gomes, R.N.; Bozza, P.T.; et al. Contribution of macrophage migration inhibitory factor to the pathogenesis of dengue virus infection. FASEB J. 2010, 24, 218–228. [Google Scholar] [CrossRef] [PubMed]
- Bozza, M.; Satoskar, A.R.; Lin, G.; Lu, B.; Humbles, A.A.; Gerard, C.; David, J.R. Targeted disruption of migration inhibitory factor gene reveals its critical role in sepsis. J. Exp. Med. 1999, 189, 341–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, B.L.; Zhu, S.L.; Huang, X.R.; Ma, J.; Xia, H.H.; Bucala, R.; Wong, B.C.; Lan, H.Y. Essential role for macrophage migration inhibitory factor in gastritis induced by Helicobacter pylori. Am. J. Pathol. 2009, 174, 1319–1328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.C.; Lei, H.Y.; Liu, C.C.; Shiesh, S.C.; Chen, S.H.; Liu, H.S.; Lin, Y.S.; Wang, S.T.; Shyu, H.W.; Yeh, T.M. Correlation of serum levels of macrophage migration inhibitory factor with disease severity and clinical outcome in dengue patients. Am. J. Trop. Med. Hyg. 2006, 74, 142–147. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.Y.; Nanji, A.A.; Luk, J.M.; Huang, X.R.; Lo, C.M.; Chen, Y.X.; Yuen, S.T.; Lan, H.Y.; Lau, G.K. Macrophage migration inhibitory factor expression correlates with inflammatory changes in human chronic hepatitis B infection. Liver Int. 2005, 25, 571–579. [Google Scholar] [CrossRef]
- Doernberg, S.; Schaaf, B.; Dalhoff, K.; Leng, L.; Beitin, A.; Quagliarello, V.; Bucala, R. Association of macrophage migration inhibitory factor (MIF) polymorphisms with risk of meningitis from Streptococcus pneumoniae. Cytokine 2011, 53, 292–294. [Google Scholar] [CrossRef] [Green Version]
- Xia, H.H.; Lam, S.K.; Huang, X.R.; Wong, W.M.; Leung, S.Y.; Yuen, S.T.; Lan, H.Y.; Wong, B.C. Helicobacter pylori infection is associated with increased expression of macrophage migratory inhibitory factor—By epithelial cells, T cells, and macrophages—In gastric mucosa. J. Infect. Dis. 2004, 190, 293–302. [Google Scholar] [CrossRef] [Green Version]
- Malu, D.T.; Belanger, B.; Desautels, F.; Kelendji, K.; Dalko, E.; Sanchez-Dardon, J.; Leng, L.; Bucala, R.; Satoskar, A.R.; Scorza, T. Macrophage migration inhibitory factor: A downregulator of early T cell-dependent IFN-gamma responses in Plasmodium chabaudi adami (556 KA)-infected mice. J. Immunol. 2011, 186, 6271–6279. [Google Scholar] [CrossRef]
- Martiney, J.A.; Sherry, B.; Metz, C.N.; Espinoza, M.; Ferrer, A.S.; Calandra, T.; Broxmeyer, H.E.; Bucala, R. Macrophage migration inhibitory factor release by macrophages after ingestion of Plasmodium chabaudi-infected erythrocytes: Possible role in the pathogenesis of malarial anemia. Infect. Immun. 2000, 68, 2259–2267. [Google Scholar] [CrossRef] [Green Version]
- McDevitt, M.A.; Xie, J.; Shanmugasundaram, G.; Griffith, J.; Liu, A.; McDonald, C.; Thuma, P.; Gordeuk, V.R.; Metz, C.N.; Mitchell, R.; et al. A critical role for the host mediator macrophage migration inhibitory factor in the pathogenesis of malarial anemia. J. Exp. Med. 2006, 203, 1185–1196. [Google Scholar] [CrossRef] [PubMed]
- Chaisavaneeyakorn, S.; Moore, J.M.; Othoro, C.; Otieno, J.; Chaiyaroj, S.C.; Shi, Y.P.; Nahlen, B.L.; Lal, A.A.; Udhayakumar, V. Immunity to placental malaria. IV. Placental malaria is associated with up-regulation of macrophage migration inhibitory factor in intervillous blood. J. Infect. Dis. 2002, 186, 1371–1375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jain, V.; McClintock, S.; Nagpal, A.C.; Dash, A.P.; Stiles, J.K.; Udhayakumar, V.; Singh, N.; Lucchi, N.W. Macrophage migration inhibitory factor is associated with mortality in cerebral malaria patients in India. BMC Res. Notes 2009, 2, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Awandare, G.A.; Martinson, J.J.; Were, T.; Ouma, C.; Davenport, G.C.; Ong’echa, J.M.; Wang, W.; Leng, L.; Ferrell, R.E.; Bucala, R.; et al. MIF (macrophage migration inhibitory factor) promoter polymorphisms and susceptibility to severe malarial anemia. J. Infect. Dis. 2009, 200, 629–637. [Google Scholar] [CrossRef] [Green Version]
- Awandare, G.A.; Ouma, C.; Keller, C.C.; Were, T.; Otieno, R.; Ouma, Y.; Davenport, G.C.; Hittner, J.B.; Ong’echa, J.M.; Ferrell, R.; et al. A macrophage migration inhibitory factor promoter polymorphism is associated with high-density parasitemia in children with malaria. Genes Immun. 2006, 7, 568–575. [Google Scholar] [CrossRef] [Green Version]
- Jha, A.N.; Sundaravadivel, P.; Pati, S.S.; Patra, P.K.; Thangaraj, K. Variations in ncRNA gene LOC284889 and MIF-794CATT repeats are associated with malaria susceptibility in Indian populations. Malar. J. 2013, 12, 345. [Google Scholar] [CrossRef] [Green Version]
- Subramanya, H.S.; Roper, D.I.; Dauter, Z.; Dodson, E.J.; Davies, G.J.; Wilson, K.S.; Wigley, D.B. Enzymatic ketonization of 2-hydroxymuconate: Specificity and mechanism investigated by the crystal structures of two isomerases. Biochemistry 1996, 35, 792–802. [Google Scholar] [CrossRef]
- Juttner, S.; Bernhagen, J.; Metz, C.N.; Rollinghoff, M.; Bucala, R.; Gessner, A. Migration inhibitory factor induces killing of Leishmania major by macrophages: Dependence on reactive nitrogen intermediates and endogenous TNF-alpha. J. Immunol. 1998, 161, 2383–2390. [Google Scholar]
- Alonso, D.; Serrano, E.; Bermejo, F.J.; Corral, R.S. HIF-1alpha-regulated MIF activation and Nox2-dependent ROS generation promote Leishmania amazonensis killing by macrophages under hypoxia. Cell. Immunol. 2019, 335, 15–21. [Google Scholar] [CrossRef]
- Satoskar, A.R.; Bozza, M.; Rodriguez Sosa, M.; Lin, G.; David, J.R. Migration-inhibitory factor gene-deficient mice are susceptible to cutaneous Leishmania major infection. Infect. Immun. 2001, 69, 906–911. [Google Scholar] [CrossRef] [Green Version]
- Xu, D.; McSorley, S.J.; Tetley, L.; Chatfield, S.; Dougan, G.; Chan, W.L.; Satoskar, A.; David, J.R.; Liew, F.Y. Protective effect on Leishmania major infection of migration inhibitory factor, TNF-alpha, and IFN-gamma administered orally via attenuated Salmonella typhimurium. J. Immunol. 1998, 160, 1285–1289. [Google Scholar] [PubMed]
- Kar, S.; Metz, C.; McMahon-Pratt, D. CD4+ T cells play a dominant role in protection against New World leishmaniasis induced by vaccination with the P-4 amastigote antigen. Infect. Immun. 2005, 73, 3823–3827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiser, W.Y.; Pozzi, L.M.; David, J.R. Human recombinant migration inhibitory factor activates human macrophages to kill Leishmania donovani. J. Immunol. 1991, 147, 2006–2011. [Google Scholar] [PubMed]
- Wu, J.; Cunha, F.Q.; Liew, F.Y.; Weiser, W.Y. IL-10 inhibits the synthesis of migration inhibitory factor and migration inhibitory factor-mediated macrophage activation. J. Immunol. 1993, 151, 4325–4332. [Google Scholar] [PubMed]
- Mishra, A.; Sundaravadivel, P.; Tripathi, S.K.; Jha, R.K.; Badrukhiya, J.; Basak, N.; Anerao, I.; Sharma, A.; Idowu, A.E.; Mishra, A.; et al. Variations in macrophage migration inhibitory factor gene are not associated with visceral leishmaniasis in India. J. Infect. Public Health 2019, 12, 380–387. [Google Scholar] [CrossRef] [PubMed]
- de Jesus Fernandes Covas, C.; Cardoso, C.C.; Gomes-Silva, A.; Santos Oliveira, J.R.; Da-Cruz, A.M.; Moraes, M.O. Candidate gene case-control and functional study shows macrophage inhibitory factor (MIF) polymorphism is associated with cutaneous leishmaniasis. Cytokine 2013, 61, 168–172. [Google Scholar] [CrossRef] [Green Version]
- Santos-Oliveira, J.R.; Regis, E.G.; Leal, C.R.; Cunha, R.V.; Bozza, P.T.; Da-Cruz, A.M. Evidence that lipopolisaccharide may contribute to the cytokine storm and cellular activation in patients with visceral leishmaniasis. PLoS Negl. Trop. Dis. 2011, 5, e1198. [Google Scholar] [CrossRef] [Green Version]
- Sparkes, A.; De Baetselier, P.; Roelants, K.; De Trez, C.; Magez, S.; Van Ginderachter, J.A.; Raes, G.; Bucala, R.; Stijlemans, B. The non-mammalian MIF superfamily. Immunobiology 2017, 222, 473–482. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, S.; Jiang, N.; Farr, L.; Ngobeni, R.; Moonah, S. Parasite-Produced MIF Cytokine: Role in Immune Evasion, Invasion, and Pathogenesis. Front. Immunol. 2019, 10, 1995. [Google Scholar] [CrossRef]
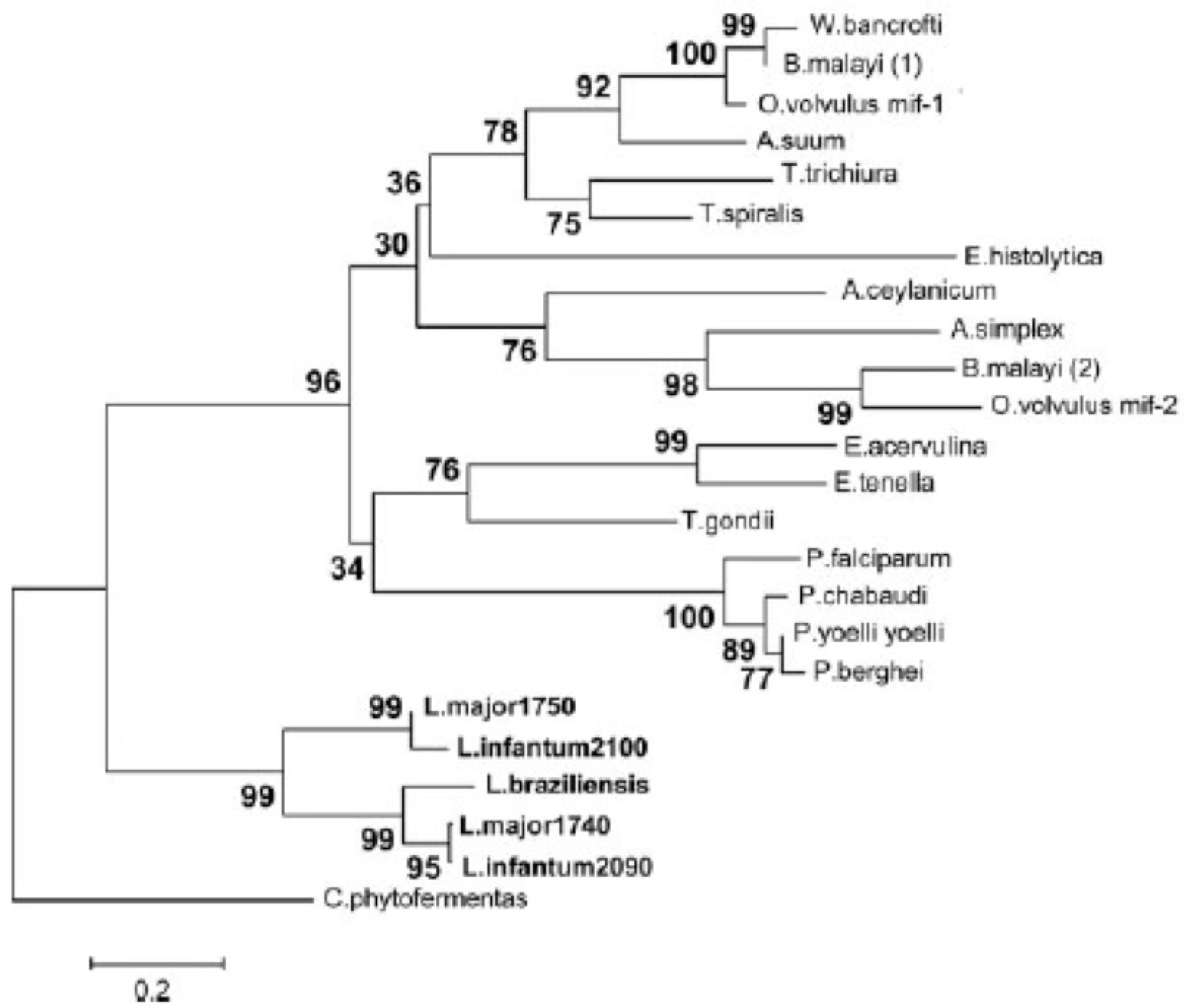
- Vermeire, J.J.; Cho, Y.; Lolis, E.; Bucala, R.; Cappello, M. Orthologs of macrophage migration inhibitory factor from parasitic nematodes. Trends Parasitol. 2008, 24, 355–363. [Google Scholar] [CrossRef] [Green Version]
- Stiles, J.K.; Hicock, P.I.; Shah, P.H.; Meade, J.C. Genomic organization, transcription, splicing and gene regulation in Leishmania. Ann. Trop. Med. Parasitol. 1999, 93, 781–807. [Google Scholar] [CrossRef] [PubMed]
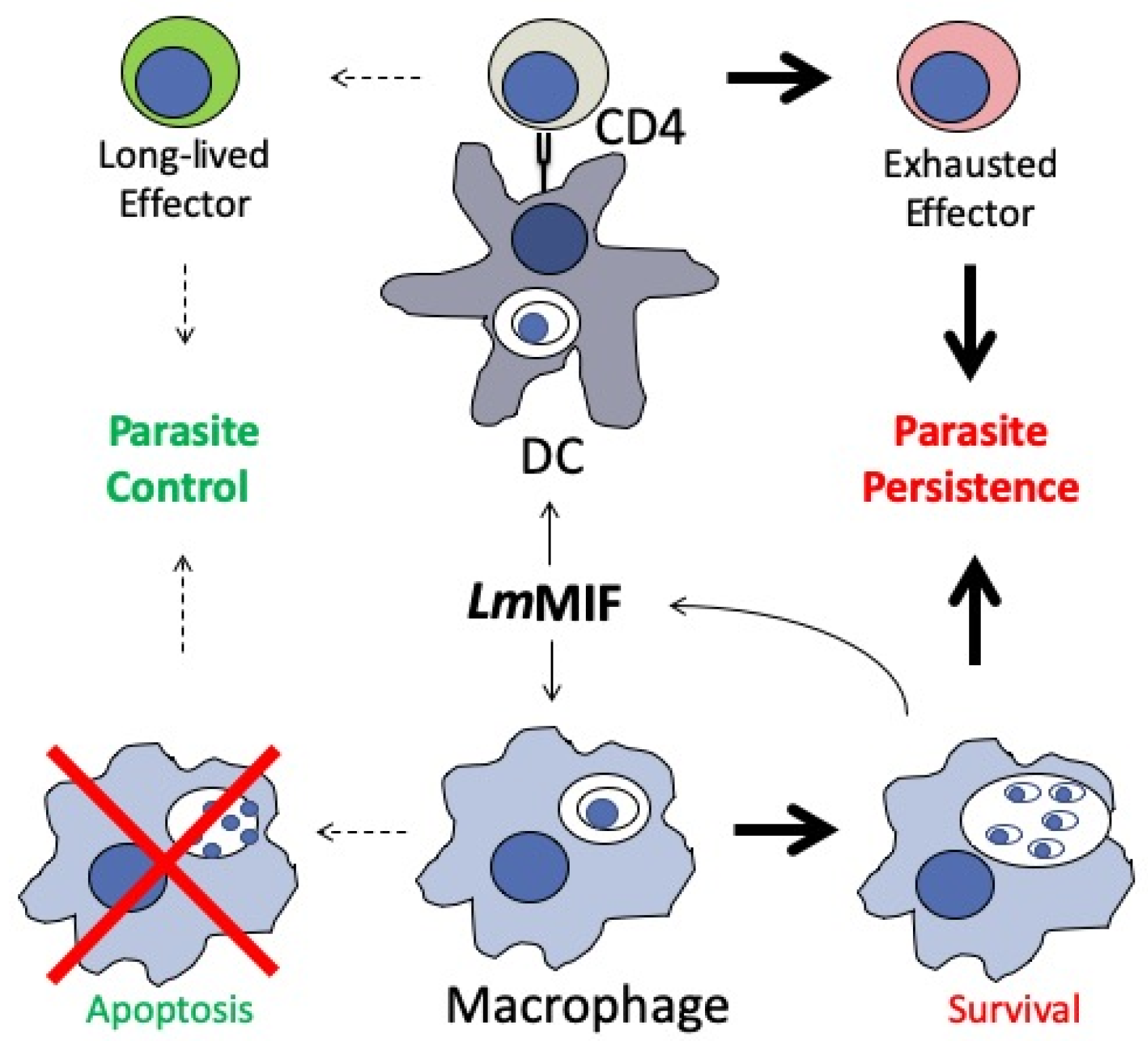
- Holowka, T.; Castilho, T.M.; Garcia, A.B.; Sun, T.; McMahon-Pratt, D.; Bucala, R. Leishmania-encoded orthologs of macrophage migration inhibitory factor regulate host immunity to promote parasite persistence. FASEB J. 2016, 30, 2249–2265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, T.; Holowka, T.; Song, Y.; Zierow, S.; Leng, L.; Chen, Y.; Xiong, H.; Griffith, J.; Nouraie, M.; Thuma, P.E.; et al. A Plasmodium-encoded cytokine suppresses T-cell immunity during malaria. Proc. Natl. Acad. Sci. USA 2012, 109, E2117–E2126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Younis, A.E.; Soblik, H.; Ajonina-Ekoti, I.; Erttmann, K.D.; Luersen, K.; Liebau, E.; Brattig, N.W. Characterization of a secreted macrophage migration inhibitory factor homologue of the parasitic nematode Strongyloides acting at the parasite-host cell interface. Microbes Infect. 2012, 14, 279–289. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.; Hoti, S.L.; Meena, R.L.; Vasuki, V.; Sankari, T.; Kaliraj, P. Molecular and functional characterization of macrophage migration inhibitory factor (MIF) homolog of human from lymphatic filarial parasite Wuchereria bancrofti. Parasitol. Res. 2012, 111, 2035–2047. [Google Scholar] [CrossRef]
- Qu, G.; Fetterer, R.; Jenkins, M.; Leng, L.; Shen, Z.; Murphy, C.; Han, W.; Bucala, R.; Tuo, W. Characterization of Neospora caninum macrophage migration inhibitory factor. Exp. Parasitol. 2013, 135, 246–256. [Google Scholar] [CrossRef] [Green Version]
- Falcone, F.H.; Loke, P.; Zang, X.; MacDonald, A.S.; Maizels, R.M.; Allen, J.E. A Brugia malayi homolog of macrophage migration inhibitory factor reveals an important link between macrophages and eosinophil recruitment during nematode infection. J. Immunol. 2001, 167, 5348–5354. [Google Scholar] [CrossRef] [Green Version]
- Prieto-Lafuente, L.; Gregory, W.F.; Allen, J.E.; Maizels, R.M. MIF homologues from a filarial nematode parasite synergize with IL-4 to induce alternative activation of host macrophages. J. Leukoc. Biol. 2009, 85, 844–854. [Google Scholar] [CrossRef]
- Cho, M.K.; Lee, C.H.; Yu, H.S. Amelioration of intestinal colitis by macrophage migration inhibitory factor isolated from intestinal parasites through toll-like receptor 2. Parasite Immunol. 2011, 33, 265–275. [Google Scholar] [CrossRef] [PubMed]
- Cho, M.K.; Park, M.K.; Kang, S.A.; Park, S.K.; Lyu, J.H.; Kim, D.H.; Park, H.K.; Yu, H.S. TLR2-dependent amelioration of allergic airway inflammation by parasitic nematode type II MIF in mice. Parasite Immunol. 2015, 37, 180–191. [Google Scholar] [CrossRef]
- Park, S.K.; Cho, M.K.; Park, H.K.; Lee, K.H.; Lee, S.J.; Choi, S.H.; Ock, M.S.; Jeong, H.J.; Lee, M.H.; Yu, H.S. Macrophage migration inhibitory factor homologs of Anisakis simplex suppress Th2 response in allergic airway inflammation model via CD4+CD25+Foxp3+ T cell recruitment. J. Immunol. 2009, 182, 6907–6914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fox, R.J.; Coffey, C.S.; Conwit, R.; Cudkowicz, M.E.; Gleason, T.; Goodman, A.; Klawiter, E.C.; Matsuda, K.; McGovern, M.; Naismith, R.T.; et al. Phase 2 Trial of Ibudilast in Progressive Multiple Sclerosis. N. Engl. J. Med. 2018, 379, 846–855. [Google Scholar] [CrossRef] [PubMed]
- Benedek, G.; Meza-Romero, R.; Jordan, K.; Zhang, Y.; Nguyen, H.; Kent, G.; Li, J.; Siu, E.; Frazer, J.; Piecychna, M.; et al. MIF and D-DT are potential disease severity modifiers in male MS subjects. Proc. Natl. Acad. Sci. USA 2017, 114, E8421–E8429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cournia, Z.; Leng, L.; Gandavadi, S.; Du, X.; Bucala, R.; Jorgensen, W.L. Discovery of human macrophage migration inhibitory factor (MIF)-CD74 antagonists via virtual screening. J. Med. Chem. 2009, 52, 416–424. [Google Scholar] [CrossRef] [Green Version]
- Lubetsky, J.B.; Dios, A.; Han, J.; Aljabari, B.; Ruzsicska, B.; Mitchell, R.; Lolis, E.; Al-Abed, Y. The tautomerase active site of macrophage migration inhibitory factor is a potential target for discovery of novel anti-inflammatory agents. J. Biol. Chem. 2002, 277, 24976–24982. [Google Scholar] [CrossRef] [Green Version]
- Cho, Y.; Jones, B.F.; Vermeire, J.J.; Leng, L.; DiFedele, L.; Harrison, L.M.; Xiong, H.; Kwong, Y.K.; Chen, Y.; Bucala, R.; et al. Structural and functional characterization of a secreted hookworm Macrophage Migration Inhibitory Factor (MIF) that interacts with the human MIF receptor CD74. J. Biol. Chem. 2007, 282, 23447–23456. [Google Scholar] [CrossRef] [Green Version]
- Sommerville, C.; Richardson, J.M.; Williams, R.A.; Mottram, J.C.; Roberts, C.W.; Alexander, J.; Henriquez, F.L. Biochemical and immunological characterization of Toxoplasma gondii macrophage migration inhibitory factor. J. Biol. Chem. 2013, 288, 12733–12741. [Google Scholar] [CrossRef] [Green Version]
- Qu, G.; Fetterer, R.; Leng, L.; Du, X.; Zarlenga, D.; Shen, Z.; Han, W.; Bucala, R.; Tuo, W. Ostertagia ostertagi macrophage migration inhibitory factor is present in all developmental stages and may cross-regulate host functions through interaction with the host receptor. Int. J. Parasitol. 2014, 44, 355–367. [Google Scholar] [CrossRef] [Green Version]
- Dahlgren, M.K.; Garcia, A.B.; Hare, A.A.; Tirado-Rives, J.; Leng, L.; Bucala, R.; Jorgensen, W.L. Virtual screening and optimization yield low-nanomolar inhibitors of the tautomerase activity of Plasmodium falciparum macrophage migration inhibitory factor. J. Med. Chem. 2012, 55, 10148–10159. [Google Scholar] [CrossRef] [Green Version]
- Pantouris, G.; Rajasekaran, D.; Garcia, A.B.; Ruiz, V.G.; Leng, L.; Jorgensen, W.L.; Bucala, R.; Lolis, E.J. Crystallographic and receptor binding characterization of Plasmodium falciparum macrophage migration inhibitory factor complexed to two potent inhibitors. J. Med. Chem. 2014, 57, 8652–8656. [Google Scholar] [CrossRef] [Green Version]
- Cho, Y.; Vermeire, J.J.; Merkel, J.S.; Leng, L.; Du, X.; Bucala, R.; Cappello, M.; Lolis, E. Drug repositioning and pharmacophore identification in the discovery of hookworm MIF inhibitors. Chem. Biol. 2011, 18, 1089–1101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, F.; Xu, L.; Yan, R.; Song, X.; Li, X. Evaluation of the immune response induced by DNA vaccines expressing MIF and MCD-1 genes of Trichinella spiralis in BALB/c mice. J. Helminthol. 2012, 86, 430–439. [Google Scholar] [CrossRef] [PubMed]
- Tang, F.; Xu, L.; Yan, R.; Song, X.; Li, X. A DNA vaccine co-expressing Trichinella spiralis MIF and MCD-1 with murine ubiquitin induces partial protective immunity in mice. J. Helminthol. 2013, 87, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Wen, H.; Cai, H.; Wu, M.; An, R.; Chu, D.; Yu, L.; Shen, J.; Chen, L.; Du, J. Protective Effect against Toxoplasmosis in BALB/c Mice Vaccinated with Toxoplasma gondii Macrophage Migration Inhibitory Factor. Front. Microbiol. 2019, 10, 813. [Google Scholar] [CrossRef]
- Baeza Garcia, A.; Siu, E.; Sun, T.; Exler, V.; Brito, L.; Hekele, A.; Otten, G.; Augustijn, K.; Janse, C.J.; Ulmer, J.B.; et al. Neutralization of the Plasmodium-encoded MIF ortholog confers protective immunity against malaria infection. Nat. Commun. 2018, 9, 2714. [Google Scholar] [CrossRef]
- Sundar, S.; Chakravarty, J. Leishmaniasis: An update of current pharmacotherapy. Expert Opin. Pharmacother. 2013, 14, 53–63. [Google Scholar] [CrossRef]
- Moafi, M.; Rezvan, H.; Sherkat, R.; Taleban, R. Leishmania Vaccines Entered in Clinical Trials: A Review of Literature. Int. J. Prev. Med. 2019, 10, 95. [Google Scholar] [CrossRef]



© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Holowka, T.; Bucala, R. Role of Host and Parasite MIF Cytokines during Leishmania Infection. Trop. Med. Infect. Dis. 2020, 5, 46. https://doi.org/10.3390/tropicalmed5010046
Holowka T, Bucala R. Role of Host and Parasite MIF Cytokines during Leishmania Infection. Tropical Medicine and Infectious Disease. 2020; 5(1):46. https://doi.org/10.3390/tropicalmed5010046
Chicago/Turabian StyleHolowka, Thomas, and Richard Bucala. 2020. "Role of Host and Parasite MIF Cytokines during Leishmania Infection" Tropical Medicine and Infectious Disease 5, no. 1: 46. https://doi.org/10.3390/tropicalmed5010046