The Contribution of Wastewater to the Transmission of Antimicrobial Resistance in the Environment: Implications of Mass Gathering Settings

 , ,
, ,  and
and
Abstract
:1. Introduction
1.1. The Current Status of AMR as a Major Global Health Challenge
1.2. WHO AMR Priority Pathogens List
1.3. The Main Drivers of AMR Transmission
1.4. The Environmental Reservoir of AMR from Water and Sewage
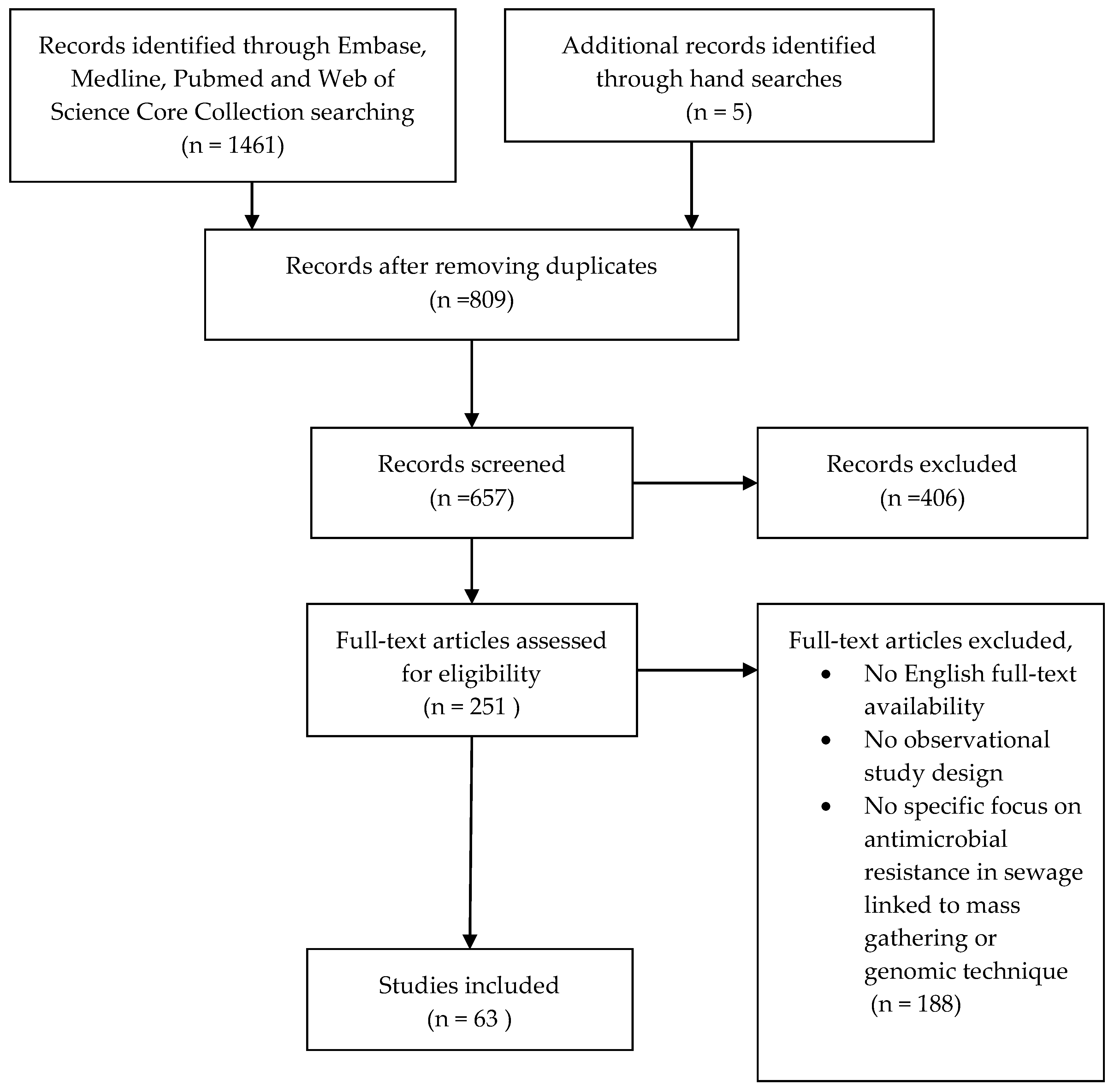
2. Materials and Methods
2.1. Search Strategy
2.2. Selection Criteria
2.3. Data Analysis
3. Results
3.1. Dissemination of Antimicrobial Resistance in Wastewater
3.2. ARB Populations Associated with Wastewaters
3.3. Selective Pressure within Wastewater Environments Promote the Emergence of Novel Variants of ARGs and ARBs
3.4. Hospital Wastewater and the Dissemination of Clinically Relevant ARGs and ARBs Populations
3.5. Impact of Wastewater Treatment Processes on AMR Dissemination
3.6. AMR Dissemination in Wastewater Associated with Mass Gathering Settings
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- World Health Organization. Antimicrobial Resistance: Global Report on Surveillance 2014. Available online: https://www.who.int/drugresistance/documents/surveillancereport/en/ (accessed on 22 June 2019).
- Review on Antimicrobial Resistance. Tackling Drug-Resistant Infections Globally: Final Report and Recommendations. Available online: https://amr-review.org/sites/default/files/160525_Final%20paper_with%20cover.pdf (accessed on 25 June 2019).
- World Health Organization. Antimicrobial Resistance. Available online: https://www.who.int/en/news-room/fact-sheets/detail/antimicrobial-resistance (accessed on 25 June 2019).
- World Health Organization. WHO Report on Surveillance of Antibiotic Consumption 2016–2018 Early Implementation. Available online: https://www.who.int/medicines/areas/rational_use/oms-amr-amc-report-2016-2018/en/ (accessed on 22 June 2019).
- Laxminarayan, R.; Duse, A.; Wattal, C.; Zaidi, A.K.; Wertheim, H.F.; Sumpradit, N.; Vlieghe, E.; Hara, G.L.; Gould, I.M.; Goossens, H.; et al. Antibiotic resistance-the need for global solutions. Lancet Infect. Dis. 2013, 13, 1057–1098. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. Global Antimicrobial Resistance Surveillance System (GLASS) Report Early Implementation 2017–2018. Available online: https://www.who.int/glass/resources/publications/early-implementation-report-2017-2018/en/ (accessed on 26 June 2019).
- Central for Disease Control and Prevention. Infographic: Antibiotic Resistance The Global Threat. Available online: https://www.cdc.gov/globalhealth/infographics/antibiotic-resistance/antibiotic_resistance_global_threat.htm (accessed on 22 June 2019).
- World Health Organization. Report on Global Priority List of Antibiotic-Resistant Bacteria to Guide Research, Dicovery, and Development of New Antibiotics. Available online: https://www.who.int/medicines/publications/global-priority-list-antibiotic-resistant-bacteria/en/ (accessed on 26 June 2019).
- Holmes, A.H.; Moore, L.S.; Sundsfjord, A.; Steinbakk, M.; Regmi, S.; Karkey, A.; Guerin, P.J.; Piddock, L.J. Understanding the mechanisms and drivers of antimicrobial resistance. Lancet 2016, 387, 176–187. [Google Scholar] [CrossRef]
- Schwartz, K.L.; Morris, S.K. Travel and the Spread of Drug-Resistant Bacteria. Curr. Infect. Dis. 2018, 20, 29. [Google Scholar] [CrossRef] [PubMed]
- Memish, Z.A.; Venkatesh, S.; Shibl, A.M. Impact of travel on international spread of antimicrobial resistance. Int. J. Antimicrob. Agents 2003, 21, 135–142. [Google Scholar] [CrossRef]
- Bell, B.G.; Schellevis, F.; Stobberingh, E.; Goossens, H.; Pringle, M. A systematic review and meta-analysis of the effects of antibiotic consumption on antibiotic resistance. BMC Infect. Dis. 2014, 14, 13. [Google Scholar] [CrossRef] [Green Version]
- Van Boeckel, T.P.; Glennon, E.E.; Chen, D.; Gilbert, M.; Robinson, T.P.; Grenfell, B.T.; Levin, S.A.; Bonhoeffer, S.; Laxminarayan, R. Reducing antimicrobial use in food animals. Science 2017, 357, 1350–1352. [Google Scholar] [CrossRef] [Green Version]
- Brinkac, L.; Voorhies, A.; Gomez, A.; Nelson, K.E. The Threat of Antimicrobial Resistance on the Human Microbiome. Microb. Ecol. 2017, 74, 1001–1008. [Google Scholar] [CrossRef]
- Mendelson, M.; Rottingen, J.A.; Gopinathan, U.; Hamer, D.H.; Wertheim, H.; Basnyat, B.; Butler, C.; Tomson, G.; Balasegaram, M. Maximising access to achieve appropriate human antimicrobial use in low-income and middle-income countries. Lancet 2016, 387, 188–198. [Google Scholar] [CrossRef] [PubMed]
- Manyi-Loh, C.; Mamphweli, S.; Meyer, E.; Okoh, A. Antibiotic Use in Agriculture and Its Consequential Resistance in Environmental Sources: Potential Public Health Implications. Molecules 2018, 23, 795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. Antimicrobial and Primary Health Care:Brief. Available online: https://apps.who.int/iris/bitstream/handle/10665/328084/WHO-HIS-SDS-2018.57-eng.pdf (accessed on 26 June 2019).
- Frost, I.; Van Boeckel, T.P.; Pires, J.; Craig, J.; Laxminarayan, R. Global Geographic Trends in Antimicrobial Resistance: The Role of International Travel. J. Travel Med. 2019. [Google Scholar] [CrossRef]
- Woolhouse, M.; Ward, M.; van Bunnik, B.; Farrar, J. Antimicrobial resistance in humans, livestock and the wider environment. Philos. Trans. R. Soc. B Biol. Sci. 2015, 370, 20140083. [Google Scholar] [CrossRef] [PubMed]
- The Parliamentary Office of Science and Technology, H.o.P.U. Reservoir of Antimicrobial Resistance. Available online: https://researchbriefings.parliament.uk/ResearchBriefing/Summary/POST-PN-0595 (accessed on 26 June 2019).
- Food and Agricultural Organization of United Nations. Antimicrobial Resistance in the Environment: Summary Report of an FAO Meeting of Experts FAO Antimicrobial Resistance Working Group. Available online: http://www.fao.org/3/BU656en/bu656en.pdf (accessed on 26 June 2019).
- >European Medicine Agency. Reflection Paper on Antimicrobial Resistance in the Environment: Considerations for Current and Future Risk Assessment of Veterinary Medicinal Products (Draft). Available online: https://www.ema.europa.eu/en/documents/scientific-guideline/draft-reflection-paper-antimicrobial-resistance-environment-considerations-current-future-risk_en.pdf (accessed on 26 June 2019).
- Wellington, E.M.; Boxall, A.B.; Cross, P.; Feil, E.J.; Gaze, W.H.; Hawkey, P.M.; Johnson-Rollings, A.S.; Jones, D.L.; Lee, N.M.; Otten, W.; et al. The role of the natural environment in the emergence of antibiotic resistance in gram-negative bacteria. Lancet Infect. Dis. 2013, 13, 155–165. [Google Scholar] [CrossRef]
- Balcazar, J.L. Bacteriophages as vehicles for antibiotic resistance genes in the environment. PLoS Pathog. 2014, 10, e1004219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berglund, B. Environmental dissemination of antibiotic resistance genes and correlation to anthropogenic contamination with antibiotics. Infect. Ecol. Epidemiol. 2015, 5, 28564. [Google Scholar] [CrossRef]
- Jiang, X.; Ellabaan, M.M.H.; Charusanti, P.; Munck, C.; Blin, K.; Tong, Y.; Weber, T.; Sommer, M.O.A.; Lee, S.Y. Dissemination of antibiotic resistance genes from antibiotic producers to pathogens. Nat. Commun. 2017, 8, 15784. [Google Scholar] [CrossRef] [Green Version]
- Hocquet, D.; Muller, A.; Bertrand, X. What happens in hospitals does not stay in hospitals: Antibiotic-resistant bacteria in hospital wastewater systems. J. Hosp. Infect. 2016, 93, 395–402. [Google Scholar] [CrossRef]
- Bouki, C.; Venieri, D.; Diamadopoulos, E. Detection and fate of antibiotic resistant bacteria in wastewater treatment plants: A review. Ecotox. Environ. Safe. 2013, 91, 1–9. [Google Scholar] [CrossRef]
- McKinney, C.W.; Dungan, R.S.; Moore, A.; Leytem, A.B. Occurrence and abundance of antibiotic resistance genes in agricultural soil receiving dairy manure. FEMS Microbiol. Ecol. 2018, 94. [Google Scholar] [CrossRef]
- Rizzo, L.; Fiorentino, A.; Anselmo, A. Advanced treatment of urban wastewater by UV radiation: Effect on antibiotics and antibiotic-resistant E. coli strains. Chemosphere 2013, 92, 171–176. [Google Scholar] [CrossRef]
- Rizzo, L.; Manaia, C.; Merlin, C.; Schwartz, T.; Dagot, C.; Ploy, M.C.; Michael, I.; Fatta-Kassinos, D. Urban wastewater treatment plants as hotspots for antibiotic resistant bacteria and genes spread into the environment: A review. Sci. Total Environ. 2013, 447, 345–360. [Google Scholar] [CrossRef]
- World Health Organization. Antimicrobial resistance: An Emerging Water, Sanitation and Hygiene Issue. Briefing Note. Available online: https://www.who.int/water_sanitation_health/emerging/AMR_briefing_note.pdf (accessed on 22 June 2019).
- Treangen, T.J.; Rocha, E.P. Horizontal transfer, not duplication, drives the expansion of protein families in prokaryotes. PLoS Genet. 2011, 7, e1001284. [Google Scholar] [CrossRef] [PubMed]
- Stecher, B.; Denzler, R.; Maier, L.; Bernet, F.; Sanders, M.J.; Pickard, D.J.; Barthel, M.; Westendorf, A.M.; Krogfelt, K.A.; Walker, A.W.; et al. Gut inflammation can boost horizontal gene transfer between pathogenic and commensal Enterobacteriaceae. Proc. Natl. Acad. Sci. USA 2012, 109, 1269–1274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nahar, A.; Islam, M.A.; Sobur, M.A.; Hossain, M.J.; Binte, S.; Zaman, M.; Rahman, B.; Kabir, S.L.; Rahman, M.T. Detection of tetracycline resistant E. coli and Salmonella spp. in sewage, river, pond and swimming pool in Mymensingh, Bangladesh. Afr. J. Microbiol. Res. 2018. [Google Scholar] [CrossRef]
- Qin, J.; Maixnerova, M.; Nemec, M.; Feng, Y.; Zhang, X.; Nemec, A.; Zong, Z. Acinetobacter cumulans sp. nov., isolated from hospital sewage and capable of acquisition of multiple antibiotic resistance genes. Syst. Appl. Microbiol. 2019, 42, 319–325. [Google Scholar] [CrossRef] [PubMed]
- Haberecht, H.B.; Nealon, N.J.; Gilliland, J.R.; Holder, A.V.; Runyan, C.; Oppel, R.C.; Ibrahim, H.M.; Mueller, L.; Schrupp, F.; Vilchez, S.; et al. Antimicrobial-Resistant Escherichia coli from Environmental Waters in Northern Colorado. J. Environ. Public Health 2019, 2019, 3862949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sekizuka, T.; Yatsu, K.; Inamine, Y.; Segawa, T.; Nishio, M.; Kishi, N.; Kuroda, M. Complete Genome Sequence of a blaKPC-2-Positive Klebsiella pneumoniae Strain Isolated from the Effluent of an Urban Sewage Treatment Plant in Japan. mSphere 2018, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cahill, N.; O’Connor, L.; Mahon, B.; Varley, A.; McGrath, E.; Ryan, P.; Cormican, M.; Brehony, C.; Jolley, K.A.; Maiden, M.C.; et al. Hospital effluent: A reservoir for carbapenemase-producing Enterobacterales? Sci. Total Environ. 2019, 672, 618–624. [Google Scholar] [CrossRef] [PubMed]
- Niestepski, S.; Harnisz, M.; Korzeniewska, E.; Aguilera-Arreola, M.G.; Contreras-Rodriguez, A.; Filipkowska, Z.; Osinska, A. The emergence of antimicrobial resistance in environmental strains of the Bacteroides fragilis group. Environ. Int. 2019, 124, 408–419. [Google Scholar] [CrossRef]
- Hendriksen, R.S.; Munk, P.; Njage, P.; van Bunnik, B.; McNally, L.; Lukjancenko, O.; Roder, T.; Nieuwenhuijse, D.; Pedersen, S.K.; Kjeldgaard, J.; et al. Global monitoring of antimicrobial resistance based on metagenomics analyses of urban sewage. Nat. Commun. 2019, 10, 1124. [Google Scholar] [CrossRef]
- Khan, F.A.; Soderquist, B.; Jass, J. Prevalence and Diversity of Antibiotic Resistance Genes in Swedish Aquatic Environments Impacted by Household and Hospital Wastewater. Front. Microbiol. 2019, 10, 688. [Google Scholar] [CrossRef] [Green Version]
- Tokajian, S.; Moghnieh, R.; Salloum, T.; Arabaghian, H.; Alousi, S.; Moussa, J.; Abboud, E.; Youssef, S.; Husni, R. Extended-spectrum beta-lactamase-producing Escherichia coli in wastewaters and refugee camp in Lebanon. Future Microbiol. 2018, 13, 81–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parnanen, K.M.M.; Narciso-da-Rocha, C.; Kneis, D.; Berendonk, T.U.; Cacace, D.; Do, T.T.; Elpers, C.; Fatta-Kassinos, D.; Henriques, I.; Jaeger, T.; et al. Antibiotic resistance in European wastewater treatment plants mirrors the pattern of clinical antibiotic resistance prevalence. Sci. Adv. 2019, 5, eaau9124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bougnom, B.P.; Zongo, C.; McNally, A.; Ricci, V.; Etoa, F.X.; Thiele-Bruhn, S.; Piddock, L.J.V. Wastewater used for urban agriculture in West Africa as a reservoir for antibacterial resistance dissemination. Environ. Res. 2019, 168, 14–24. [Google Scholar] [CrossRef] [PubMed]
- Gouliouris, T.; Raven, K.E.; Moradigaravand, D.; Ludden, C.; Coll, F.; Blane, B.; Naydenova, P.; Horner, C.; Brown, N.M.; Corander, J.; et al. Detection of vancomycin-resistant Enterococcus faecium hospital-adapted lineages in municipal wastewater treatment plants indicates widespread distribution and release into the environment. Genome Res. 2019, 29, 626–634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iweriebor, B.C.; Gaqavu, S.; Obi, L.C.; Nwodo, U.U.; Okoh, A.I. Antibiotic susceptibilities of enterococcus species isolated from hospital and domestic wastewater effluents in alice, eastern cape province of South Africa. Int. J. Environ. Res. Public Health 2015, 12, 4231–4246. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Li, J.; Chen, H.; Bond, P.L.; Yuan, Z. Metagenomic analysis reveals wastewater treatment plants as hotspots of antibiotic resistance genes and mobile genetic elements. Water Res. 2017, 123, 468–478. [Google Scholar] [CrossRef]
- Wang, J.; Mao, D.; Mu, Q.; Luo, Y. Fate and proliferation of typical antibiotic resistance genes in five full-scale pharmaceutical wastewater treatment plants. Sci. Total Environ. 2015, 526, 366–373. [Google Scholar] [CrossRef]
- Conte, D.; Palmeiro, J.K.; da Silva Nogueira, K.; de Lima, T.M.; Cardoso, M.A.; Pontarolo, R.; Degaut Pontes, F.L.; Dalla-Costa, L.M. Characterization of CTX-M enzymes, quinolone resistance determinants, and antimicrobial residues from hospital sewage, wastewater treatment plant, and river water. Ecotoxicol. Environ. Saf. 2017, 136, 62–69. [Google Scholar] [CrossRef]
- Baumlisberger, M.; Youssar, L.; Schilhabel, M.B.; Jonas, D. Influence of a non-hospital medical care facility on antimicrobial resistance in wastewater. PLoS ONE 2015, 10, e0122635. [Google Scholar] [CrossRef] [Green Version]
- Adefisoye, M.A.; Okoh, A.I. Identification and antimicrobial resistance prevalence of pathogenic Escherichia coli strains from treated wastewater effluents in Eastern Cape, South Africa. Microbiol. Open 2016, 5, 143–151. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, S.; Ogo, M.; Koike, T.; Takada, H.; Newman, B. Sulfonamide and tetracycline resistance genes in Total- and culturable-bacterial assemblages in South African aquatic environments. Front. Microbiol. 2015, 6, 796. [Google Scholar] [CrossRef] [PubMed]
- Froes, A.M.; da Mota, F.F.; Cuadrat, R.R.; Davila, A.M. Distribution and Classification of Serine beta-Lactamases in Brazilian Hospital Sewage and Other Environmental Metagenomes Deposited in Public Databases. Front. Microbiol. 2016, 7, 1790. [Google Scholar] [CrossRef] [PubMed]
- Laht, M.; Karkman, A.; Voolaid, V.; Ritz, C.; Tenson, T.; Virta, M.; Kisand, V. Abundances of tetracycline, sulphonamide and beta-lactam antibiotic resistance genes in conventional wastewater treatment plants (WWTPs) with different waste load. PLoS ONE 2014, 9, e103705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walsh, T.R.; Weeks, J.; Livermore, D.M.; Toleman, M.A. Dissemination of NDM-1 positive bacteria in the New Delhi environment and its implications for human health: An environmental point prevalence study. Lancet Infect. Dis. 2011, 11, 355–362. [Google Scholar] [CrossRef]
- Zhang, T.; Zhang, X.X.; Ye, L. Plasmid metagenome reveals high levels of antibiotic resistance genes and mobile genetic elements in activated sludge. PLoS ONE 2011, 6, e26041. [Google Scholar] [CrossRef] [Green Version]
- Chagas, T.P.; Seki, L.M.; Cury, J.C.; Oliveira, J.A.; Davila, A.M.; Silva, D.M.; Asensi, M.D. Multiresistance, beta-lactamase-encoding genes and bacterial diversity in hospital wastewater in Rio de Janeiro, Brazil. J. Appl. Microbiol. 2011, 111, 572–581. [Google Scholar] [CrossRef]
- Szczepanowski, R.; Linke, B.; Krahn, I.; Gartemann, K.H.; Gutzkow, T.; Eichler, W.; Puhler, A.; Schluter, A. Detection of 140 clinically relevant antibiotic-resistance genes in the plasmid metagenome of wastewater treatment plant bacteria showing reduced susceptibility to selected antibiotics. Microbiology 2009, 155, 2306–2319. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Yu, T.; Zhang, Y.; Yang, M.; Li, Z.; Liu, M.; Qi, R. Antibiotic resistance characteristics of environmental bacteria from an oxytetracycline production wastewater treatment plant and the receiving river. Appl. Environ. Microbiol. 2010, 76, 3444–3451. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; He, H.; Chen, S.; Huang, T.; Lu, K.; Zhang, Z.; Wang, R.; Zhang, X.; Li, H. Abundance of antibiotic resistance genes and their association with bacterial communities in activated sludge of wastewater treatment plants: Geographical distribution and network analysis. J. Environ. Sci. 2019, 82, 24–38. [Google Scholar] [CrossRef]
- Li, B.; Yang, Y.; Ma, L.; Ju, F.; Guo, F.; Tiedje, J.M.; Zhang, T. Metagenomic and network analysis reveal wide distribution and co-occurrence of environmental antibiotic resistance genes. ISME 2015, 9, 2490–2502. [Google Scholar] [CrossRef] [Green Version]
- Hembach, N.; Schmid, F.; Alexander, J.; Hiller, C.; Rogall, E.T.; Schwartz, T. Occurrence of the mcr-1 Colistin Resistance Gene and other Clinically Relevant Antibiotic Resistance Genes in Microbial Populations at Different Municipal Wastewater Treatment Plants in Germany. Front. Microbiol. 2017, 8, 1282. [Google Scholar] [CrossRef] [PubMed]
- Igbinosa, I.H.; Okoh, A.I. Antibiotic susceptibility profile of Aeromonas species isolated from wastewater treatment plant. Sci. World J. 2012, 2012, 764563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Igbinosa, E.O.; Obi, L.C.; Tom, M.; Okoh, A.I. Detection of potential risk of wastewater effluents for transmission of antibiotic resistance from Vibrio species as a reservoir in a peri-urban community in South Africa. Int. J. Environ. Health Res. 2011, 21, 402–414. [Google Scholar] [CrossRef] [PubMed]
- Johnning, A.; Kristiansson, E.; Fick, J.; Weijdegard, B.; Larsson, D.G. Resistance Mutations in gyrA and parC are Common in Escherichia Communities of both Fluoroquinolone-Polluted and Uncontaminated Aquatic Environments. Front. Microbiol. 2015, 6, 1355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sahlström, L.; Rehbinder, V.; Albihn, A.; Aspan, A.; Bengtsson, B. Vancomycin resistant enterococci (VRE) in Swedish sewage sludge. Acta Vet. Scand. 2009, 51, 24. [Google Scholar] [CrossRef] [Green Version]
- Araújo, C.; Torres, C.; Silva, N.; Carneiro, C.; Gonçalves, A.; Radhouani, H.; Correia, S.; da Costa, P.M.; Paccheco, R.; Zarazaga, M. Vancomycin-resistant enterococci from Portuguese wastewater treatment plants. J. Basic Microbiol. 2010, 50, 605–609. [Google Scholar] [CrossRef]
- Soge, O.; Tivoli, L.; Meschke, J.; Roberts, M. A conjugative macrolide resistance gene, mef(A), in environmental Clostridium perfringens carrying multiple macrolide and/or tetracycline resistance genes. J. Appl. Microbiol. 2009, 106, 34–40. [Google Scholar] [CrossRef]
- Zhang, X.-X.; Zhang, T.; Zhang, M.; Fang, H.H.P.; Cheng, S.-P. Characterization and quantification of class 1 integrons and associated gene cassettes in sewage treatment plants. Appl. Microbiol. Biotechnol. 2009, 82, 1169–1177. [Google Scholar] [CrossRef]
- Odjadjare, E.E.O.; Obi, L.C.; Okoh, A.I. Municipal Wastewater Effluents as a Source of Listerial Pathogens in the Aquatic Milieu of the Eastern Cape Province of South Africa: A Concern of Public Health Importance. Int. J. Environ. Res. Public Health 2010, 7, 2376–2394. [Google Scholar] [CrossRef]
- Okoh, A.I.; Igbinosa, E.O. Antibiotic susceptibility profiles of some Vibrio strains isolated from wastewater final effluents in a rural community of the Eastern Cape Province of South Africa. BMC Microbiol. 2010, 10, 143. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Byelashov, O.A.; Geornaras, I.; Goodridge, L.D.; Nightingale, K.K.; Belk, K.E.; Smith, G.C.; Sofos, J.N. Presence of antibiotic-resistant commensal bacteria in samples from agricultural, city, and national park environments evaluated by standard culture and real-time PCR methods. Can. J. Microbiol. 2010, 56, 761–770. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Armisen, T.; Vercammen, K.; Passerat, J.; Triest, D.; Servais, P.; Cornelis, P. Antimicrobial resistance of heterotrophic bacteria in sewage-contaminated rivers. Water Res. 2011, 45, 788–796. [Google Scholar] [CrossRef] [PubMed]
- Colomer-Lluch, M.; Jofre, J.; Muniesa, M. Antibiotic Resistance Genes in the Bacteriophage DNA Fraction of Environmental Samples. PLoS ONE 2011, 6, e17549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuentefria, D.B.; Ferreira, A.E.; Corcao, G. Antibiotic-resistant Pseudomonas aeruginosa from hospital wastewater and superficial water: Are they genetically related? J. Environ. Manag. 2011, 92, 250–255. [Google Scholar] [CrossRef] [PubMed]
- Gaze, W.H.; Zhang, L.; Abdouslam, N.A.; Hawkey, P.M.; Calvo-Bado, L.; Royle, J.; Brown, H.; Davis, S.; Kay, P.; Boxall, A.B.; et al. Impacts of anthropogenic activity on the ecology of class 1 integrons and integron-associated genes in the environment. ISME 2011, 5, 1253–1261. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.; Zhang, X.X.; Cheng, S.; Zhang, Z.; Shi, P.; Liu, B.; Wu, B.; Zhang, Y. Occurrence, abundance and elimination of class 1 integrons in one municipal sewage treatment plant. Ecotoxicology 2011, 20, 968–973. [Google Scholar] [CrossRef]
- Mokracka, J.; Koczura, R.; Jabłońska, L.; Kaznowski, A. Phylogenetic groups, virulence genes and quinolone resistance of integron-bearing Escherichia coli strains isolated from a wastewater treatment plant. Antonie Van Leeuwenhoek 2011, 99, 817–824. [Google Scholar] [CrossRef] [Green Version]
- Amaya, E.; Reyes, D.; Paniagua, M.; Calderón, S.; Rashid, M.U.; Colque, P.; Kühn, I.; Möllby, R.; Weintraub, A.; Nord, C.E. Antibiotic resistance patterns of Escherichia coli isolates from different aquatic environmental sources in Leon, Nicaragua. Clin. Microbiol. Infect. 2012, 18, E347–E354. [Google Scholar] [CrossRef] [Green Version]
- Mokracka, J.; Koczura, R.; Kaznowski, A. Multiresistant Enterobacteriaceae with class 1 and class 2 integrons in a municipal wastewater treatment plant. Water Res. 2012, 46, 3353–3363. [Google Scholar] [CrossRef]
- Spindler, A.; Otton, L.M.; Fuentefria, D.B.; Corcao, G. Beta-lactams resistance and presence of class 1 integron in Pseudomonas spp. isolated from untreated hospital effluents in Brazil. Antonie Van Leeuwenhoek 2012, 102, 73–81. [Google Scholar] [CrossRef]
- Gundogdu, A.; Jennison, A.V.; Smith, H.V.; Stratton, H.; Katouli, M. Extended-spectrum beta-lactamase producing Escherichia coli in hospital wastewaters and sewage treatment plants in Queensland, Australia. Can. J. Microbiol. 2013, 59, 737–745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zarfel, G.; Galler, H.; Feierl, G.; Haas, D.; Kittinger, C.; Leitner, E.; Grisold, A.J.; Mascher, F.; Posch, J.; Pertschy, B.; et al. Comparison of extended-spectrum-beta-lactamase (ESBL) carrying Escherichia coli from sewage sludge and human urinary tract infection. Environ. Pollut. 2013, 173, 192–199. [Google Scholar] [CrossRef] [PubMed]
- Colomer-Lluch, M.; Mora, A.; Lopez, C.; Mamani, R.; Dahbi, G.; Marzoa, J.; Herrera, A.; Viso, S.; Blanco, J.E.; Blanco, M.; et al. Detection of quinolone-resistant Escherichia coli isolates belonging to clonal groups O25b: H4-B2-ST131 and O25b: H4-D-ST69 in raw sewage and river water in Barcelona, Spain. J. Antimicrob. Chemother. 2013, 68, 758–765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sadowy, E.; Luczkiewicz, A. Drug-resistant and hospital-associated Enterococcus faecium from wastewater, riverine estuary and anthropogenically impacted marine catchment basin. BMC Microbiol. 2014, 14, 66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, P.; He, S.; Huang, S.; Li, K.; Liu, Z.; Xue, G.; Sun, W. Impacts of coexisting antibiotics, antibacterial residues, and heavy metals on the occurrence of erythromycin resistance genes in urban wastewater. Appl. Microbiol. Biotechnol. 2015, 99, 3971–3980. [Google Scholar] [CrossRef]
- Nishiyama, M.; Iguchi, A.; Suzuki, Y. Identification of Enterococcus faecium and Enterococcus faecalis as vanC-type Vancomycin-Resistant Enterococci (VRE) from sewage and river water in the provincial city of Miyazaki, Japan. J. Environ. Sci. Health 2015, 50, 16–25. [Google Scholar] [CrossRef]
- Zhang, S.; Han, B.; Gu, J.; Wang, C.; Wang, P.; Ma, Y.; Cao, J.; He, Z. Fate of antibiotic resistant cultivable heterotrophic bacteria and antibiotic resistance genes in wastewater treatment processes. Chemosphere 2015, 135, 138–145. [Google Scholar] [CrossRef]
- Simo Tchuinte, P.L.; Stalder, T.; Venditti, S.; Ngandjio, A.; Dagot, C.; Ploy, M.C.; Barraud, O. Characterisation of class 3 integrons with oxacillinase gene cassettes in hospital sewage and sludge samples from France and Luxembourg. Int. J. Antimicrob. Agents 2016, 48, 431–434. [Google Scholar] [CrossRef]
- Young, S.; Nayak, B.; Sun, S.; Badgley, B.D.; Rohr, J.R.; Harwood, V.J. Vancomycin-Resistant Enterococci and Bacterial Community Structure following a Sewage Spill into an Aquatic Environment. Appl. Environ. Microbiol. 2016, 82, 5653. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Shin, S.G.; Jang, H.M.; Kim, Y.B.; Lee, J.; Kim, Y.M. Characterization of antibiotic resistance genes in representative organic solid wastes: Food waste-recycling wastewater, manure, and sewage sludge. Sci. Total Environ. 2017, 579, 1692–1698. [Google Scholar] [CrossRef] [PubMed]
- An, X.L.; Chen, Q.L.; Zhu, D.; Zhu, Y.G.; Gillings, M.R.; Su, J.Q. Impact of Wastewater Treatment on the Prevalence of Integrons and the Genetic Diversity of Integron Gene Cassettes. Appl. Environ. Microbiol. 2018, 84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haller, L.; Chen, H.; Ng, C.; Le, T.H.; Koh, T.H.; Barkham, T.; Sobsey, M.; Gin, K.Y. Occurrence and characteristics of extended-spectrum beta-lactamase- and carbapenemase- producing bacteria from hospital effluents in Singapore. Sci. Total Environ. 2018, 615, 1119–1125. [Google Scholar] [CrossRef] [PubMed]
- Galler, H.; Feierl, G.; Petternel, C.; Reinthaler, F.F.; Haas, D.; Habib, J.; Kittinger, C.; Luxner, J.; Zarfel, G. Multiresistant Bacteria Isolated from Activated Sludge in Austria. Int. J. Environ. Res. Public Health 2018, 15, 479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quach-Cu, J.; Herrera-Lynch, B.; Marciniak, C.; Adams, S.; Simmerman, A.; Reinke, R.A. The Effect of Primary, Secondary, and Tertiary Wastewater Treatment Processes on Antibiotic Resistance Gene (ARG) Concentrations in Solid and Dissolved Wastewater Fractions. Water 2018, 10, 37. [Google Scholar] [CrossRef] [Green Version]
- Yousfi, K.; Touati, A.; Lefebvre, B.; Garneau, P.; Brahmi, S.; Gharout-Sait, A.; Harel, J.; Bekal, S. Characterization of multidrug-resistant Gram-negative bacilli isolated from hospitals effluents: First report of a blaOXA-48-like in Klebsiella oxytoca, Algeria. Braz. J. Microbiol. 2019, 50, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Flandroy, L.; Poutahidis, T.; Berg, G.; Clarke, G.; Dao, M.C.; Decaestecker, E.; Furman, E.; Haahtela, T.; Massart, S.; Plovier, H.; et al. The impact of human activities and lifestyles on the interlinked microbiota and health of humans and of ecosystems. Sci. Total Environ. 2018, 627, 1018–1038. [Google Scholar] [CrossRef]
- Hiller, C.X.; Hubner, U.; Fajnorova, S.; Schwartz, T.; Drewes, J.E. Antibiotic microbial resistance (AMR) removal efficiencies by conventional and advanced wastewater treatment processes: A review. Sci. Total Environ. 2019, 685, 596–608. [Google Scholar] [CrossRef]
- Ahmed, Q.A.; Arabi, Y.M.; Memish, Z.A. Health risks at the Hajj. Lancet 2006, 367, 1008–1015. [Google Scholar] [CrossRef]
- Sridhar, S.; Gautret, P.; Brouqui, P. A comprehensive review of the Kumbh Mela: Identifying risks for spread of infectious diseases. Clin. Microbiol. Infect. 2015, 21, 128–133. [Google Scholar] [CrossRef] [Green Version]
- Mutreja, A.; Kim, D.W.; Thomson, N.R.; Connor, T.R.; Lee, J.H.; Kariuki, S.; Croucher, N.J.; Choi, S.Y.; Harris, S.R.; Lebens, M.; et al. Evidence for several waves of global transmission in the seventh cholera pandemic. Nature 2011, 477, 462–465. [Google Scholar] [CrossRef] [Green Version]
- Abd El Ghany, M.; Chander, J.; Mutreja, A.; Rashid, M.; Hill-Cawthorne, G.A.; Ali, S.; Naeem, R.; Thomson, N.R.; Dougan, G.; Pain, A. The population structure of Vibrio cholerae from the Chandigarh Region of Northern India. PLoS Negl. Trop. Dis. 2014, 8, e2981. [Google Scholar] [CrossRef] [PubMed]
- Reddy, B.; Dubey, S.K. River Ganges water as reservoir of microbes with antibiotic and metal ion resistance genes: High throughput metagenomic approach. Environ. Pollut. 2019, 246, 443–451. [Google Scholar] [CrossRef] [PubMed]
- Samson, R.; Shah, M.; Yadav, R.; Sarode, P.; Rajput, V.; Dastager, S.G.; Dharne, M.S.; Khairnar, K. Metagenomic insights to understand transient influence of Yamuna River on taxonomic and functional aspects of bacterial and archaeal communities of River Ganges. Sci. Total Environ. 2019, 674, 288–299. [Google Scholar] [CrossRef] [PubMed]
- Abd El Ghany, M.; Alsomali, M.; Almasri, M.; Padron Regalado, E.; Naeem, R.; Tukestani, A.; Asiri, A.; Hill-Cawthorne, G.A.; Pain, A.; Memish, Z.A. Enteric Infections Circulating during Hajj Seasons, 2011–2013. Emerg. Infect. Dis. 2017, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abd El Ghany, M.; Al-Tawfiq, J.A.; Hill-Cawthorne, G.A.; Memish, Z.A. Hajj-Beyond traveller’s diarrhea. Travel Med. Infect. Dis. 2018, 21, 80–81. [Google Scholar] [CrossRef] [Green Version]
- Leangapichart, T.; Rolain, J.M.; Memish, Z.A.; Al-Tawfiq, J.A.; Gautret, P. Emergence of drug resistant bacteria at the Hajj: A systematic review. Travel Med. Infect. Dis. 2017, 18, 3–17. [Google Scholar] [CrossRef] [Green Version]
- Abd El Ghany, M.; Sharaf, H.; Hill-Cawthorne, G.A. Hajj vaccinations-facts, challenges, and hope. Int. J. Infect. Dis. 2016, 47, 29–37. [Google Scholar] [CrossRef] [Green Version]
- Leangapichart, T.; Gautret, P.; Griffiths, K.; Belhouchat, K.; Memish, Z.; Raoult, D.; Rolain, J.M. Acquisition of a High Diversity of Bacteria during the Hajj Pilgrimage, Including Acinetobacter baumannii with blaOXA-72 and Escherichia coli with blaNDM-5 Carbapenemase Genes. Antimicrob. Agents Chemother. 2016, 60, 5942–5948. [Google Scholar] [CrossRef] [Green Version]
- Olaitan, A.O.; Dia, N.M.; Gautret, P.; Benkouiten, S.; Belhouchat, K.; Drali, T.; Parola, P.; Brouqui, P.; Memish, Z.; Raoult, D.; et al. Acquisition of extended-spectrum cephalosporin- and colistin-resistant Salmonella enterica subsp. enterica serotype Newport by pilgrims during Hajj. Int. J. Antimicrob. Agents 2015, 45, 600–604. [Google Scholar] [CrossRef]
- World Health Organization. Global Action Plan on Antimicrobial Resistance. Available online: https://www.who.int/antimicrobial-resistance/publications/global-action-plan/en/ (accessed on 22 June 2019).
- Lothigius, A.; Sjoling, A.; Svennerholm, A.M.; Bolin, I. Survival and gene expression of enterotoxigenic Escherichia coli during long-term incubation in sea water and freshwater. J. Appl. Microbiol. 2010, 108, 1441–1449. [Google Scholar] [CrossRef] [Green Version]
- Hernroth, B.; Lothigius, A.; Bolin, I. Factors influencing survival of enterotoxigenic Escherichia coli, Salmonella enterica (serovar Typhimurium) and Vibrio parahaemolyticus in marine environments. FEMS Microbiol. Ecol. 2010, 71, 272–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kingsley, R.A.; Langridge, G.; Smith, S.E.; Makendi, C.; Fookes, M.; Wileman, T.M.; El Ghany, M.A.; Keith Turner, A.; Dyson, Z.A.; Sridhar, S.; et al. Functional analysis of Salmonella Typhi adaptation to survival in water. Environ. Microbiol. 2018, 20, 4079–4090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jani, K.; Bandal, J.; Rale, V.; Shouche, Y.; Sharma, A. Antimicrobial resistance pattern of microorganisms isolated and identified from Godavari River across the mass gathering event. J. Biosci. 2019, 44, 121. [Google Scholar] [CrossRef] [PubMed]
- Diwan, V.; Purohit, M.; Chandran, S.; Parashar, V.; Shah, H.; Mahadik, V.K.; Stalsby Lundborg, C.; Tamhankar, A.J. A Three-Year Follow-Up Study of Antibiotic and Metal Residues, Antibiotic Resistance and Resistance Genes, Focusing on Kshipra-A River Associated with Holy Religious Mass-Bathing in India: Protocol Paper. Int. J. Environ. Res. Public Health 2017, 14, 574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jani, K.; Dhotre, D.; Bandal, J.; Shouche, Y.; Suryavanshi, M.; Rale, V.; Sharma, A. World’s Largest Mass Bathing Event Influences the Bacterial Communities of Godavari, a Holy River of India. Microbial. Ecol. 2018, 76, 706–718. [Google Scholar] [CrossRef]
- Iredell, J.; Brown, J.; Tagg, K. Antibiotic resistance in Enterobacteriaceae: Mechanisms and clinical implications. BMJ 2016, 352, h6420. [Google Scholar] [CrossRef]
- Partridge, S.R. Analysis of antibiotic resistance regions in Gram-negative bacteria. FEMS Microbiol. Rev. 2011, 35, 820–855. [Google Scholar] [CrossRef] [Green Version]
- Partridge, S.R. Resistance mechanisms in Enterobacteriaceae. Pathology 2015, 47, 276–284. [Google Scholar] [CrossRef]
- Wright, G.D. Antibiotic resistance in the environment: A link to the clinic? Curr. Opp. Microbiol. 2010, 13, 589–594. [Google Scholar] [CrossRef]
- Pruden, A. Balancing water sustainability and public health goals in the face of growing concerns about antibiotic resistance. Environ. Sci. Tech. 2014, 48, 5–14. [Google Scholar] [CrossRef]
- Amos, G.C.; Hawkey, P.M.; Gaze, W.H.; Wellington, E.M. Waste water effluent contributes to the dissemination of CTX-M-15 in the natural environment. J. Antimicrob. Chemother. 2014, 69, 1785–1791. [Google Scholar] [CrossRef] [PubMed]
- Dropa, M.; Lincopan, N.; Balsalobre, L.C.; Oliveira, D.E.; Moura, R.A.; Fernandes, M.R.; da Silva, Q.M.; Matte, G.R.; Sato, M.I.; Matte, M.H. Genetic background of novel sequence types of CTX-M-8- and CTX-M-15-producing Escherichia coli and Klebsiella pneumoniae from public wastewater treatment plants in Sao Paulo, Brazil. Environ. Sci. Pollut. Res. Int. 2016, 23, 4953–4958. [Google Scholar] [CrossRef] [PubMed]
- Amos, G.C.; Zhang, L.; Hawkey, P.M.; Gaze, W.H.; Wellington, E.M. Functional metagenomic analysis reveals rivers are a reservoir for diverse antibiotic resistance genes. Vet. Microbiol. 2014, 171, 441–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Zhang, X.X.; Huang, K.; Miao, Y.; Shi, P.; Liu, B.; Long, C.; Li, A. Metagenomic profiling of antibiotic resistance genes and mobile genetic elements in a tannery wastewater treatment plant. PLoS ONE 2013, 8, e76079. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Li, B.; Zou, S.; Fang, H.H.; Zhang, T. Fate of antibiotic resistance genes in sewage treatment plant revealed by metagenomic approach. Water Res. 2014, 62, 97–106. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| No. | References | Source of Samples | Types of Investigated Samples | Key Findings |
|---|---|---|---|---|
| 1 | Nahar et al., 2019 [35] | Sewage and environmental water | Sewage, river, pond and swimming pool water | AMR bacteria of E. coli and Salmonella spp. were detected in all environmental samples. |
| 2 | Qin et al., 2019 [36] | Sewage | Hospital sewage | Novel species (Acinetobacter cumulans) containing ARGs conferring resistance to carbapenems, cephalosporin or aminoglycoside were identified. |
| 3 | Haberecht et al., 2019 [37] | Sewage and environmental waters | Sewage water, wastewater treatment plant (WWTP) (influent and effluent) and surface water (ambient water) | Increased abundance of ARB and multidrug resistant (MDR) strains were detected in influent compared to effluent wastewater. Extended-spectrum β-lactamases (ESBL)-producing E. coli strains have been identified in environmental surface water. |
| 4 | Sekizuka et al., 2018 [38] | Sewage | WWTP effluent | Carbapenem-producing strain of K. pneumonia carrying blaKPC-2 was detected. This novel resistant strain rarely detected in clinical settings in Japan. |
| 5 | Cahill et al., 2019 [39] | Sewage and environmental wastewater | Hospital and municipal wastewater (pre- and post-hospital) | Higher rates of carbapenemase-producing Enterobacteriaceae (CPE) have been detected in hospital effluent. |
| 6 | Niestepski et al., 2019 [40] | Sewage, environmental water and human faeces | Hospital wastewater, WWTP (influent and effluent) and human faeces | The highest drug-resistance levels were observed in the strains isolated from influent and effluent WWTP water. Bacteria of Bacteroides fragilis group (BFG) isolated from the WWTPs characterised by higher resistant profiles than those that have been recovered from human and rat faeces. |
| 7 | Hendrieksen et al., 2019 [41] | Sewage | Domestic sewage | Clinically relevant ARGs associated with resistance to macrolides, tetracycline, aminoglycoside, beta-lactams and sulfonamides were identified. The abundance of ARGs were higher in samples collected from low-income compared to high-income countries. |
| 8 | Khan et al., 2019 [42] | Sewage and environmental waters | Hospital wastewater, WWTP samples and downstream water | β-lactamase genes, including blaIMP−1, blaIMP−2 and blaOXA−23, were detected only in hospital sewage, while blaOXA−48, blaCTX−M−8 and blaSFC−1, blaVIM−1 and blaVIM−13 were only detected in downstream river water but not in the WWTP. |
| 9 | Tokajian et al., 2018 [43] | Sewage and environmental waters | Refugee camp sewage water and rivers effluent | Higher rates of AMR E. coli isolates, including ESBL-producing strains, and those which showed resistance to different antimicrobial drugs, including aminoglycosides, fluoroquinolones and trimethoprim/sulfamethosazole, were detected in samples from refugee camps. |
| 10 | Parnanen et al., 2019 [44] | Sewage | Influent and effluent WWTPs samples from different countries | Significantly higher rates of ARGs were identified in effluent samples from low-income compared to high-income countries. |
| 11 | Bougnom et al., 2019 [45] | Sewage | Urban wastewater for agriculture (three canals with different settings) | Higher rates of ARG that confer resistance to 11 major antimicrobial drug groups, including aminoglycoside, tetracycline, beta-lactams and macrolides, were detected in urban wastewater. There was difference in the composition of ARGs associated with ESBL within city water from three canals that received water from different environments, including hospitals. |
| 12 | Gouliouris et al., 2019 [46] | Sewage | Municipal wastewater (untreated and treated) and hospital sewage | Higher rates of vancomycin and ampicillin-resistant E. faecium closely related to hospital isolates have been detected in untreated wastewater plants receiving directly from hospital sewage. |
| 13 | Iweriebor et al., 2015 [47] | Sewage | Municipal and hospital wastewater | Ninety-one percent and 100% of the Enterococcus spp. (E. faecalis and E. durans) isolated from the hospital wastewater and final effluent wastewater, respectively, were resistant to vancomycin and erythromycin. |
| 14 | Guo et al., 2017 [48] | Sewage | Aerobic-activated sludge (AAS) and an aerobically digested sludge (ADS) | Although MGEs, including plasmids; transposons; integrons (e.g., intI1) and insertion sequences (e.g., ISSsp4, ISMsa21 and ISMba16) were abundant in both the activated and digested sludge. However, distinct microbial populations were associated with the two sledge samples. |
| 15 | Wang JL, et al., 2015 [49] | Sewage | Pharmaceutical WTP (all stages of processing) | The abundance of clinically relevant ARGs, including sul1, sul2, tetO, tetT, tetW and tetM, remained consistently higher throughout the processing stages and discharged into the environment. |
| 16 | Conte D et al., 2017 [50] | Sewage and environmental waters | Hospital effluent, sanitary effluent, different sites within WWTP and upstream and downstream river water | ESBL-producing K. pneumonia and E. coli isolates were higher in hospital effluent and WWTP and river samples, respectively. Quinolone-resistant isolates were identified in hospital effluent, sanitary effluent, outflow sewage and surface water samples. MDR bacteria were detected in the hospital effluent and river waters. |
| 17 | Baumlisberger et al., 2015 [51] | Sewage | Up- and downstream wastewater from nursing home | No obvious difference in ARG and MGE abundances were detected between up- and downstream samples. |
| 18 | Adefisoye et al., 2016 [52] | Sewage | Final effluents of WWTP | MDR E coli isolates associated with neonatal meningitis; intestinal (enterotoxigenic E. coli (ETEC), enteropathogenic E. coli (EPEC) and enteroaggregative E. coli (EAEC)) and ex-intestinal (UPEC) were identified. |
| 19 | Suzuki et al., 2015 [53] | Sewage and environmental waters | Effluents of WWTP and surface water | High levels of ARGs associated with resistance to sulfamethosazole and oxytetracycline were detected in environmental surface water. |
| 20 | Froes AM et al., 2016 [54] | Sewage | Hospital’s wastewater | Diverse ARGs of serine β-lactamases, including uncommon β lactamase genes blaPER, blaVEB and blaGES, were detected in hospital’s wastewater. |
| 21 | Laht M et al., 2014 [55] | Sewage | WWTP | High levels of ARGs associated with resistance to tetracycline, sulfonamide and β-lactam were detected in all stages in WWTP wastewater. No difference in ARGs abundance was identified after the purification process. |
| 22 | Walsh et al., 2011 [56] | Sewage and environmental water | Seepage water, public tap water and control samples: sewage effluent samples from Wales | blaNDM-1-producing bacteria were isolated from 17% (12 out 171) and 4% (2 out 50) of seepage and tap water samples, respectively. The detected strains included 11 species in which blaNDM-1 had not previously been reported (e.g., Shigella boydii and Vibrio cholerae). |
| 23 | Zhang T et al., 2011 [57] | Sewage | Activated sludge of WWTP | Novel plasmids carrying ARGs associated with tetracycline, aminoglycoside and β-lactam resistance were identified. ARGs associated with resistance to tetracycline, macrolide and MDR were highly enriched in the activated sludge. |
| 24 | Chagas TP et al., 2011 [58] | Sewage | Influent, clarifier tank effluent and chlorine tank effluent from hospital STP (sewage treatment plant) | Multiresistant and ESBL-producing bacteria (high resistant rates to amikacin, trimethoprim/sulphametoxazole and cefalothin) were identified in the chlorine contact tank effluent. |
| 25 | Szczepanowski et al., 2009 [59] | Sewage | Activated sludge samples and final effluent of WWTP | Clinically relevant ARGs associated with resistance to several antimicrobial drugs, including aminoglycoside and β-lactam, were identified in activated sludge and effluent wastewater. |
| 26 | Li D et al., 2010 [60] | Sewage and environmental waste water | Wastewater, river water-up (RWU) and -downstream (RWD) associated with oxytetracycline production WWTP | High concentrations of oxytetracycline were identified in wastewater and in river water downstream but not in upstream waters. MDR phenotypes isolates were identified in the WW and RWD and less frequent in RWU. |
| 27 | Zhang H et al., 2019 [61] | Sewage | Samples from 18 WWTPs | Activated sludge was the main reservoir of ARGs associated with resistance to sulfonamide (sul1 and sul2) and tetracycline (tetW, tetX and tetQ). |
| 28 | Li, B et al., 2015 [62] | Sewage, environmental water and faecal samples | Samples from AAS and ADS and different environmental waters and faecal samples (human, chicken and pig) | High level of ARGs, including those associated with MDR, and resistance to bacitracin; tetracycline; β-lactam; macrolide, lincosamide and streptogramin (MLS); aminoglycoside; quinolone and sulphonamide were detected in all samples. |
| 29 | Hembach et al., 2017 [63] | Sewage | Influent and effluent water from seven WWTPs | mcr-1 (associated with resistance to colistin) was detected in influent samples of all seven WWTPs and some of effluent waters (i.e., it was not eliminated during wastewater treatment reaching the aquatic environment). AMR strains of A. baumannii, E. coli and K. pneumonia were detected in both influent and effluents samples. |
| 30 | Igbinosa IH et al., 2012 [64] | Sewage | WWTP | AMR Aeromonas spp. isolates resistant to penicillin, oxacillin, ampicillin and vancomycin were identified. Class A pse1 β-lactamase, class 1 integron and the blaTEM gene were detected in 20.8%, 20.8% and 8.3% of the identified isolates, respectively. |
| 31 | Igbinosa EO et al., 2011 [65] | Sewage | Final effluents WWTP | Vibrio spp. strains (including V. parahaemolyticus, V. fluvialis and V. vulnificus) resistant to erythromycin, chloramphenicol, nitrofurantoin, cefuroxime and cephalothin were detected. SXT antibiotic resistance gene cluster (floR, strB, sul2, dfrA18 and dfrA1) were also identified in % of these strains. |
| 32 | Johnning A, et. Al 2015 [66] | Sewage and environmental water | Up and downstream WWTP and from industrially polluted sites and sediment samples from a pristine lake | Mutations in chromosomal genes gyrA and parC, associated with resistance to fluoroquinolone, were detected in E. coli communities. High abundance of mutations was correlated with the concentration of fluoroquinolones in investigated samples (i.e., samples polluted with high concentrations of fluoroquinolone). |
| 33 | Sahlström L et al., 2009 [67] | Sewage and clinical samples | WWTP and isolates from humans and chickens | Vancomycin-resistant isolates of Enterococcus spp., including E. faecium, E. hirae and E. durans, were detected. |
| 34 | Araújo C et al., 2010 [68] | Sewage | Sludge and sewage of urban and poultry slaughter house WWTP | Vancomycin-resistant isolates of Enterococcus spp., including E. faecium, E. gallinarum and E. casseliflavus, which were also resistant to varied groups of antimicrobial drugs (kanamycin, tetracycline, erythromycin, ciprofloxacin, ampicillin, streptomycin and gentamicin), were detected. |
| 35 | Soge O et al., 2009 [69] | Sewage and environmental water | Water, soil and sewage | The majority of Clostridium perfringens strains recovered from water samples were found to carry more than one ARG encoding resistance to tetracycline and erythromycin. |
| 36 | Zhang X et al., 2009 [70] | Sewage | WWTP | Enterobacteriaceae strains carrying class 1 integrons and ARGs associated with resistance to trimethoprim (dfr17) and streptomycin (aadA5) were detected. |
| 37 | Odjadjare EO et al., 2010 [71] | Sewage | WWTP final effluent, discharge point and upstream and downstream of the discharge point | Most of the Listeria spp. isolates recovered from final effluents were MDR strains. |
| 38 | Okoh AI et al., 2010 [72] | Sewage | WWTP final effluents | MDR Vibrio spp. strains that showed resistance to varied antimicrobial drugs (including sulfamethoxazole, trimethoprim, cotrimoxazole, chloramphenicol, streptomycin, ampicillin, tetracycline, nalidixic acid and gentamicin) were detected. |
| 39 | Yang H et al., 2010 [73] | Sewage and environmental waters | Soil, water and faecal samples | Higher copies of tetracycline ARGs and higher abundance of tetracycline-resistant bacteria were identified in farm (cattle), compared to nonfarm, wastewater samples. |
| 40 | Garcia-Armisen T et al., 2011 [74] | Environmental water | Sewage-contaminated rivers | Most of the ARB detected in the Zenne river, downstream of Brussels, were MDR strains. The abundance of AMR communities (heterotrophic and faecal bacteria) was not correlated with the level of contamination of river water with sewage. |
| 41 | Colomer-Lluch M et al., 2011 [75] | Sewage and environmental water | Urban sewage and river water | β-lactamase genes (blaTEM and blaCTX-M9) and one encoding penicillin-binding protein (mecA) were detected in the DNA phages recovered from all the samples. |
| 42 | Fuentefria DB et al., 2011 [76] | Sewage and environmental water | Hospital wastewater and superficial water | Genetically distinct populations of AMR Pseudomonas aeruginosa were detected in these different environments (hospital wastewater and superficial water that received this wastewater discharge). |
| 43 | Gaze WH et al., 2011 [77] | Sewage | Industrial waste, sewage sludge and pig slurry | Higher prevalence of class 1 integrons was detected in bacteria recovered from sewage sludge and pig slurry (exposed to antibiotic residues and detergents) compared to agricultural soils to which these waste products are amended. It has been estimated that ~1019 bacteria carrying class 1 integrons enter the United Kingdom environment by disposal of sewage sludge each year. |
| 44 | Ma L et al., 2011 [78] | Sewage | WWTP | The abundance of bacteria (E. coli, Klebsiella spp. and Aeromonas veronii) carrying class I integronase gene intI1 were higher in effluent compared to influent wastewater. intI1 was detected in 20.4%, 30.9% and 38.9% of bacteria recovered from influent, activated sludge and effluent wastewater, respectively. This study suggested a role of activated sludge (characterized by high biomass and biodiversity) in developing AMR through the dissemination of integrons. |
| 45 | Mokracka J et al., 2011 [79] | Sewage | WWTP | Quinolone- and fluoroquinolone-resistant strains constituted 56% and 50.4% of recovered integron-bearing E. coli strains (including diarrheagenic and extraintestinal strains carrying virulence traits), respectively. Virulent extraintestinal strains constituted ~50% of all isolates and were detected in samples recovered from all wastewater treatment stages, including final effluent. |
| 46 | Amaya E et al., 2012 [80] | Sewage and environmental waste water | Hospital wastewater and well waters | High levels of MDR E. coli isolates were recovered from samples collected from both hospital wastewaters and environmental well water. E. coli strains harbouring blaCTX-M1 and blaCTX-M9 were predominated in samples collected from wells and hospital wastewater, respectively. |
| 47 | Mokracka J et al., 2012 [81] | Sewage | Municipal WWTP | MDR Enterobacteriaceae strains carrying class 1 and class 2 integrons (12.1%; 221 out of 1832) were identified in different stages of a municipal wastewater treatment plant (61.5%, 12.7% and 25.8% of ARB were originated from raw sewage, aeration tank and final effluent, respectively). The abundance of ARGs and MDR bacteria, particularly the level of ARG diversity and B-lactamase-producers, were higher in final effluent samples. |
| 48 | Splindler A et al., 2012 [82] | Sewage | Untreated hospital effluents | Half of Pseudomonas spp. isolates recovered from untreated hospital effluent wastewater were MDR strains, while 41.9% (52 out of 124) of the isolates were found to carry intlI. |
| 49 | Gundogdu, A. et al., 2012 [83] | Sewage | Untreated hospital wastewaters and WWTP | High level of ESBL-producing E. coli isolates were detected in untreated hospital wastewaters (blaSHV), with distinct genotypes (blaCTX-M) associated with the samples recovered from WWTP. |
| 50 | Zarfel, G et al., 2013 [84] | Sewage and clinical samples | Sewage and human urinary tract infection samples | ESBL-producing bacteria carrying blaCTX-M were predominated in both sewage sludge (blaCTX-M-15) and UTI (blaCTX-M-1) samples. The study suggested the occurrence of a genetic exchange between the ESBL-resistant E. coli populations from human infections and those present in sewage sludge. |
| 51 | Colomer-Lluch M et al., 2013 [85] | Sewage and environmental water | Sewage and river water samples | Quinolone-resistant E. coli strains of clinically relevant ST69 and ST131 (carrying virulence traits) predominated in samples recovered from urban wastewater and both river and wastewaters, respectively. Similar virulence and macro-restriction profiles were identified in environmental and human isolates of ST131. |
| 52 | Sadowy E et al., 2014 [86] | Sewage and environmental water | Wastewater, riverine estuary and anthropogenically impacted marine catchment basin | AMR isolates of Enterococcus spp., especially fluoroquinolone- and aminoglycoside-resistant E. faecium that shared virulence determinants and ST similar to nosocomial high-risk enterococcal clonal complexes (HiRECC), were detected. |
| 53 | Gao P et al., 2015 [87] | Sewage | WWTP | Positive correlations were observed between the occurrence of heavy metals (e.g., zinc and lead and ereB, mefA&E and ermB) and antibacterial residues (e.g., triclosan with ereA, ereB, mefA&E and ermB) in urban wastewaters and the abundance of erythromycin-resistant genes. |
| 54 | Nishiyama M et al., 2015 [88] | Sewage and environmental water | Sewage and urban river water samples | vanC-type vancomycin-resistant E. faecium and E. faecalis, which are the major types of enterococci in humans, were detected in both sewage and urban river water samples. |
| 55 | Zhang S et al., 2015 [89] | Sewage | WWTP | Gram-negative and -positive isolates dominated WWTP influent and effluent samples, respectively. The frequency of detection of tetracycline-, sulphonamide-, streptomycin- and β-lactam-resistance genes (except sulA and blaCTX-M) were higher in ARB from influent compared to effluent samples. The abundances of ARGs in activated sludge were higher in aerobic compartments than in anoxic ones. |
| 56 | Simo Tchuinte PL et al., [90] | Sewage | Hospital effluent and sludge | Novel class 3 integrons with oxacillinase gene cassette, including aminoglycoside and β-lactam-resistant genes (blaOXA-10, blaOXA-368 or blaOXA-2), were identified in Acinetobacter johnsonii, Aeromonas allosaccharophila and Citrobacter freundii, which were recovered from hospital effluent samples. |
| 57 | Young S et al., 2016 [91] | Sewage and environment water | Water and sediment samples from sewage spill site | Nosocomial pathogen; vancomycin-resistant E. faecium (harbouring vanA associated with a high resistance level) were isolated from water and sediment for up to 3 days after a sewage spill. vanA gene were found to persist for an additional week within these environments. Culturable levels of enterococci in water exceeded recreational water guidelines for 2 weeks following the spill, declining about five orders of magnitude in sediments and two orders of magnitude in the water column over 6 weeks. |
| 58 | Lee J et al., 2017 [92] | Sewage | Food waste-recycling wastewater (FRW), manure and sewage sludge | The abundance of ARGs was greatest in manure, followed by sewage sludge and FRW. However, different patterns in the diversity and mechanisms of ARGs were identified. ARG associated with β-lactam resistance were higher in the FRW, and sulfonamides-resistant genes are higher in sludge. Total ARGs is associated with class 1 integron only in manure and sludge. |
| 59 | An XL et al., 2018 [93] | Sewage | Influent, activated sludge and effluents of urban WWTP | High concentration of class 1 integron gene cassette (including trimethoprim, aminoglycoside and beta-lactam resistance genes) were identified in activated sludge. |
| 60 | Haller L et al., 2018 [94] | Sewage | Hospital effluents | MDR bacteria belonging to Enterobacteriaceae and other species, including ESBL- and carbapenemase-producers, were identified. |
| 61 | Galler H et al.,2018 [95] | Sewage | Activated sludge | Clinically relevant ARBs, including ESBL-Enterobacteriaceae, MRSA and vancomycin-resistant Enterococcus spp., were detected. ARG associated with resistance to β-lactam, vancomycin (vanA) and methicilin (mecA) were identified. |
| 62 | Quach-Cu J et al., 2018 [96] | Sewage | Raw wastewater, activated sludge and secondary and tertiary WWTP effluent | The abundance of blaSHV, blaTEM and sul1 were higher in raw wastewater than other samples. |
| 63 | Yousfi K et al., 2019 [97] | Sewage | Hospital effluents | Enterobacteriaceae isolates (including E. coli and K. pneumoniae) and non-Enterobacteriaceae Gram-negative bacterial isolates (including A. baumannii and A. hydrophila) showed high levels of resistance to β-lactam and non-β-lactam-antibiotics, and most of them are multidrug-resistant. This study is the first study that found genes encoding carbapenemases, including blaOXA-23 and blaOXA-48, like in A. baumannii, K. oxytoca and S. xiamenensis in Algerian hospital effluents. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fouz, N.; Pangesti, K.N.A.; Yasir, M.; Al-Malki, A.L.; Azhar, E.I.; Hill-Cawthorne, G.A.; Abd El Ghany, M. The Contribution of Wastewater to the Transmission of Antimicrobial Resistance in the Environment: Implications of Mass Gathering Settings. Trop. Med. Infect. Dis. 2020, 5, 33. https://doi.org/10.3390/tropicalmed5010033
Fouz N, Pangesti KNA, Yasir M, Al-Malki AL, Azhar EI, Hill-Cawthorne GA, Abd El Ghany M. The Contribution of Wastewater to the Transmission of Antimicrobial Resistance in the Environment: Implications of Mass Gathering Settings. Tropical Medicine and Infectious Disease. 2020; 5(1):33. https://doi.org/10.3390/tropicalmed5010033
Chicago/Turabian StyleFouz, Nour, Krisna N. A. Pangesti, Muhammad Yasir, Abdulrahman L. Al-Malki, Esam I. Azhar, Grant A. Hill-Cawthorne, and Moataz Abd El Ghany. 2020. "The Contribution of Wastewater to the Transmission of Antimicrobial Resistance in the Environment: Implications of Mass Gathering Settings" Tropical Medicine and Infectious Disease 5, no. 1: 33. https://doi.org/10.3390/tropicalmed5010033