
A Poorly Known Catfish Clade in an Endangered Neotropical Biodiversity Hotspot: Relationships and Distribution Patterns of the Cambeva variegata Group (Siluriformes: Trichomycteridae)
 , , , ,
, , , ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
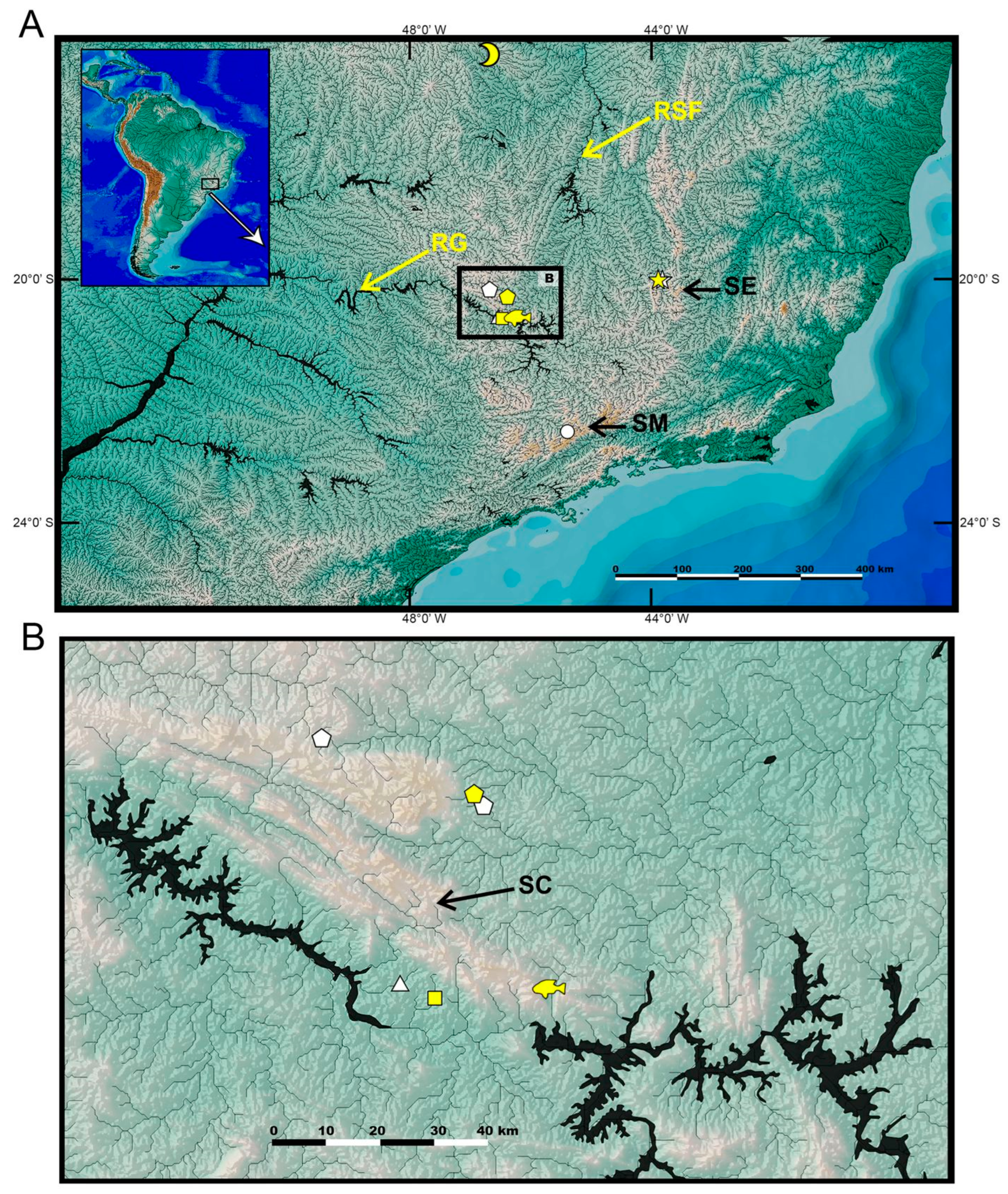
2.1. Specimens
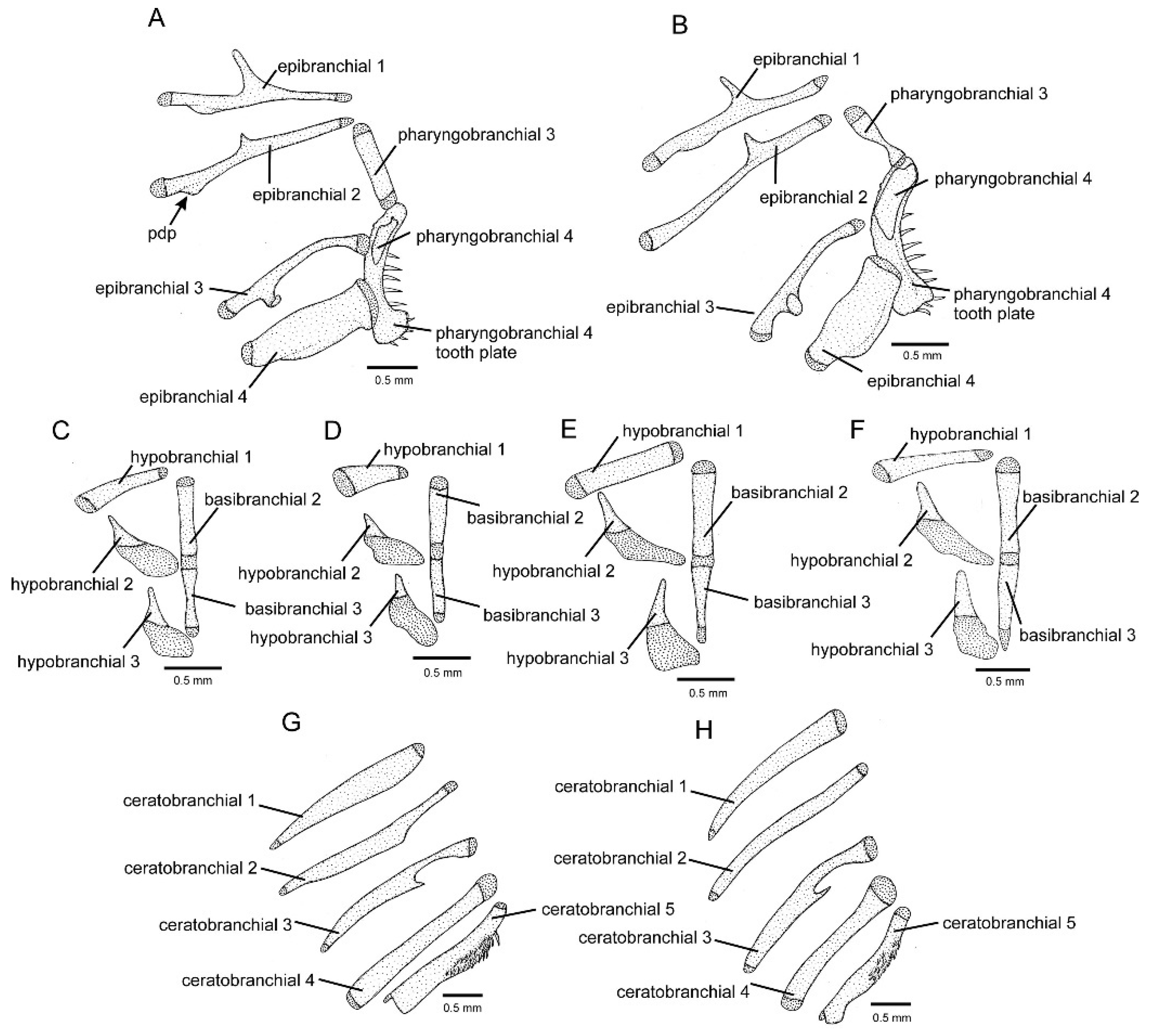
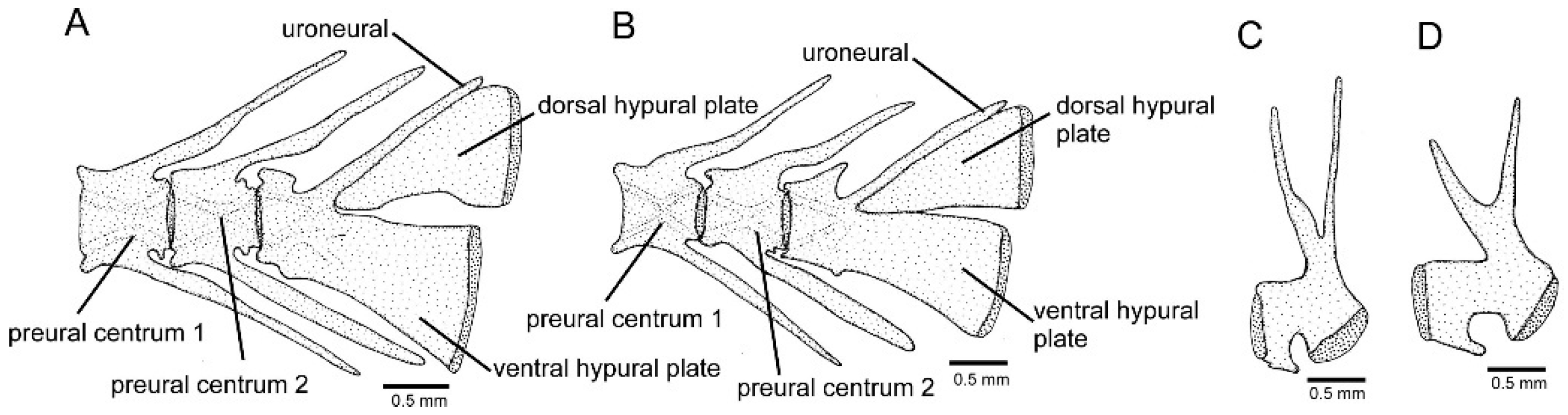
2.2. Morphological Data
2.3. DNA Extraction, Amplification, and Sequencing
2.4. Phylogenetic Analysis
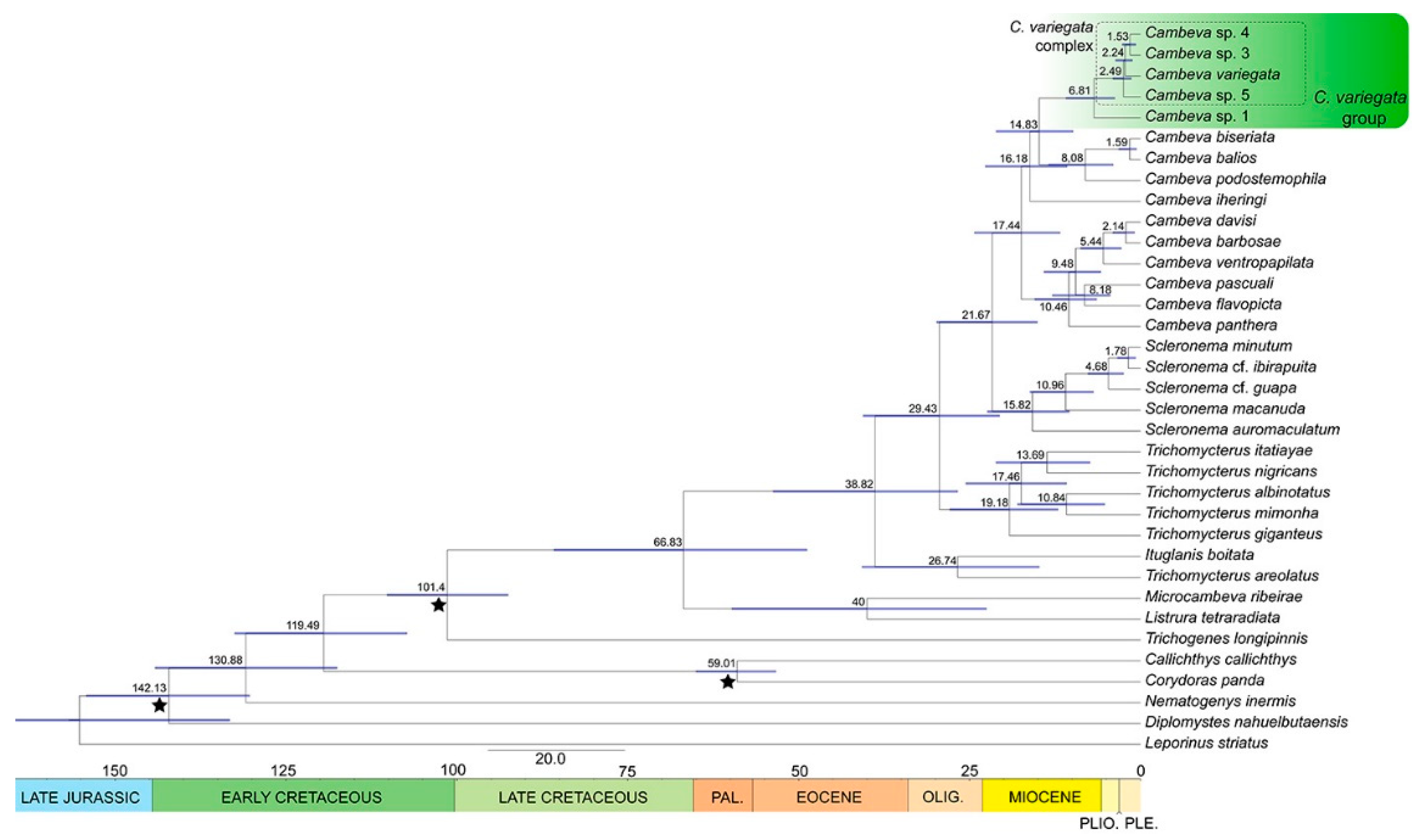
2.5. Divergence Time Estimation
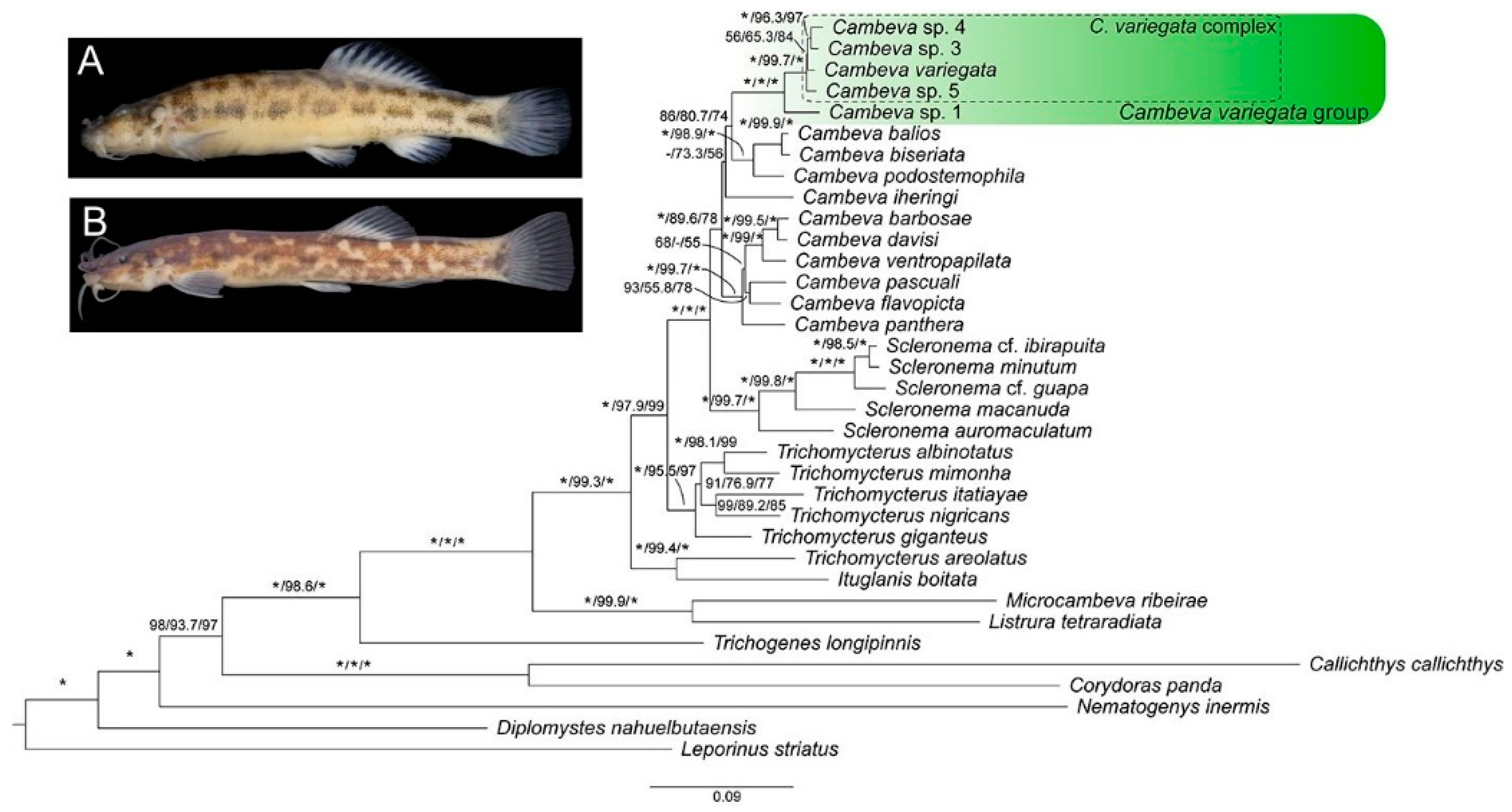
3. Results
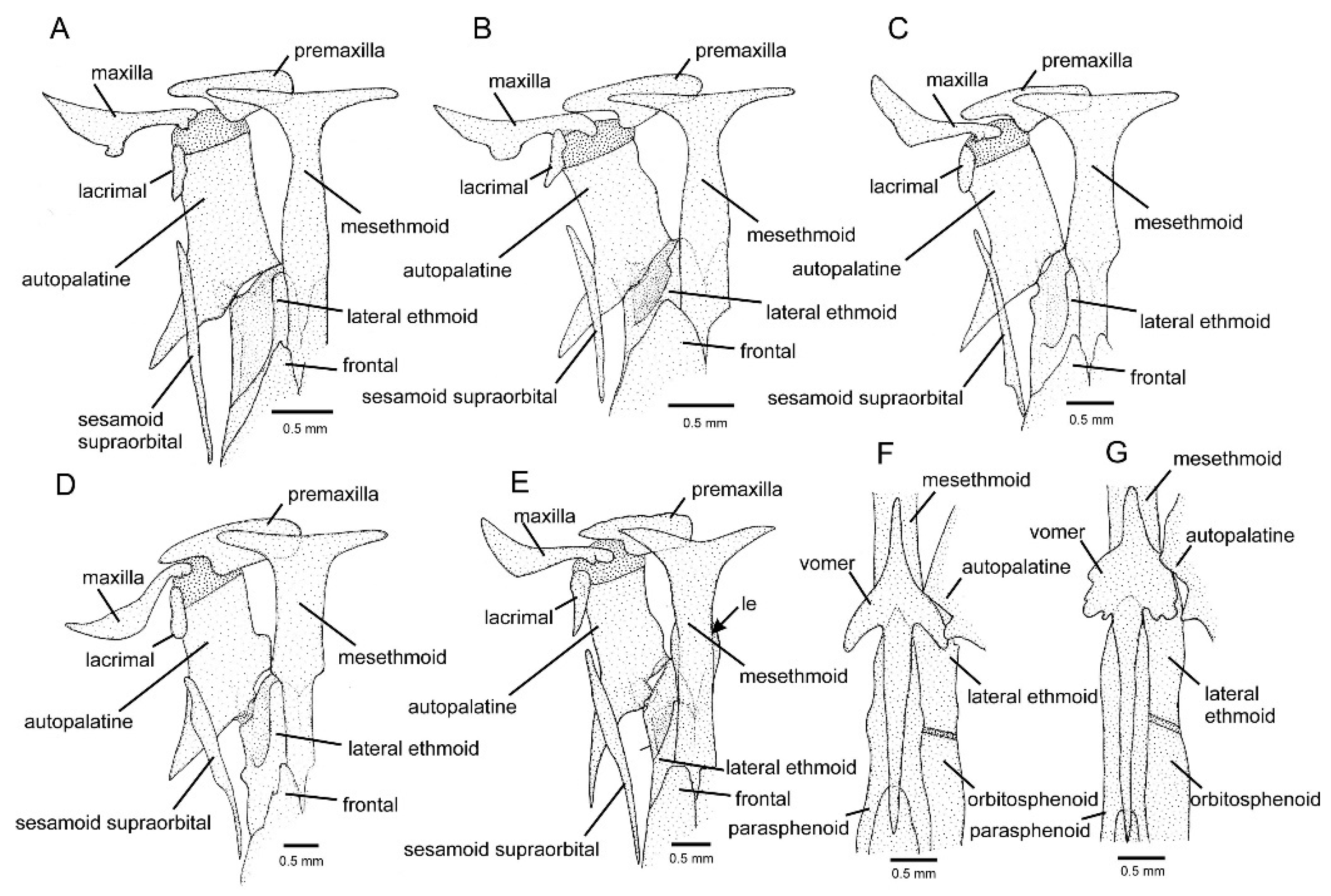
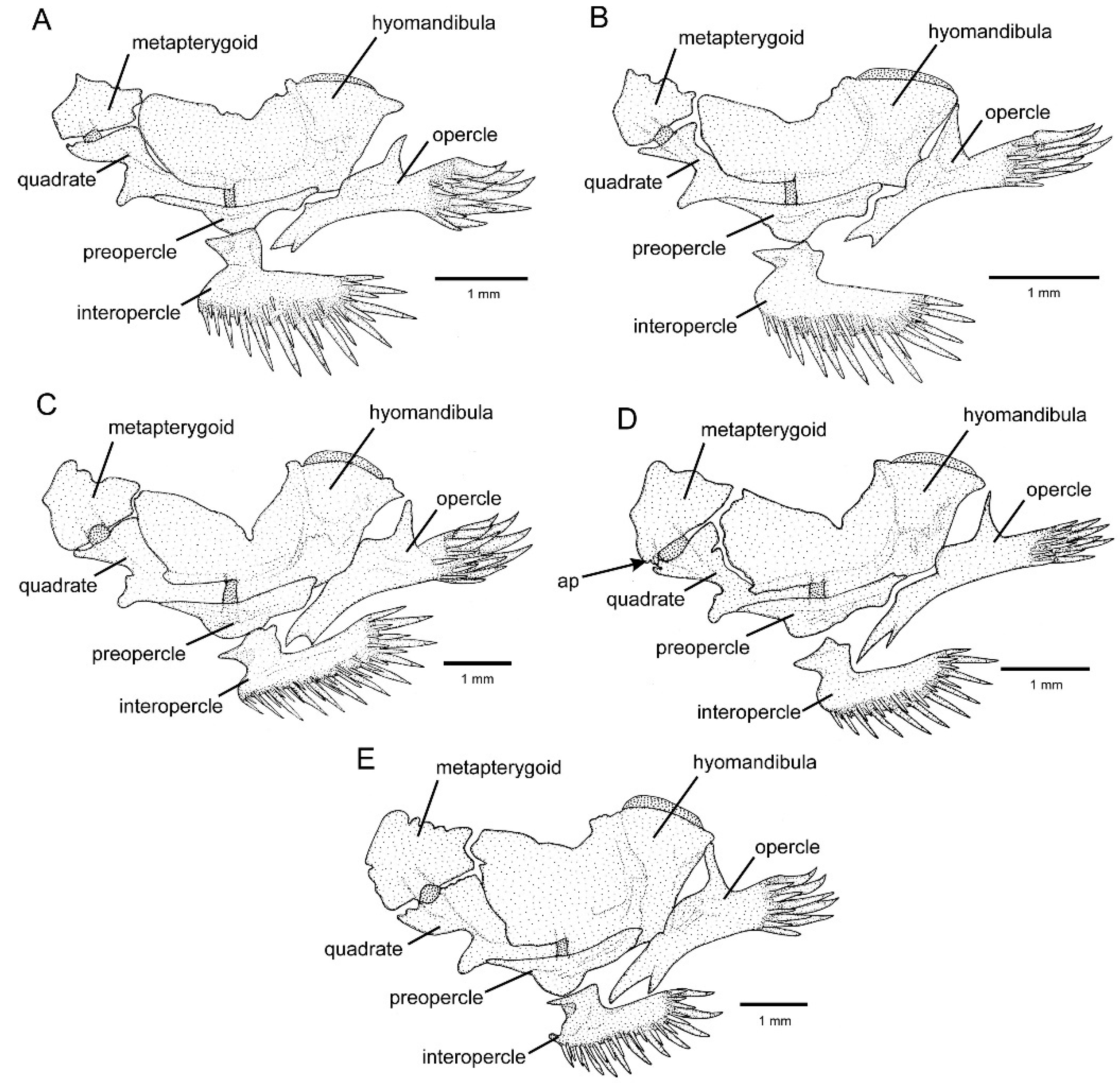
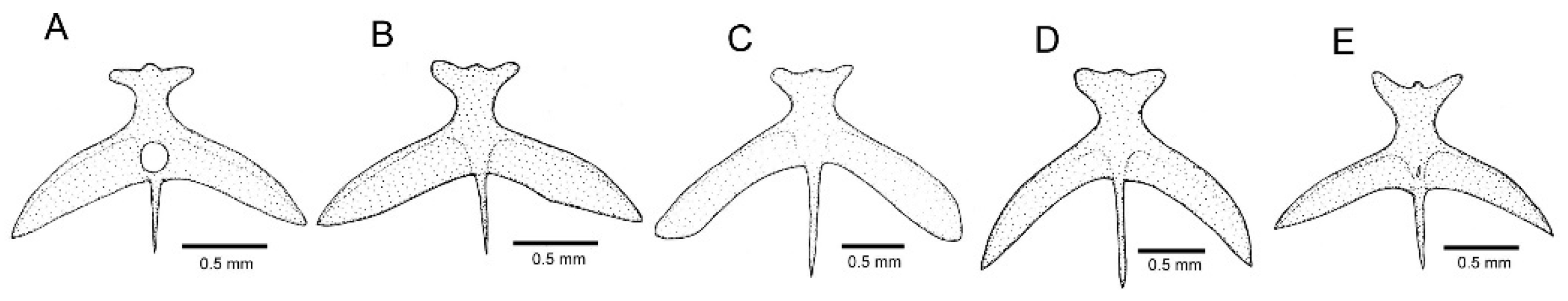
3.1. Phylogenetic Relationships and Comparative Morphology
3.2. Time-Calibrated Phylogeny
4. Discussion
4.1. Phylogenetic Relationships
4.2. Distribution Patterns
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| COI | ND4 | tRNA Ser His Leu | CYTB | RAG2 | |
|---|---|---|---|---|---|
| Leporinus striatus | JN989019.1 | — | — | EU183020.1 | AY804096.1 |
| Diplomystes nahuelbutaensis | AP012011.1 | NC015823.1 | NC015823.1 | MN640590 | DQ492317 |
| Nematogenys inermis | EU359428 | AY307250.1 | AY307250.1 | — | KY858182.1 |
| Corydoras panda | NC049097.1 | GU210065.1 | GU210065.1 | NC049097.1 | KP960362.1 |
| Callichthys callichthys | MZ051783.1 | AY307241.1 | AY307241.1 | KP960058 | DQ492324 |
| Trichogenes longipinnis | OQ810037 | MN389484 | MN389484 | MK123704 | MF431117 |
| Microcambeva ribeirae | MN385807.1 | MN389502.1 | MN389502.1 | OK334290 | MN385832 |
| Listrura tetraradiata | JQ231083 | MN389497 | MN389497.1 | JQ231088.1 | MN385826.1 |
| Trichomycterus areolatus | AP012026.1 | AP012026.1 | AP012026.1 | FJ772214 | KY858188 |
| Ituglanis boitata | OQ810038 | MN389485.1 | MN389485.1 | MK123706 | MK123758 |
| Trichomycterus itatiayae | MW671552 | OR948809 | — | MW679291 | OL779233 |
| Trichomycterus nigricans | MN813005 | MN389488.1 | MN389488.1 | MK123723 | MK123765 |
| Trichomycterus albinotatus | MN813007 | OM324337.1 | OM324337.1 | MK123716 | MN812990 |
| Trichomycterus mimonha | MW196749 | OM324343.1 | OM324343.1 | MW196758 | MW196783 |
| Trichomycterus giganteus | MT470413.1 | PP333226 | PP336672 | MK123720.1 | MT446426.1 |
| Scleronema minutum | MK123685 | MN389486.1 | MN389486.1 | MK123707 | MK123759.1 |
| Scleronema cf. guapa | PP319012 | PP333227 | PP336673 | MK123709.1 | MF431118.1 |
| Scleronema cf. ibirapuita | MK123688.1 | PP333228 | PP336674 | MK123710.1 | MK123761.1 |
| Scleronema macanuda | MK123686.1 | PP333229 | PP336675 | MK123708.1 | MK123760.1 |
| Scleronema auromaculatum | OM037445.1 | — | — | OM037134.1 | OM037136.1 |
| Cambeva barbosae | MK123689.1 | MN389487.1 | MN389487.1 | OQ110808 | OQ110815.1 |
| Cambeva balios | OQ810040 | PP333230 | PP336676 | OQ814186 | OQ814193 |
| Cambeva pascuali | MF034463 | PP333231 | PP336677 | OQ110811 | OQ110820 |
| Cambeva panthera | OQ810041 | PP333232 | PP336678 | OQ814187 | OQ814194 |
| Cambeva flavopicta | OQ810042 | PP333233 | PP336679 | OQ814188 | OQ814195 |
| Cambeva davisi | PP319014 | PP333234 | PP336680 | MK123714 | MK123762 |
| Cambeva biseriata | PP319015 | PP333235 | PP336681 | OQ110806 | OQ110817 |
| Camveba ventropapilata | PP319016 | PP333236 | PP336682 | OQ110807 | OQ110818 |
| Cambeva iheringi | GU701893 | — | — | KY858074 | KY858223 |
| Cambeva sp. 1 | PP319017 | PP333237 | PP336683 | PP328532 | PP333215 |
| Cambeva sp. 5 | PP319018 | PP333238 | PP336684 | PP328533 | PP333216 |
| Cambeva variegata | PP319019 | PP333239 | PP336685 | PP328534 | PP333217 |
| Cambeva sp. 3 | PP319020 | PP333240 | PP336686 | PP328535 | PP333218 |
| Cambeva sp. 4 | PP319021 | PP333241 | PP336687 | PP328536 | PP333219 |
| Cambeva podostemophila | OQ810043 | — | — | OQ814189 | OQ814196 |
Appendix B
| Partition | Base Pairs | Evolutive Model |
|---|---|---|
| COI 1st | 244 | TRN+I+G |
| COI 2nd | 244 | HKY+I |
| COI 3rd | 244 | TRN+I+G |
| ND4 1st | 231 | GTR+I+G |
| ND4 2nd, CYTB 1nd | 562 | TRN+I+G |
| ND4 3rd | 231 | GTR+G |
| tRNA His Ser Leu | 162 | GTR+G |
| CYTB 1st | 331 | TRN+I+G |
| CYTB 3rd | 331 | GTR+I+G |
| RAG2 1st | 263 | HKY+G |
| RAG2 2nd | 263 | GTR+I |
| RAG2 3rd | 263 | K80+I |
References
- Costa, W.J.E.M.; Azevedo-Santos, V.M.; Mattos, J.L.O.; Katz, A.M. Molecular phylogeny, taxonomy and distribution patterns of trichomycterine catfishes in the middle Rio Grande drainage, south-eastern Brazil (Siluriformes: Trichomycteridae). Fishes 2023, 8, 206. [Google Scholar] [CrossRef]
- Katz, A.M.; Barbosa, M.A.; Mattos, J.L.O.; Costa, W.J.E.M. Multigene analysis of the catfish genus Trichomycterus and description of a new South American trichomycterine genus (Siluriformes, Trichomycteridae). Zoosyst. Evol. 2018, 94, 557–566. [Google Scholar] [CrossRef]
- Costa, W.J.E.M. Description de huit nouvelles espèces du genre Trichomycterus (Siluriformes: Trichomycteridae), du Brésil oriental. Rev. Franç. d’aquariol. Herpetol. 1992, 18, 101–110. [Google Scholar]
- Ferrer, J.; Malabarba, L.R. Systematic revision of the Neotropical catfish genus Scleronema (Siluriformes: Trichomycteridae), with descriptions of six new species from Pampa grasslands. Neotrop. Ichthyol. 2020, 18, e190081. [Google Scholar] [CrossRef]
- Costa, W.J.E.M.; Sampaio, W.M.S.; Giongo, P.; de Almeida, F.B.; Azevedo-Santos, V.M.; Katz, A.M. An enigmatic interstitial trichomycterine catfish from south-eastern Brazil found at about 1000 km away from its sister group (Siluriformes: Trichomycteridae). Zool. Anz. 2022, 297, 85–96. [Google Scholar] [CrossRef]
- Bockmann, F.A.; Ferrer, J.; Rizzato, P.P.; Esguícero, A.L.H.; Duboc, L.F.; Ingenito, L.F.S. Anatomy, ecology, and behavior of a new species of Scleronema Eigenmann, 1917 (Siluriformes: Trichomycteridae) from coastal drainages in the southern Brazilian Atlantic Rainforest, with comments on the monophyly and phylogeny of the genus. Zootaxa 2023, 5297, 1–47. [Google Scholar] [CrossRef] [PubMed]
- de Pinna, M.C.C. A new subfamily of Trichomycteridae (Teleostei, Siluriformes), lower locarioid relationships and a discussion on the impact of additional taxa for phylogenetic analysis. Zool. J. Linn. Soc. 1992, 106, 175–229. [Google Scholar] [CrossRef]
- Thereza, M.R.; Langeani, F. Bagres e Cascudos do Rio Grande, Alto Rio Paraná; Editora CRV: Curitiba, Brazil, 2019; 122p. [Google Scholar]
- Ochoa, L.E.; Roxo, F.F.; DoNascimiento, C.; Sabaj, M.H.; Datovo, A.; Alfaro, M.; Oliveira, C. Multilocus analysis of the catfish family Trichomycteridae (Teleostei: Ostariophysi: Siluriformes) supporting a monophyletic Trichomycterinae. Mol. Phyl. Evol. 2017, 115, 71–81. [Google Scholar] [CrossRef]
- Costa, W.J.E.M.; Feltrin, C.R.M.; Mattos, J.L.M.; Amorim, P.F.A.; Katz, A.M. Phylogenetic relationships of new taxa support repeated pelvic fin loss in mountain catfishes from southern Brazil (Siluriformes: Trichomycteridae). Zool. Anz. 2023, 305, 82–90. [Google Scholar] [CrossRef]
- Myers, N.; Mittermeir, R.A.; Mittermeir, C.G.; da Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef]
- Leary, S.; Underwood, W.; Anthony, R.; Cartner, S.; Corey, D.; Grandin, T.; Greenacre, C.; Gwaltney-Brant, S.; McCrackin, M.; Meyer, R.; et al. AVMA Guidelines for the Euthanasia of Animals: 2020 Edition. 2020. Available online: http://www.avma.org/sites/default/files/2020-02/Guidelines-on-Euthanasia-2020.pdf (accessed on 3 December 2022).
- Taylor, W.R.; Van Dyke, G.C. Revised procedures for staining and clearing small fishes and other vertebrates for bone and cartilage study. Cybium 1985, 9, 107–119. [Google Scholar]
- Costa, W.J.E.M. Comparative osteology, phylogeny and classification of the eastern South American catfish genus Trichomycterus (Siluriformes: Trichomycteridae). Taxonomy 2021, 1, 160–191. [Google Scholar] [CrossRef]
- Kubicek, K.M. Developmental osteology of Ictalurus punctatus and Noturus gyrinus (Siluriformes: Ictaluridae) with a discussion of siluriform bone homologies. Verteb. Zool. 2022, 72, 661–727. [Google Scholar] [CrossRef]
- Arratia, G.; Huaquin, L. Morphology of the lateral line system and of the skin of diplomystid and certain primitive loricarioid catfishes and systematic and ecological considerations. Bonn. Zool. Monogr. 1995, 36, 1–110. [Google Scholar]
- Bockmann, F.A.; Sazima, I. Trichomycterus maracaya, a new catfish from the upper rio Paraná, southeastern Brazil (Siluriformes: Trichomycteridae), with notes on the T. brasiliensis species-complex. Neotrop. Ichthyol. 2004, 2, 61–74. [Google Scholar] [CrossRef]
- Villa-Verde, L.; Lazzarotto, H.; Lima, S.Q.M. A new glanapterygine catfish of the genus Listrura (Siluriformes: Trichomycteridae) from southeastern Brazil, corroborated by morphological and molecular data. Neotrop. Ichthyol. 2012, 10, 527–538. [Google Scholar] [CrossRef]
- Ward, R.D.; Zemlak, T.S.; Innes, B.H.; Last, P.R.; Hebert, P.D. DNA barcoding Australia’s fish species. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2005, 360, 1847–1857. [Google Scholar] [CrossRef] [PubMed]
- Shimabukuro-Dias, C.K.; Oliveira, C.; Reis, R.E.; Foresti, F. Molecular phylogeny of the armored catfish family Callichthyidae (Ostariophysi, Siluriformes). Mol. Phylog. Evol. 2004, 32, 152–163. [Google Scholar] [CrossRef]
- Costa, W.J.E.M.; Henschel, E.; Katz, A.M. Multigene phylogeny reveals convergent evolution in small interstitial catfishes from the Amazon and Atlantic forests (Siluriformes: Trichomycteridae). Zool. Scr. 2020, 49, 159–173. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Chenna, R.; Sugawara, H.; Koike, T.; Lopez, R.; Gibson, T.J.; Higgins, D.G.; Thompson, J.D. Multiple sequence alignment with the Clustal series of programs. Nucleic Acids Res. 2003, 31, 3497–3500. [Google Scholar] [CrossRef]
- Lanfear, R.; Frandsen, P.B.; Wright, A.M.; Senfeld, T.; Calcott, B. PartitionFinder 2: New methods for selecting partitioned models of evolution for molecular and morphological phylogenetic analyses. Mol. Biol. Evol. 2017, 34, 772–773. [Google Scholar] [CrossRef]
- Suchard, M.A.; Lemey, P.; Baele, G.; Ayres, D.L.; Drummond, A.J.; Rambaut, A. Bayesian phylogenetic and phylodynamic data integration using BEAST 1.10. Vir. Evol. 2018, 4, vey016. [Google Scholar] [CrossRef]
- Gernhard, T. The conditioned reconstruction process. J. Theor. Biol. 2008, 253, 769–778. [Google Scholar] [CrossRef]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarisation in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [PubMed]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New models and efficient methods for phylogenetic inference in the genomic era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [PubMed]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the ultrafast bootstrap approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef] [PubMed]
- Guindon, S.; Dufayard, J.F.; Lefor, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [PubMed]
- Lundberg, J.G.; Sullivan, J.P.; Rodiles-Hernández, R.; Hendrickson, D.A. Discovery of African roots for the Mesoamerican Chiapas catfish, Lacantunia enigmatica, requires an ancient intercontinental passage. Proc. Acad. Nat. Sci. Philad. 2007, 156, 39–53. [Google Scholar] [CrossRef]
- Betancur, R.-R.; Órti, G.; Pyron, R.A. Fossil-based comparative analyses reveal ancient marine ancestry erased by extinction in ray-finned fishes. Ecol. Lett. 2015, 18, 441–450. [Google Scholar] [CrossRef] [PubMed]
- Ochoa, L.E.; Datovo, A.; DoNascimiento, C.; Roxo, F.F.; Sabaj, M.H.; Chang, J.; Melo, B.F.; Silva, G.S.C.; Foresti, F.; Alfaro, M.; et al. Phylogenomic analysis of trichomycterid catfshes (Teleostei: Siluriformes) inferred from ultraconserved elements. Sci. Rep. 2020, 10, 2697. [Google Scholar] [CrossRef]
- Riccomini, C.; Sant’anna, L.G.; Ferrari, A.L. Evolução geológica do Rift Continental do Sudeste do Brasil. In Geologia do Continente Sul-Americano: Evolução da obra de Fernando Flávio Marques de Almeida; Mantesso-Neto, V., Bartorelli, A., Carneiro, C.D.R., Brito-Neves, B.B., Eds.; Beca: São Paulo, Brazil, 2004; pp. 383–405. [Google Scholar]
- Rezende, E.A.; Salgado, A.A.R.; Castro, P.T.A. Evolução da rede de drenagem e evidências de antigas conexões entre as bacias dos rios Grande e São Francisco no sudeste brasileiro. Ver. Brasil. Geomorfol. 2018, 19, 483–501. [Google Scholar] [CrossRef]
- Vilardo, P.J.; Katz, A.M.; Costa, W.J.E.M. Phylogeny and historical biogeography of neotropical catfishes Trichomycterus (Siluriformes: Trichomycteridae) from eastern Brazil. Mol. Phylog. Evol. 2023, 186, 107836. [Google Scholar] [CrossRef]
- Moreira Filho, O.; Buckup, P.A. A poorly known case of watershed transposition between the São Francisco and upper Paraná river basins. Neotrop. Ichthyol. 2005, 3, 449–452. [Google Scholar] [CrossRef]
- Giulietti, A.M.; Pirani, J.R.; Harley, R.M. Espinhaço Range region, eastern Brazil. In Centres of Plant Diversity: A Guide and Strategy for Their Conservation; Davis, S.D., Heywood, V.H., Herrera-Macbryde, O., Villa-Lobos, J., Hamilton, A.C., Eds.; IUCN Publication Unit: Cambridge, UK, 1997; pp. 397–404. [Google Scholar]
- Gonzaga, D.R.; Peixoto, A.L.; Neto, L.M. Patterns of richness and distribution of Cactaceae in the Serra da Mantiqueira, Southeast Brazil, and implications for its conservation. Acta Bot. Bras. 2018, 33, 1–9. [Google Scholar] [CrossRef]
- Lütken, C.F. Siluridae novae Brasiliae centralis a clarissimo J. Reinhardt in provincia Minas-geraës circa oppidulum Lagoa Santa, praecipue in flumine Rio das Velhas et affluentibus collectae, secundum characteres essentiales breviter descriptae. Overs. Kongel. Danske Vidensk. Selsk. Forhandl. Kjobenhavn 1874, 1873, 29–36. [Google Scholar]
- Eigenmann, C.H.; Eigenmann, R.S. Preliminary notes on South American Nematognathi, II. Proc. Calif. Acad. Sci. 1889, 2, 28–56. [Google Scholar] [CrossRef]
- Miranda Ribeiro, A. Fauna brasiliense, peixes, tomo 4. Arquiv. Mus. Nac. 1912, 16, 1–504. [Google Scholar]
- Eigenmann, C.H. The Pygidiidae, a family of South American catfishes. Mem. Carnegie Mus. 1918, 7, 259–398. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Costa, W.J.E.M.; Mattos, J.L.O.; Azevedo-Santos, V.M.; Feltrin, C.R.M.; Amorim, P.F.; Ottoni, F.P.; Vilardo, P.J.; Katz, A.M. A Poorly Known Catfish Clade in an Endangered Neotropical Biodiversity Hotspot: Relationships and Distribution Patterns of the Cambeva variegata Group (Siluriformes: Trichomycteridae). Fishes 2024, 9, 116. https://doi.org/10.3390/fishes9040116
Costa WJEM, Mattos JLO, Azevedo-Santos VM, Feltrin CRM, Amorim PF, Ottoni FP, Vilardo PJ, Katz AM. A Poorly Known Catfish Clade in an Endangered Neotropical Biodiversity Hotspot: Relationships and Distribution Patterns of the Cambeva variegata Group (Siluriformes: Trichomycteridae). Fishes. 2024; 9(4):116. https://doi.org/10.3390/fishes9040116
Chicago/Turabian StyleCosta, Wilson J. E. M., José Leonardo O. Mattos, Valter M. Azevedo-Santos, Caio R. M. Feltrin, Pedro F. Amorim, Felipe P. Ottoni, Paulo J. Vilardo, and Axel M. Katz. 2024. "A Poorly Known Catfish Clade in an Endangered Neotropical Biodiversity Hotspot: Relationships and Distribution Patterns of the Cambeva variegata Group (Siluriformes: Trichomycteridae)" Fishes 9, no. 4: 116. https://doi.org/10.3390/fishes9040116