
Age, Growth, and Reproductive Biology of Euchiloglanis davidi in the Middle and Lower Yalong River, China
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Age Determination
2.3. Length–Weight Relationship
2.4. Growth Parameters
2.5. Fecundity and Oocyte Diameter
2.6. Statistical Analyses
3. Results
3.1. Comparison of Calcified Structures
3.2. Length–Frequency Distribution and SL–W Relationship
3.3. Age and Growth
3.4. Gonad Development and Secondary Sex Characteristics
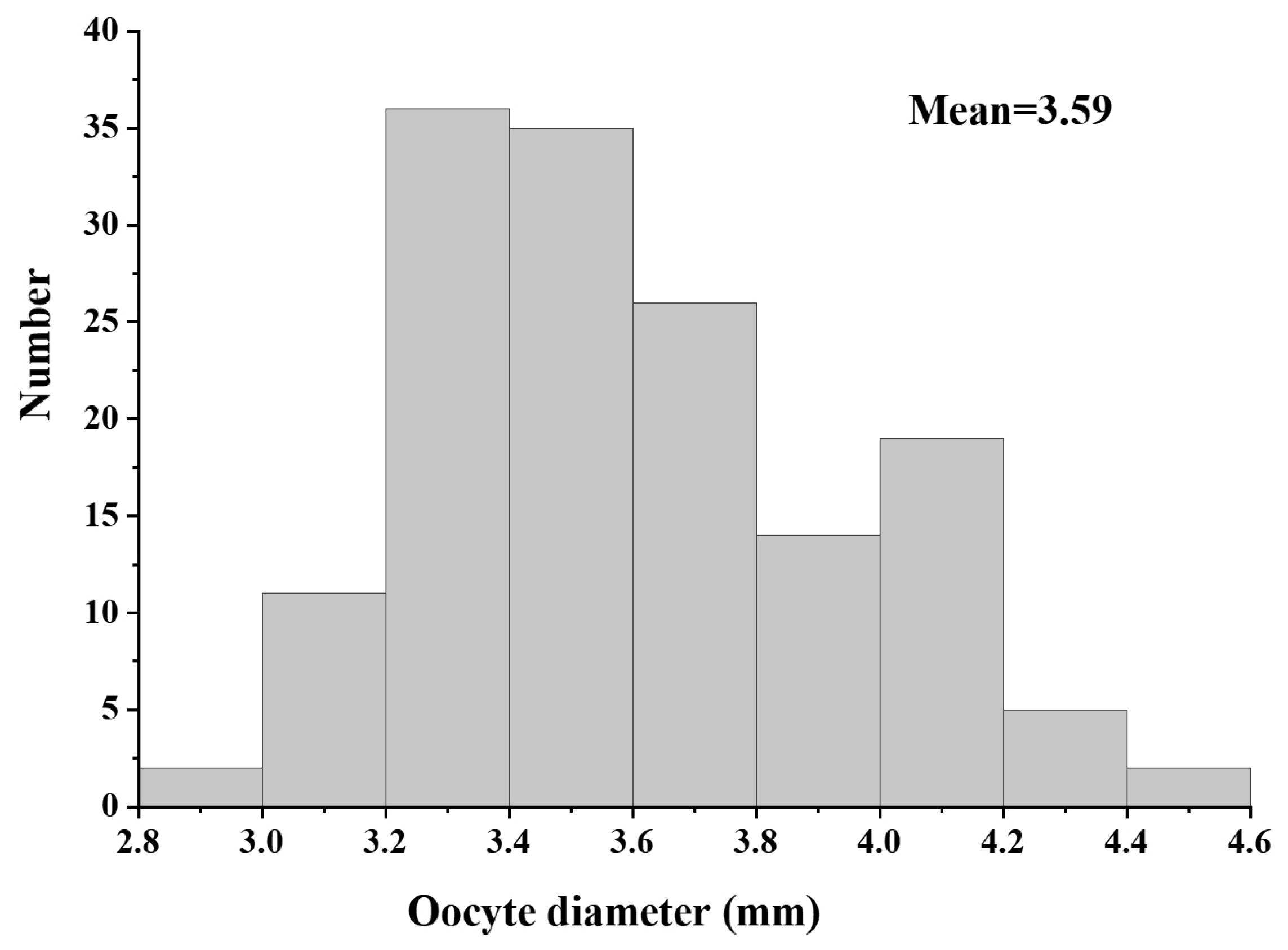
3.5. Oocyte Diameter and Sexual Maturity
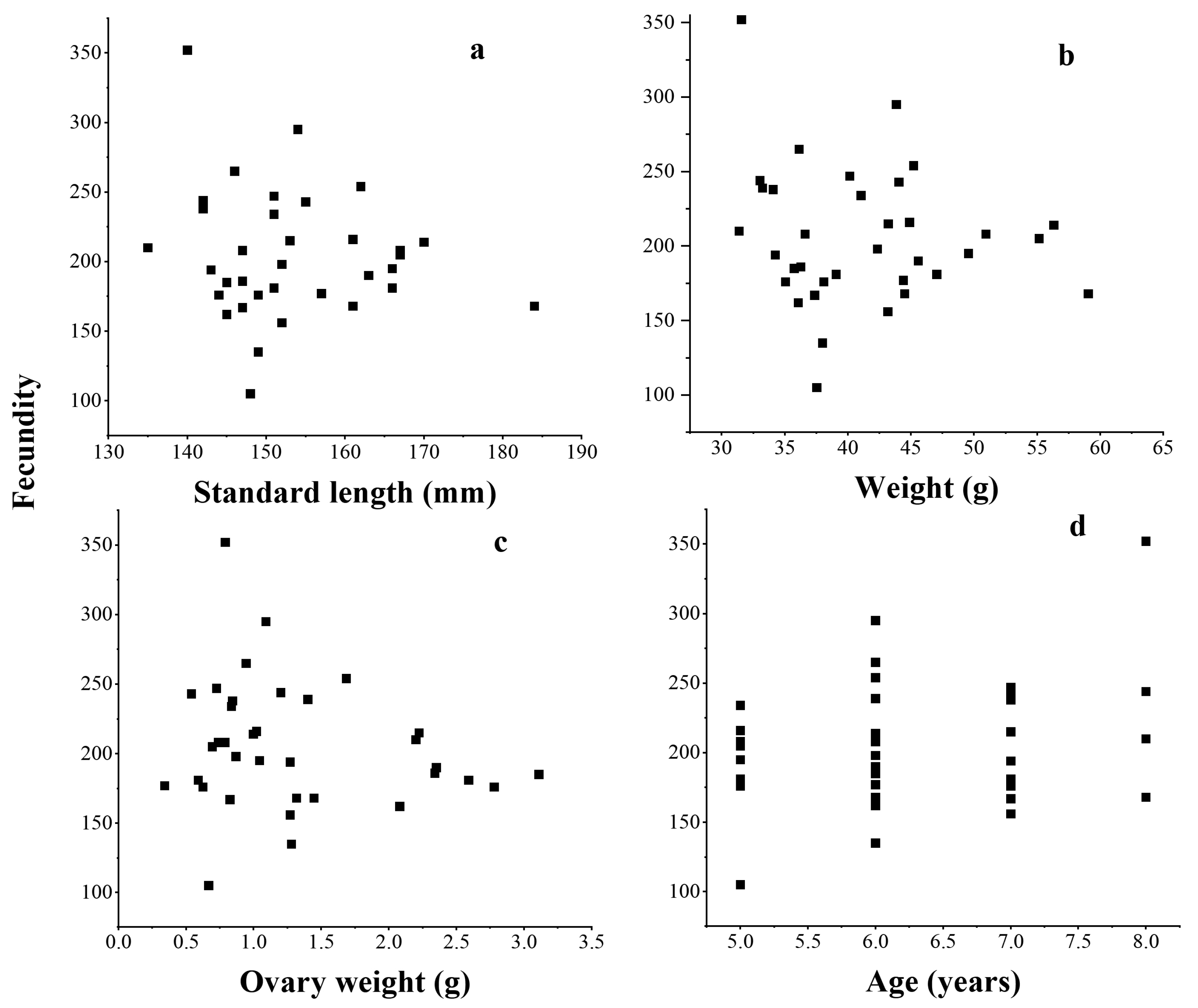
3.6. Fecundity
4. Discussion
4.1. Comparison of Calcified Structures
4.2. Growth Characteristics
4.3. Fecundity
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Feng, J.; Yang, D.; Qin, Z.; Jiang, B.; Pan, Y. Plasmic biochemistry indicators, classification of blood cells and haecytogenesis of wild Euchiloglanis davidi. J. Fish. China 2009, 33, 581–589. [Google Scholar] [CrossRef]
- Ding, R.H. The Fishes of Sichuan, China; Sichuan Publishing House of Science and Technology: Chengdu, China, 1994. [Google Scholar]
- Geng, G.B.; Yan, S.G. Lists of wild animals under special state protection in China. Chin. J. Wildlife 2021, 42, 605–640. [Google Scholar]
- Chu, X.L. Taxonomic revision of the genera Pareuchiloglanis and Euchiloglanis. Zool Res. 1981, 2, 25–31. [Google Scholar]
- Zhou, W.; Li, X.; Thomson, A.W. Two new species of the Glyptosternine catfish genus Euchiloglanis (Teleostei: Sisoridae) from southwest China with redescriptions of E. davidi and E. kishinouyei. Zootaxa 2011, 2871, 1–18. [Google Scholar] [CrossRef]
- Guo, X.G.; Zhang, Y.G.; He, S.P. Morphological variations and species validity of genus Euchiloglanis (Siluriformes: Sisoridae) in China. Acta Hydrobiol. Sin. 2004, 28, 260–268. [Google Scholar]
- Zou, Y.C.; Hu, H.J.; Zhang, P.Y.; Wen, Z.Y.; Wei, Q.W. The complete mitochondrial genome of Euchiloglanis davidi and its phylogenetic implications. Mitochondrial DNA Part B 2019, 4, 1249–1250. [Google Scholar] [CrossRef]
- Luo, Q.S.; Zhong, M.C. Morphological observations on the skull of Euchiloglanis davidi (Sisoridae). J. Southwest China Norm. Univ. 1990, 15, 233–238. [Google Scholar]
- Pan, Y.Y.; Feng, J.; Du, W.P.; Jiang, B.G.; Luo, B.; Pang, W. The study about rate of flesh in body and nutritional composition of muscle in Euchiloglanic spp. Acta Hydrobiol. Sin. 2009, 33, 980–985. [Google Scholar] [CrossRef]
- Li, H.J.; Xie, C.X. Age and growth of the Tibetan catfish Glyptosternum maculatum in the Brahmaputra River, China. Zool. Stud. 2008, 47, 555–563. [Google Scholar]
- Chen, Z.; Lu, H.; Liu, W.; Liu, K.; Chen, X. Beak microstructure estimates of the age, growth, and population structure of purpleback flying squid (Sthenoteuthis oualaniensis) in the Xisha Islands Waters of the South China Sea. Fishes 2022, 7, 187. [Google Scholar] [CrossRef]
- Bostancı, D.; Yedier, S.; Kontaş, S.; Kurucu, G.; Polat, N. Length-weight, length-length relationships and condition factors of some fish species in Yalıköy Stream (Ordu-Turkey). Yunus Res. Bull. 2017, 17, 375–383. [Google Scholar]
- Yedier, S. Estimation of some population parameters of Squalius cephalus (Linnaeus 1758) in Tabakane Stream (Ordu-Turkey). Sak. Univ. J. Sci. 2022, 26, 14–23. [Google Scholar] [CrossRef]
- Wang, Y.F.; Lei, X.H.; Wen, X.; Fang, G.H.; Tan, Q.F.; Tian, Y.; Wang, C.; Wang, H. Effects of damming and climatic change on the eco-hydrological system: A case study in the Yalong River, southwest China. Ecol. Indic. 2019, 105, 663–674. [Google Scholar] [CrossRef]
- Jin, G.; Wang, Y.S.; Wu, W.B.; Guo, T.; Xu, J.L. Distribution features of landslides in the Yalong River Basin, Southwest China. Environ. Earth Sci. 2021, 80, 285. [Google Scholar] [CrossRef]
- Wang, Y.R.; Li, J.; Li, K.F.; Rui, J.L. Hydraulic parameters of habitat requirement for fish in water reduced river reach due to diversion of hydropower station. J. Hydraul. Eng. 2007, 38, 107–111. [Google Scholar] [CrossRef]
- Ding, C.Z. Reproductive biology of Glyptosternon maculatum in Yarlung Tsangpo River in Tibet, China. Acta Hydrobiol. Sin. 2010, 34, 762–768. [Google Scholar] [CrossRef]
- Esmaeili, H.R.; Sayyadzadeh, G.; Chermahini, M.A. Sexual dimorphism in two catfish species, Mystus pelusius (Solander, 1794) and Glyptothorax silviae Coad, 1981 (Teleostei: Siluriformes). Turk. J. Zool. 2017, 41, 144–149. [Google Scholar] [CrossRef]
- Ding, C.Z.; Chen, Y.F.; He, D.K. Assessing the accuracy of using whole and sectioned vertebrae to determine the age of an endemic sisorid catfish, Glyptosternon maculatum, in Tibet, China. Ichthyol. Res. 2011, 58, 72–76. [Google Scholar] [CrossRef]
- Ma, B.S.; Xie, C.X.; Huo, B.; Yang, X.F.; Li, P. Age validation, and comparison of otolith, vertebra and opercular bone for estimating age of Schizothorax o’connori in the Yarlung Tsangpo River, Tibet. Environ. Biol. Fishes 2011, 90, 159–169. [Google Scholar] [CrossRef]
- Paul, L.J.; Horn, P.L. Age and growth of sea perch (Helicolenus percoides) from two adjacent areas off the east coast of South Island, New Zealand. Fish. Res. 2009, 95, 169–180. [Google Scholar] [CrossRef]
- Pauly, D. Fish Population Dynamics in Tropical Waters: A Manual for Use with Programmable Calculators; ICLARM: Manila, Philippines, 1984; Volume 8, p. 325. [Google Scholar]
- Von Bertalanffy, L. A quantitative theory of organic growth (inquiries on growth laws. II). Hum. Biol. 1938, 10, 181–213. [Google Scholar]
- Munro, J.L.; Pauly, D. A simple method for comparing the growth of fishes and invertebrates. Fishbyte 1983, 1, 5–6. [Google Scholar]
- Casselman, J.M. Determination of age and growth. Biol. Fish Growth 1987, 209–242. [Google Scholar]
- Campana, S.E. Accuracy, precision and quality control in age determination, including a review of the use and abuse of age validation methods. J. Fish Biol. 2001, 59, 197–242. [Google Scholar] [CrossRef]
- Khan, M.A.; Khan, S. Age and growth assessment of the near-threatened fish Rita rita (Siluriformes: Bagridae) in the Ganges basin. Rev. Biol. Trop. 2023, 71. [Google Scholar] [CrossRef]
- Shen, Y.J.; Pu, D.Y.; Gao, M.; Wang, H.L.; Wang, Z.J. Studies on the relationship between the age and growth of Glyptothorax fukiensis in the Jialing River. J. Southwest Agric. Univ. 2005, 27, 106–110. [Google Scholar]
- Andrian Feitoza, L.; Kiyoshi Okada, E.; Maria Ambrósio, A. Age and growth of Pterodoras granulosus (Valenciennes, 1833),(Siluriformes, Doradidae) in Itaipu reservoir, state of Paraná, Brazil. Acta Sci. Biol. Sci. 2004, 26, 47–53. [Google Scholar]
- Balboni, L.; Vargas, F.; Colautti, D. Age and growth of Pseudoplatystoma corruscans (Siluriformes: Pimelodidae) at the confluence of the Paraná and Paraguay rivers. Neotrop. Ichthyol. 2021, 19. [Google Scholar] [CrossRef]
- Wang, Y.M.; Cao, M.; Xie, B.W. Age and growth of Euchiloglanis kishinouyei in the Dadu River Basin. Chin. J. Zool. 2016, 51, 228–240. [Google Scholar]
- Huang, J.; Huang, Z.H.; Wei, Q.W. Age and growth of Creteuchiloglianis macropterus. Acta Hydrobiol. Sin. 2018, 42, 10. [Google Scholar]
- Jin, H.Y.; Li, L.; Jin, X.; Wang, N.M.; Ma, Q.Z.; Yin, J.S.; Ma, B. Age and growth of Pseudecheneis sulcatus in lower reach of Yarlung Zangbo River of Medog county reach. Freshw. Fish 2020, 50, 10–17. [Google Scholar]
- Froese, R. Cube law, condition factor and weight–length relationships: History, meta-analysis and recommendations. J. Appl. Ichthyol. 2006, 22, 241–253. [Google Scholar] [CrossRef]
- Chen, F.; Ma, B.S.; Xie, C.X.; Zhang, H.J. Age and growth characteristics of Pareuchiloglanis kamengensis in Tibet. Freshw. Fish 2018, 48, 45–51. [Google Scholar]
- Zhang, C.; Zhu, T.B.; Zhou, C.W.; Pan, Y.Z.; Liu, F.; Mou, Z.B. Length–weight relationships of three small fish species caught in the Yarlung Zangbo River basin, China. J. Appl. Ichthyol. 2018, 34, 1009–1010. [Google Scholar] [CrossRef]
- Branstetter, S. Age and growth estimates for blacktip, Carcharhinus limbatus, and spinner, C. brevipinna, sharks from the northwestern Gulf of Mexico. Copeia 1987, 1987, 964–974. [Google Scholar] [CrossRef]
- Gallucci, V.F.; Quinn, T.J. Reparameterizing, fitting, and testing a simple growth model. T. Aa. Fish. Soc. 1979, 108, 14–25. [Google Scholar] [CrossRef]
- Copp, G.H.; Fox, M.G.; Przybylski, M.; Godinho, F.N.; Gispert, A. Life-time growth patterns of pumpkinseed Lepomis gibbosus introduced to Europe, relative to North American populations. Folia Zool. 2004, 53, 237–254. [Google Scholar]
- Wootton, R.J. The effect of size of food ration on egg production in the female three-spined stickleback, Gasterosteus aculeatus L. J. Fish Biol. 1973, 5, 89–96. [Google Scholar] [CrossRef]
- Healey, M.C. The dynamics of exploited lake trout populations and implications for management. J. Wildlife Manag. 1978, 42, 307–328. [Google Scholar] [CrossRef]
- Suzuki, H.I.; Agostinho, A.A.; Winemiller, K.O. Relationship between oocyte morphology and reproductive strategy in loricariid catfishes of the Paraná River, Brazil. J. Fish Biol. 2000, 57, 791–807. [Google Scholar] [CrossRef]
- Shi, J.R.; Luo, D.M.; Liang, L.Y.; Wang, Y.M.; Peng, P.L.; Xie, B.W. Individual fecundity of Euchiloglanis kishinouyei in the Zumuzu River. Sichuan J. Zool. 2019, 38, 305–310. [Google Scholar] [CrossRef]
- Lin, P.C.; Hu, H.M.; Gong, Z.; Wang, J.; Gao, X. Reproductive Characteristics of Pseudecheneis sulcatus (Siluriforms: Sisoridae) in the Lower Yarlung Zangbo River, Tibet. Fishes 2023, 8, 106. [Google Scholar] [CrossRef]
- Jin, H.Y.; Li, L.; Jin, X.; Wang, N.M.; Ma, Q.Z.; Yin, J.S.; Ma, B. Fecundity of Pseudecheneis sulcatus in downstream of Yarlung Zangbo River in Xizang. Fish Sci. 2020, 39, 744–751. [Google Scholar] [CrossRef]
- Chu, Q.Z.; Chen, G.; Zhang, J.D.; Pan, C.H.; Zhou, H. Feeding habits and productivity of Glyptothorax fukiensis in Waters of Western Guangdong. J. Guangdong Ocean Univ. 2009, 29, 10–13. [Google Scholar] [CrossRef]
- Al-Husaini, M.; Al-Baz, A.; Dashti, T.; Rajab, S.; Husain, H. Age, growth, and reproductive parameters of four species of sea catfish (Siluriformes: Ariidae) from Kuwaiti waters. Reg. Stud. Mar. Sci. 2021, 46, 101885. [Google Scholar] [CrossRef]
- Roff, D.A. The evolution of life histories: Theory and analysis. In Theory and Analysis; Chapman and Hall: London, UK, 1992; Volume 6, pp. 760–762. [Google Scholar]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rank | Readability Level | Annuli Characteristics |
|---|---|---|
| 1 | Excellent | The annulus is clear and obvious, which can accurately identify the age |
| 2 | Good | The annulus is basically clear, and can identify the age accurately |
| 3 | Average | Some annuli are not clear or cannot be accurately identified |
| 4 | Poor | Most annuli cannot be accurately identified and may count as possible |
| 5 | Unreadable | Annulus cannot be identified |
| Calcified Structure | Proportion of Readability Scores (%) | ||||
|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | |
| Vertebrae | 47 | 27.35 | 8.55 | 9.40 | 7.69 |
| Age | Number | Standard Length (mm) | Weight (g) | ||
|---|---|---|---|---|---|
| Mean ± S.D. | Range | Mean ± S.D. | Range | ||
| 1 | 31 | 76.6 ± 15.8 | 67–85 | 7.0 ± 7.2 | 3.42–11.16 |
| 2 | 41 | 80.0 ± 16.0 | 67–97 | 7.5 ± 7.2 | 4.30–11.68 |
| 3 | 18 | 101.7 ± 18.4 | 89–126 | 15.8 ± 8.1 | 8.57–36.02 |
| 4 | 3 | 126.3 ± 30.0 | 117–132 | 28.0 ± 15.2 | 17.20–38.36 |
| 5 | 12 | 145.3 ± 31.5 | 121–162 | 36.4 ± 14.7 | 28.09–44.40 |
| 6 | 16 | 149.9 ± 36.6 | 134–170 | 39.7 ± 17.6 | 28.98–53.54 |
| 7 | 13 | 160.9 ± 34.6 | 145–190 | 49.1 ± 17.0 | 36.07–75.06 |
| 8 | 5 | 160.2 ± 35.7 | 144–184 | 45.4 ± 17.5 | 36.60–59.05 |
| 9 | 1 | 159 | 159 | 46.86 | 46.86 |
| 10 | 1 | 179 | 179 | 71.38 | 71.38 |
| Total | 141 | 107.3 ± 35.6 | 67–190 | 20.8 ± 17.7 | 3.42–75.06 |
| Species | River Drainage | n | Length Range (mm) | Age Range | Growth Parameters | Reference | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| L∞ (mm) | k | t0 | b | Ø | ||||||
| Glyptosternum maculatum | Yarlung Zangbo River | 128 | 115–270 | 3–13 | 342.66 (♀) | 0.114 (♀) | 0.7488 (♀) | 3.1420 (♀) | 5.07 (♀) | [10] |
| 77 | 147–320 | 3–13 | 460.24 (♂) | 0.082 (♂) | 0.2312 (♂) | 3.1474 (♂) | 5.32 (♂) | |||
| Glyptothorax fukiensis | Jialing River | 238 | 42.9–102.9 | 1–5 | 140.20 | 0.254 | 0.0696 | 3.2130 | 3.70 | [28] |
| Euchiloglanis kishinouyei | Dadu River | 383 | 92–190 | 3–13 | 208.42 | 0.089 | 1.2000 | 2.9279 | 4.63 | [31] |
| Pseudecheneis sulcatus | Yarlung Zangbo River | 203 | 50–144 | 1–9 | 189.56 | 0.118 | 1.9810 | 2.9292 | 4.55 | [33] |
| Euchiloglanis davidi | Yalong River | 150 | 67–190 | 1–10 | 260.80 | 0.104 | 1.9420 | 2.6530 | 3.85 | This study |
| Species | River Drainage | n | Standard Length (mm) | Absolute Fecundity (Eggs) | Relative Fecundity (Eggs/g) | Reference |
|---|---|---|---|---|---|---|
| Glyptosternum maculatum | Yarlung Zangbo River | 128 | 133–243 | 525–2058 | 3.24–27.01 | [17] |
| Euchiloglanis kishinouyei | Zumuzu River | 69 | 120–165 | 123–489 | 2.80–13.95 | [43] |
| Pseudecheneis sulcatus | Yarlung Zangbo River | 52 | 114–200 | 247–2886 | 16.80–77.10 | [44] |
| Pseudecheneis sulcatus | Yarlung Zangbo River | 78 | 92.99–144.49 | 346–799 | 20.69–92.80 | [45] |
| Glyptothorax fukiensis | Western Guangdong waters | 49 | 31.9–63.7 | 45–420 | 31.70–130.40 | [46] |
| Euchiloglanis davidi | Yalong River | 35 | 135–184 | 105–352 | 2.90–8.98 | This study |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, J.; Zhu, X.; Yang, X.; Hu, X.; Lin, P.; Xu, B.; Wei, K.; Ma, B. Age, Growth, and Reproductive Biology of Euchiloglanis davidi in the Middle and Lower Yalong River, China. Fishes 2023, 8, 435. https://doi.org/10.3390/fishes8090435
Chen J, Zhu X, Yang X, Hu X, Lin P, Xu B, Wei K, Ma B. Age, Growth, and Reproductive Biology of Euchiloglanis davidi in the Middle and Lower Yalong River, China. Fishes. 2023; 8(9):435. https://doi.org/10.3390/fishes8090435
Chicago/Turabian StyleChen, Jie, Xiangyun Zhu, Xin Yang, Xianqin Hu, Pengcheng Lin, Bin Xu, Kaijin Wei, and Baoshan Ma. 2023. "Age, Growth, and Reproductive Biology of Euchiloglanis davidi in the Middle and Lower Yalong River, China" Fishes 8, no. 9: 435. https://doi.org/10.3390/fishes8090435