
Different Diets Based on Yellow Mealworm (Tenebrio molitor)—Part B: Modification of the Intestinal Inflammatory Response and the Microbiota Composition of Rainbow Trout (Oncorhynchus mykiss)
 , , , , , , , and
, , , , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Diets, Animals and Rearing Conditions
2.2. Sample Collection
2.3. Chemical Analyses
2.4. Non-Specific Immune Status
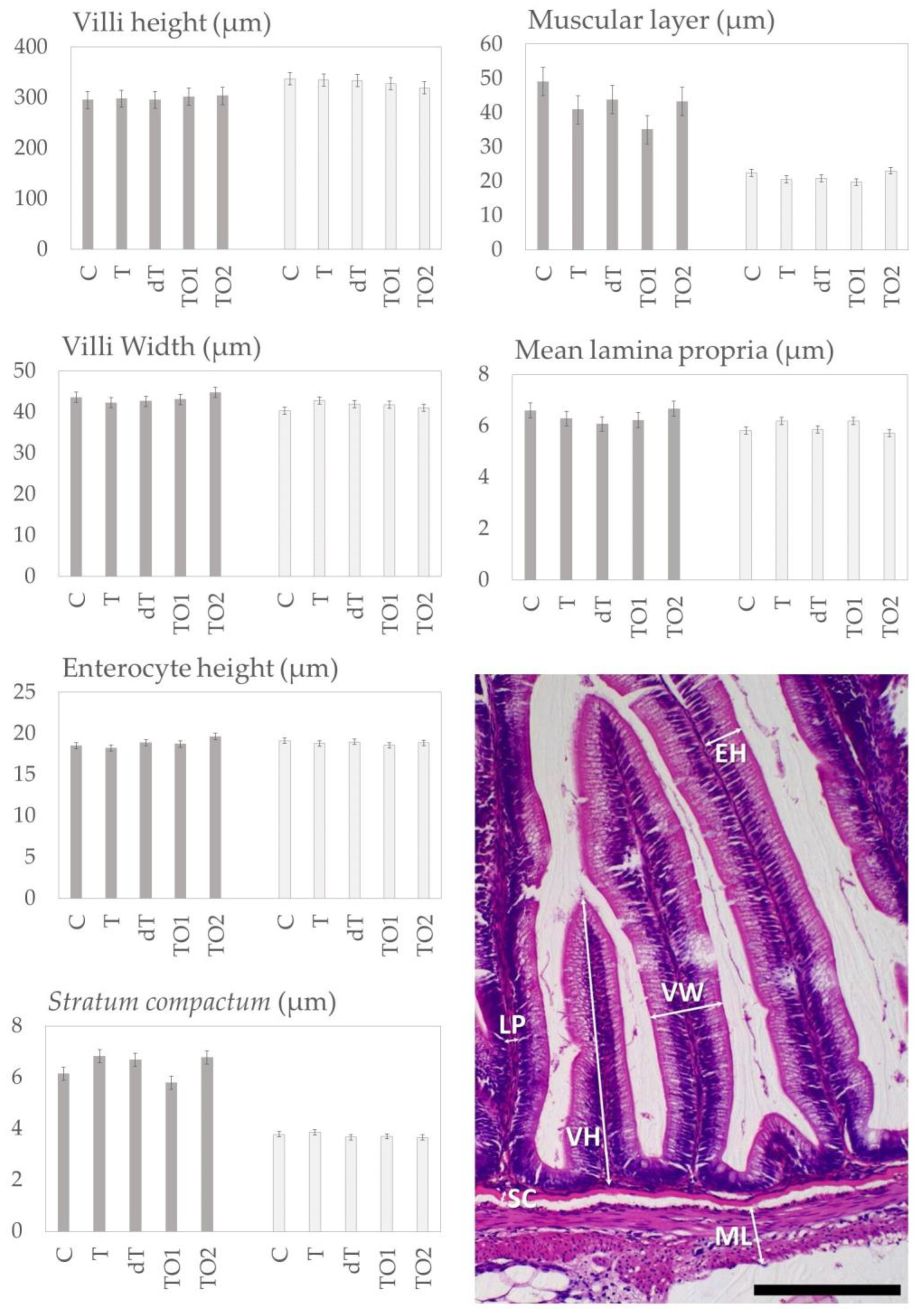
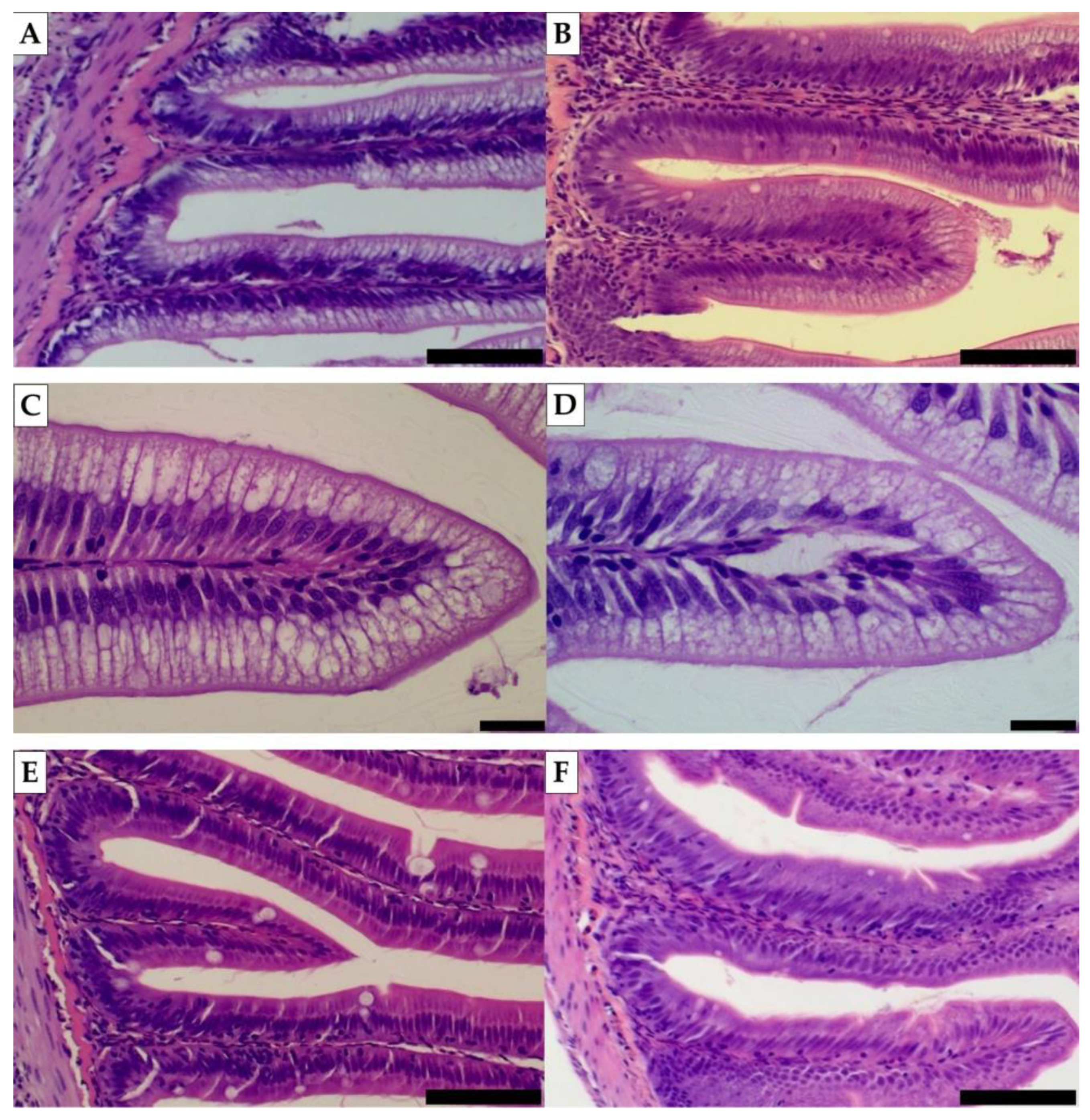
2.5. Histomorphology
2.5.1. Samples Processing
2.5.2. Distal Intestine and Pyloric Caeca Histomorphology Analyses
2.6. Distal Intestine Gut Content Microbiota Analysis
2.7. Statistical Analyses
3. Results and Discussion
3.1. Protein Use, Biometric Indices and Butchering Yield
3.2. Immunological System
3.3. Gut Health
3.3.1. Intestinal Histomorphology
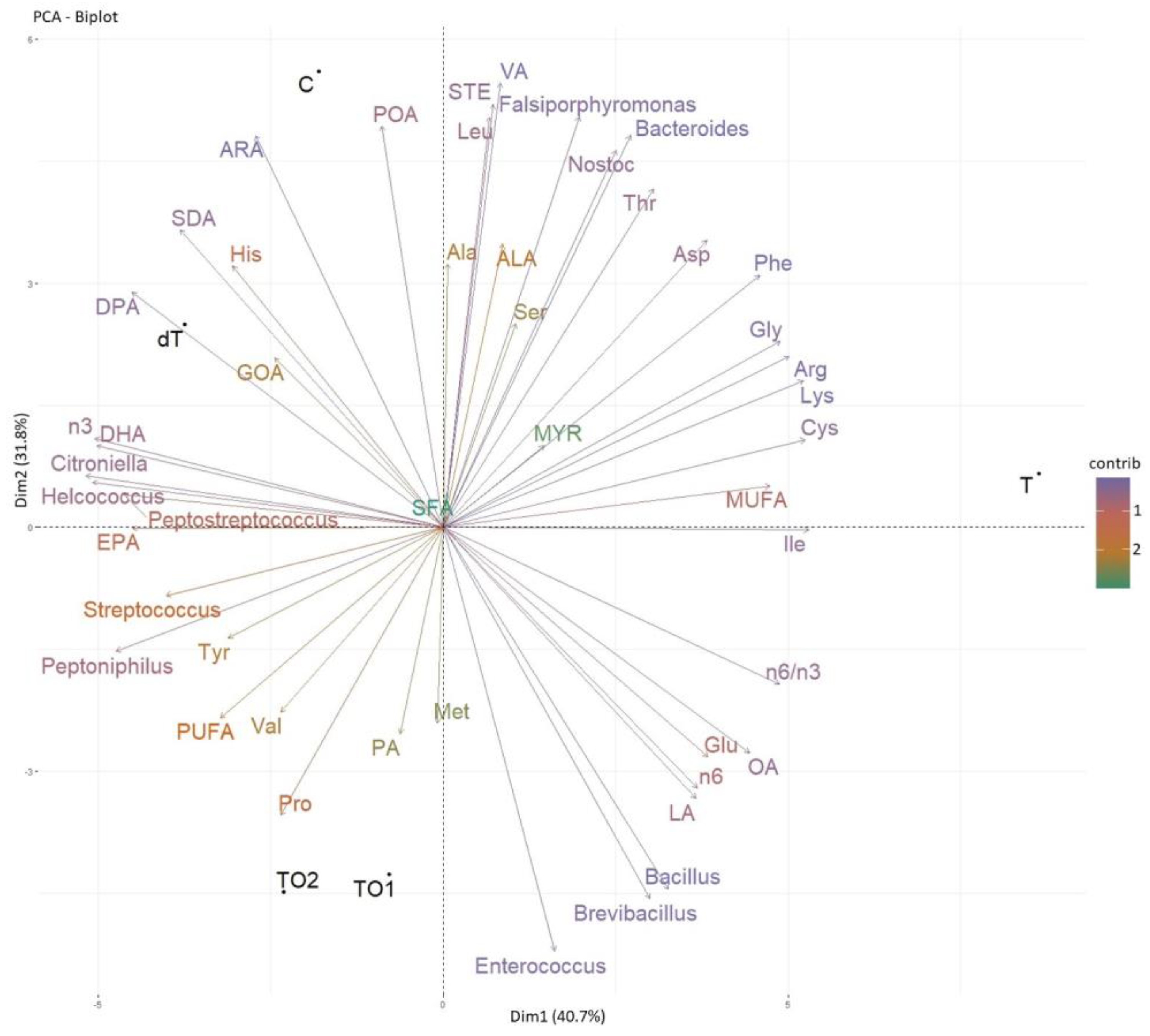
3.3.2. Gut Content Microbiota Analyses
- Alpha diversity
- Bacterial composition

{kind=link}
{kind=link}
{kind=link}
| Alpha Diversity Index | C | T | dT | TO1 | TO2 | SEM | p Value | F Value | DF |
| Chao1 | 309.75 a | 278.26 ab | 240.81 bc | 230.33 c | 224 c | 9.68 | <0.0001 | 14.12 | 4 |
| Simpson | 0.93 | 0.95 | 0.91 | 0.93 | 0.93 | 0.01 | 0.1253 | 2.15 | 4 |
| Shannon | 3.85 ab | 4.03 a | 3.67 b | 3.78 ab | 3.79 ab | 0.07 | 0.034 | 3.47 | 4 |
| Relative OTU Composition at Phylum Level | C | T | dT | TO1 | TO2 | SEM | p Value | F Value | DF |
| Actinomycetota 1 | 2.76 a | 1.83 b | 2.05 b | 2.21 ab | 1.78 b | 0.15 | 0.0018 | 7.28 | 4 |
| Bacteroidota 2 | 12.44 a | 11.21 ab | 8.52 ab | 5.84 b | 5.50 b | 1.41 | 0.0103 | 4.86 | 4 |
| Cyanobacteria | 13.86 a | 12.51 a | 7.99 b | 4.75 c | 6.11 bc | 0.58 | <0.0001 | 47.33 | 4 |
| Bacillota 3 | 65.67 c | 69.17 bc | 78.78 ab | 79.04 ab | 83.59 a | 2.97 | 0.0034 | 6.36 | 4 |
| Pseudomonadota 4 | 4.14 | 4.08 | 1.92 | 7.61 | 2.01 | 2.79 | 0.6128 | 0.69 | 4 |
| Other | 1.13 | 1.20 | 0.74 | 0.54 | 1.00 | - | - | - | - |
| Relative OTU Composition at Genus Level | C | T | dT | TO1 | TO2 | SEM | p Value | F Value | DF |
| Bacteroides | 3.7 a | 3.61 ab | 2.59 abc | 1.69 bc | 1.42 c | 0.44 | 0.0054 | 5.69 | 4 |
| Falsiporphyromonas | 7.47 a | 6.46 ab | 5.11 ab | 3.62 b | 3.55 b | 0.83 | 0.0155 | 4.35 | 4 |
| Nostoc | 13.18 a | 11.91 a | 7.56 b | 4.49 c | 5.77 bc | 0.56 | <0.0001 | 46.84 | 4 |
| Bacillus | 0.21 b | 2.94 a | 0.25 b | 2.79 a | 2.5 a | 0.17 | <0.0001 | 69.58 | 4 |
| Brevibacillus | 0.02 b | 8.06 a | 0.004 b | 7.12 a | 8.05 a | 0.5 | <0.0001 | 71.58 | 4 |
| Enterococcus | 0.23 b | 1.78 a | 0.27 b | 2.51 a | 2.41 a | 0.19 | <0.0001 | 33.63 | 4 |
| Streptococcus | 7.45 | 7.16 | 8.75 | 8.12 | 7.85 | 0.47 | 0.1948 | 1.73 | 4 |
| Helcococcus | 1.18 ab | 0.86 b | 1.39 a | 1.22 a | 1.13 ab | 0.08 | 0.0055 | 5.67 | 4 |
| Peptoniphilus | 14.86 ab | 13.06 b | 18.47 a | 16.2 ab | 17.64 a | 0.9 | 0.0048 | 5.86 | 4 |
| Citroniella | 2.01 a | 1.55 b | 2.27 a | 2.04 a | 1.95 ab | 0.1 | 0.0023 | 6.89 | 4 |
| Peptostreptococcus | 20.23 bc | 15.84 c | 27.04 a | 19.33 bc | 23.05 ab | 1.55 | 0.0018 | 7.32 | 4 |
| Other | 29.46 | 26.77 | 26.3 | 30.86 | 24.67 | - | p-value | F-value | DF |
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. The State of World Fisheries and Aquaculture 2022: Towards Blue Transformation; FAO: Rome, Italy, 2022. [Google Scholar] [CrossRef]
- Kok, B.; Malcorps, W.; Tlusty, M.F.; Eltholth, M.M.; Auchterlonie, N.A.; Little, D.C.; Harmsen, R.; Newton, R.W.; Davies, S.J. Fish as feed: Using economic allocation to quantify the Fish In: Fish Out ratio of major fed aquaculture species. Aquaculture 2020, 528, 735474. [Google Scholar] [CrossRef]
- Aas, T.S.; ÅsgÅrd, T.; Ytrestøyl, T. Utilization of feed resources in the production of rainbow trout (Oncorhynchus mykiss) in Norway in 2020. Aquac. Rep. 2022, 26, 101317. [Google Scholar] [CrossRef]
- Barroso, F.G.; de Haro, C.; Sánchez-Muros, M.-J.; Venegas, E.; Martínez-Sánchez, A.; Pérez-Bañón, C. The potential of various insect species for use as food for fish. Aquaculture 2014, 422–423, 193–201. [Google Scholar] [CrossRef]
- Makkar, H.P.; Tran, G.; Heuzé, V.; Ankers, P. State-of-the-art on use of insects as animal feed. Anim. Feed Sci. Technol. 2014, 197, 1–33. [Google Scholar] [CrossRef]
- Alfiko, Y.; Xie, D.; Astuti, R.T.; Wong, J.; Wang, L. Insects as a feed ingredient for fish culture: Status and trends. Aquac. Fish. 2022, 7, 166–178. [Google Scholar] [CrossRef]
- Nasdaq. Fishmeal Value. Available online: https://data.nasdaq.com/data/ODA/PFSHMEAL_USD (accessed on 20 January 2023).
- O’Keefe, T. Plant Protein Ingredients for Aquaculture Feeds: Use Considerations & Quality Standards; U.S. Soybean Export Council: Singapore, 2002. [Google Scholar]
- Krogdahl, Å.; Penn, M.; Thorsen, J.; Refstie, S.; Bakke, A.M. Important antinutrients in plant feedstuffs for aquaculture: An update on recent findings regarding responses in salmonids. Aquac. Res. 2010, 41, 333–344. [Google Scholar] [CrossRef]
- Hajra, A.; Mazumder, A.; Verma, A.; Ganguly, D.P.; Mohanty, B.P.; Sharma, A.P. Antinutritional factors in plant origin fish feed ingredients: The problems and probable remedies. In Advances in Fish Research; Goswami, U.C., Ed.; Narendra Publishing House: Delhi, India, 2013; Chapter 11; Volume V, ISBN 9789380428581. [Google Scholar]
- Baeverfjord, G.; Krogdahl, Å. Development and regression of soybean meal induced enteritis in Atlantic salmon, Salmo salar L., distal intestine: A comparison with the intestines of fasted fish. J. Fish. Dis. 1996, 19, 375–387. [Google Scholar] [CrossRef]
- Djordjevic, B.; Morales-Lange, B.; Øverland, M.; Mercado, L.; Lagos, L. Immune and proteomic responses to the soybean meal diet in skin and intestine mucus of Atlantic salmon (Salmo salar L.). Aquac. Nutr. 2021, 27, 929–940. [Google Scholar] [CrossRef]
- Todd Lorenz, R.; Cysewski, G.R. Commercial potential for Haematococcus microalgae as a natural source of astaxanthin. Trends Biotechnol. 2000, 18, 160–167. [Google Scholar] [CrossRef]
- Roy, S.S.; Pal, R. Microalgae in aquaculture: A review with special references to nutritional value and fish dietetics. Proc. Zool. Soc. 2015, 68, 1–8. [Google Scholar] [CrossRef]
- Hansen, J.Ø.; Hofossæter, M.; Sahlmann, C.; Ånestad, R.; Reveco-Urzua, F.E.; Press, C.M.; Mydland, L.T.; Øverland, M. Effect of Candida utilis on growth and intestinal health of Atlantic salmon (Salmo salar) parr. Aquaculture 2019, 511, 734239. [Google Scholar] [CrossRef]
- Agboola, J.O.; Øverland, M.; Skrede, A.; Hansen, J.Ø. Yeast as major protein-rich ingredient in aquafeeds: A review of the implications for aquaculture production. Rev. Aquac. 2021, 13, 949–970. [Google Scholar] [CrossRef]
- Van Huis, A. Potential of insects as food and feed in assuring food security. Annu. Rev. Entomol. 2013, 58, 563–583. [Google Scholar] [CrossRef] [PubMed]
- Van Huis, A.; Oonincx, D.G.A.B. The environmental sustainability of insects as food and feed. A review. Agron. Sustain. Dev. 2017, 37, 43. [Google Scholar] [CrossRef]
- Henry, M.A.; Gai, F.; Enes, P.; Pérez-Jiménez, A.; Gasco, L. Effect of partial dietary replacement of fishmeal by yellow mealworm (Tenebrio molitor) larvae meal on the innate immune response and intestinal antioxidant enzymes of rainbow trout (Oncorhynchus mykiss). Fish Shellfish Immunol. 2018, 83, 308–313. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Ji, H.; Yu, H.; Zhou, J. Influence of dietary black soldier fly (Hermetia illucens Linnaeus) pulp on growth performance, antioxidant capacity and intestinal health of juvenile mirror carp (Cyprinus carpio var. specularis). Aquac. Nutr. 2020, 26, 432–443. [Google Scholar] [CrossRef]
- Melenchón, F.; Larrán, A.M.; de Mercado, E.; Hidalgo, M.C.; Cardenete, G.; Barroso, F.G.; Fabrikov, D.; Lourenço, H.M.; Pessoa, M.F.; Tomás-Almenar, C. Potential use of black soldier fly (Hermetia illucens) and mealworm (Tenebrio molitor) insectmeals in diets for rainbow trout (Oncorhynchus mykiss). Aquac. Nutr. 2021, 27, 491–505. [Google Scholar] [CrossRef]
- Hidalgo, M.C.; Morales, A.E.; Pula, H.J.; Tomás-Almenar, C.; Sánchez-Muros, M.J.; Melenchón, F.; Fabrikov, D.; Cardenete, G. Oxidative metabolism of gut and innate immune status in skin and blood of tench (Tinca tinca) fed with different insect meals (Hermetia illucens and Tenebrio molitor). Aquaculture 2022, 529, 735731. [Google Scholar] [CrossRef]
- Ngo, D.-N.; Kim, M.-M.; Kim, S.-K. Chitin oligosaccharides inhibit oxidative stress in live cells. Carbohydr. Polym. 2008, 74, 228–234. [Google Scholar] [CrossRef]
- Ngo, D.-H.; Kim, S.-K. Chapter two—Antioxidant effects of chitin, chitosan, and their derivatives. Adv. Food Nutr. Res. 2014, 73, 15–31. [Google Scholar] [CrossRef]
- Lee, C.G.; da Silva, C.A.; Lee, J.Y.; Hartl, D.; Elias, J.A. Chitin regulation of immune responses: An old molecule with new roles. Curr. Opin. Immunol. 2008, 20, 684–689. [Google Scholar] [CrossRef] [PubMed]
- Lindsay, G.J.H.; Walton, M.J.; Adron, J.W.; Fletcher, T.C.; Cho, C.Y.; Cowey, C.B. The growth of rainbow trout (Salmo gairdneri) given diets containing chitin and its relationship to chitinolytic enzymes and chitin digestibility. Aquaculture 1984, 37, 315–334. [Google Scholar] [CrossRef]
- Köprücü, K.; Özdemir, Y. Apparent digestibility of selected feed ingredients for Nile tilapia (Oreochromis niloticus). Aquaculture 2005, 250, 308–316. [Google Scholar] [CrossRef]
- Alegbeleye, W.O.; Obasa, S.O.; Olude, O.O.; Otubu, K.; Jimoh, W. Preliminary evaluation of the nutritive value of the variegated grasshopper (Zonocerus variegatus L.) for African catfish Clarias gariepinus (Burchell. 1822) fingerlings. Aquac. Res. 2012, 43, 412–420. [Google Scholar] [CrossRef]
- Lieberman, S.; Enig, M.G.; Preuss, H.G. A review of monolaurin and lauric acid: Natural virucidal and bactericidal agents. J. Altern. Complement. Med. 2006, 12, 310–314. [Google Scholar] [CrossRef]
- Gasco, L.; Finke, M.; van Huis, A. Can diets containing insects promote animal health? J. Insects Food Feed 2018, 4, 1–4. [Google Scholar] [CrossRef]
- Finke, M.D. Complete nutrient composition of commercially raised invertebrates used as food for insectivores. Zoo Biol. 2002, 21, 269–285. [Google Scholar] [CrossRef]
- Otvos, L. Antibacterial peptides isolated from insects. J. Peptide Sci. 2000, 6, 497–511. [Google Scholar] [CrossRef]
- Huyben, D.; Vidakovic, A.; Hallgren, S.W.; Langeland, M. High-throughput sequencing of gut microbiota in rainbow trout (Oncorhynchus mykiss) fed larval and pre-pupae stages of black soldier fly (Hermetia illucens). Aquaculture 2019, 500, 485–491. [Google Scholar] [CrossRef]
- Terova, G.; Rimoldi, S.; Ascione, C.; Gini, E.; Ceccotti, C.; Gasco, L. Rainbow trout (Oncorhynchus mykiss) gut microbiota is modulated by insect meal from Hermetia illucens prepupae in the diet. Rev. Fish Biol. Fish. 2019, 29, 465–486. [Google Scholar] [CrossRef]
- Weththasinghe, P.; Rocha, S.D.C.; Øyås, O.; Lagos, L.; Hansen, J.Ø.; Mydland, L.T.; Øverland, M. Modulation of Atlantic salmon (Salmo salar) gut microbiota composition and predicted metabolic capacity by feeding diets with processed black soldier fly (Hermetia illucens) larvae meals and fractions. Anim. Microbiome 2022, 4, 9. [Google Scholar] [CrossRef] [PubMed]
- Melenchón, F.; Larrán, A.M.; Sanz, M.-Á.; Rico, D.; Fabrikov, D.; Barroso, F.G.; Galafat, A.; Alarcón, F.J.; Morales, A.E.; Hidalgo, M.C.; et al. Different diets based on yellow mealworm (Tenebrio molitor)—Part A: Facing the decrease of omega-3 fatty acids in fillets of rainbow trout (Oncorhynchus mykiss). Fishes 2023, accepted. [Google Scholar]
- National Research Council. Nutrient Requirements of Fish; National Academies Press: Washington, DC, USA, 1993. [Google Scholar]
- Blanco Cachafeiro, M.C. La Trucha: Cría Industrial, 2nd ed.; Mundi-Prensa: Madrid, Spain, 2005. [Google Scholar]
- Cho, C.; Slinger, S.; Bayley, H. Bioenergetics of salmonid fishes: Energy intake, expenditure and productivity. Comp. Biochem. Physiol. Part B Comp. Biochem. 1982, 73, 25–41. [Google Scholar] [CrossRef]
- European Parliament. Book Directive 2010/63/EU of the European Parliament and of the Council of 22 September 2010 on the Protection of Animals Used for Scientific Purposes; European Parliament: Strasbourg, France, 2010; pp. 33–78. [Google Scholar]
- Real Decreto 53/2013, de 1 de Febrero, por El Que Se Establecen las Normas Básicas Aplicables para la Protección de los Animales Utilizados en Experimentación y Otros Fines Científicos, Incluyendo la Docencia. 2013. Available online: https://www.boe.es/eli/es/rd/2013/02/01/53 (accessed on 27 January 2023).
- Atkinson, J.L.; Hilton, J.W.; Slinger, S.J. Evaluation of acid-insoluble ash as an indicator of feed digestibility in rainbow trout (Salmo gairdneri). Can. J. Fish. Aquat. Sci. 1984, 41, 1384–1386. [Google Scholar] [CrossRef]
- Janssen, R.H.; Vincken, J.-P.; van den Broek, L.A.M.; Fogliano, V.; Lakemond, C.M.M. Nitrogen-to-protein conversion factors for three edible insects: Tenebrio molitor, Alphitobius diaperinus, and Hermetia illucens. J. Agric. Food Chem. 2017, 65, 2275–2278. [Google Scholar] [CrossRef]
- Swain, P.; Dash, S.; Sahoo, P.K.; Routray, P.; Sahoo, S.K.; Gupta, S.D.; Meher, P.K.; Sarangi, N. Non-specific immune parameters of brood Indian major carp Labeo rohita and their seasonal variations. Fish Shellfish Immunol. 2007, 22, 38–43. [Google Scholar] [CrossRef] [PubMed]
- Mashiter, K.E.; Morgan, M.R. Carbonic anhydrase levels in the tissues of flounders adapted to sea water and fresh water. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 1975, 52, 713–717. [Google Scholar] [CrossRef] [PubMed]
- Thompson, I.; Choubert, G.; Houlihan, D.F.; Secombes, C.J. The effect of dietary vitamin A and astaxanthin on the immunocompetence of rainbow trout. Aquaculture 1995, 133, 91–102. [Google Scholar] [CrossRef]
- Huang, Z.-H.; Ma, A.-J.; Wang, X. The immune response of turbot, Scophthalmus maximus (L.), skin to high water temperature. J. Fish Dis. 2011, 34, 619–627. [Google Scholar] [CrossRef]
- Mohanty, B.R.; Sahoo, P.K. Immune responses and expression profiles of some immune-related genes in Indian major carp, Labeo rohita to Edwardsiella tarda infection. Fish Shellfish Immunol. 2010, 28, 613–621. [Google Scholar] [CrossRef]
- Panigrahi, A.; Kiron, V.; Puangkaew, J.; Kobayashi, T.; Satoh, S.; Sugita, H. The viability of probiotic bacteria as a factor influencing the immune response in rainbow trout Oncorhynchus mykiss. Aquaculture 2005, 243, 241–254. [Google Scholar] [CrossRef]
- Bradford, M. A rapid and sensitive method for the quantitation of microgram quantities of protein using the principle of protein dye-binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Melenchón, F.; de Mercado, E.; Pula, H.J.; Cardenete, G.; Barroso, F.G.; Fabrikov, D.; Lourenço, H.M.; Pessoa, M.-F.; Lagos, L.; Weththasinghe, P.; et al. Fishmeal dietary replacement up to 50%: A comparative study of two insect meals for rainbow trout (Oncorhynchus mykiss). Animals 2022, 12, 179. [Google Scholar] [CrossRef]
- Hernández, M.; Quijada, N.M.; Rodríguez-Lázaro, D.; Eiros, J.M. Bioinformatics of next generation sequencing in clinical microbiology diagnosis. Rev. Argent. Microbiol. 2020, 52, 150–161. [Google Scholar] [CrossRef]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef]
- Joshi, N.A.; Fass, J.N. Sickle: A Sliding-Windows, Adaptive, Quality-Based Trimming Tool for FastQ Files; Version 1.33; 2011; Available online: https://github.com/najoshi/sickle (accessed on 27 April 2023).
- Abad, D.; Quijada, N.M.; Hernández, M. QiimeReporter. 2019. Available online: https://github.com/dabadgarcia/qiimereporter (accessed on 27 April 2023).
- Callahan, B.; McMurdie, P.; Rosen, M.; Han, A.W.; Johnson, A.J.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naïve bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Chao, A. Nonparametric estimation of the number of classes in a population. Scand. J. Stat. 1984, 11, 265–270. Available online: https://www.jstor.org/stable/4615964 (accessed on 27 April 2023).
- Simpson, E.H. Measurement of diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Shannon, C.E. A mathematical theory of communication. BSTJ 1948, 27, 379–423. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 27 April 2023).
- RStudio Team. RStudio: Integrated Development Environment for R; RStudio, PBC: Boston, MA, USA, 2022; Available online: http://www.rstudio.com/ (accessed on 27 April 2023).
- Kroeckel, S.; Harjes, A.-G.E.; Roth, I.; Katz, H.; Wuertz, S.; Susenbeth, A.; Schulz, C. When a turbot catches a fly: Evaluation of a pre-pupae meal of the Black Soldier Fly (Hermetia illucens) as fish meal substitute—Growth performance and chitin degradation in juvenile turbot (Psetta maxima). Aquaculture 2012, 364–365, 345–352. [Google Scholar] [CrossRef]
- Marono, S.; Piccolo, G.; Loponte, R.; di Meo, C.; Attia, Y.A.; Nizza, A.; Bovera, F. In vitro crude protein digestibility of Tenebrio molitor and Hermetia illucens insect meals and its correlation with chemical composition traits. J. Anim. Sci. 2015, 14, 3889. [Google Scholar] [CrossRef]
- German, D.P.; Horn, M.H. Gut length and mass in herbivorous and carnivorous prickleback fishes (Teleostei: Stichaeidae): Ontogenetic, dietary, and phylogenetic effects. Mar. Biol. 2006, 148, 1123–1134. [Google Scholar] [CrossRef]
- Kramer, D.L.; Bryant, M.J. Intestine length in the fishes of a tropical stream: 2. Relationships to diet—The long and short of a convoluted issue. Environ. Biol. Fishes 1995, 42, 129–141. [Google Scholar] [CrossRef]
- Esteban, M.A.; Cuesta, A.; Ortuño, J.; Meseguer, J. Immunomodulatory effects of dietary intake of chitin on gilthead seabream (Sparus aurata L.) innate immune system. Fish Shellfish Immunol. 2001, 11, 303–315. [Google Scholar] [CrossRef] [PubMed]
- Veldkamp, T.; Dong, L.; Paul, A.; Govers, C. Bioactive properties of insect products for monogastric animals—A review. J. Insects Food Feed 2021, 8, 1027–1040. [Google Scholar] [CrossRef]
- Kumar, V.; Fawole, F.J.; Romano, N.; Hossain, M.S.; Labh, S.N.; Overturf, K.; Small, B.C. Insect (black soldier fly, Hermetia illucens) meal supplementation prevents the soybean meal-induced intestinal enteritis in rainbow trout and health benefits of using insect oil. Fish Shellfish Immunol. 2021, 109, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Iger, Y.; Abraham, M. Rodlet cells in the epidermis of fish exposed to stressors. Tissue Cell 1997, 29, 431–438. [Google Scholar] [CrossRef]
- Ross, N.W.; Firth, K.J.; Wang, A.; Burka, J.F.; Johnson, S.C. Changes in hydrolytic enzyme activities of naïve Atlantic salmon Salmo salar skin mucus due to infection with the salmon louse Lepeophtheirus salmonis and cortisol implantation. Dis. Aquat. Org. 2000, 41, 43–51. [Google Scholar] [CrossRef]
- Moreno, I.M.; Mate, A.; Repetto, G.; Vázquez, C.M.; Cameán, A.M. Influence of microcystin-LR on the activity of membrane enzymes in rat intestinal mucosa. J. Physiol. Biochem. 2003, 59, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Molina, R.; Moreno, I.; Pichardo, S.; Jos, A.; Moyano, R.; Monterde, J.G.; Cameán, A. Acid and alkaline phosphatase activities and pathological changes induced in tilapia fish (Oreochromis sp.) exposed subchronically to microcystins from toxic cyanobacterial blooms under laboratory conditions. Toxicon 2005, 46, 725–735. [Google Scholar] [CrossRef] [PubMed]
- Bruni, L.; Husein, Y.; Secci, G.; Tulli, F.; Parisi, G. Rainbow trout (Oncorhynchus mykiss) skin as potential n-3 fatty acid source. Waste Biomass Valoriz. 2021, 12, 5665–5673. [Google Scholar] [CrossRef]
- Zhou, Z.; Ringø, E.; Olsen, R.E.; Song, S.K. Dietary effects of soybean products on gut microbiota and immunity of aquatic animals: A review. Aquac. Nutr. 2018, 24, 644–665. [Google Scholar] [CrossRef]
- Agboola, J.O.; Mensah, D.D.; Hansen, J.Ø.; Lapeña, D.; Mydland, L.T.; Arntzen, M.Ø.; Horn, S.J.; Øyås, O.; Press, C.M.; Øverland, M. Effects of yeast species and processing on intestinal health and transcriptomic profiles of Atlantic salmon (Salmo salar) fed soybean meal-based diets in seawater. Int. J. Mol. Sci. 2022, 23, 1675. [Google Scholar] [CrossRef]
- Xu, X.; Ji, H.; Belghit, I.; Liland, N.S.; Wu, W.; Li, X. Effects of black soldier fly oil rich in n-3 HUFA on growth performance, metabolism and health response of juvenile mirror carp (Cyprinus carpio var. specularis). Aquaculture 2021, 533, 736144. [Google Scholar] [CrossRef]
- Løvmo, S.D.; Sundh, H.; Whatmore, P.; Nordvi, M.F.; Sigholt, T.; Madato, A.; Bardal, T.; Olsen, R.E. Intestinal health in Atlantic salmon post-smolt (Salmo salar) when fed low and high HUFA diets. Aquaculture 2022, 557, 738318. [Google Scholar] [CrossRef]
- Basto, A.; Calduch-Giner, J.; Oliveira, B.; Petit, L.; Sá, T.; Maia, M.R.G.; Fonseca, S.C.; Matos, E.; Pérez-Sánchez, J.; Valente, L.M.P. The use of defatted Tenebrio molitor larvae meal as a main protein source is supported in European sea bass (Dicentrarchus labrax) by data on growth performance, lipid metabolism, and flesh quality. Front. Physiol. 2021, 12, 659567. [Google Scholar] [CrossRef]
- Józefiak, A.; Nogales-Mérida, S.; Rawski, M.; Kierończyk, B.; Mazurkiewicz, J. Effects of insect diets on the gastrointestinal tract health and growth performance of Siberian sturgeon (Acipenser baerii Brandt, 1869). BMC Vet. Res. 2019, 15, 348. [Google Scholar] [CrossRef]
- Mikołajczak, Z.; Rawski, M.; Mazurkiewicz, J.; Kierończyk, B.; Józefiak, D. The Effect of hydrolyzed insect meals in sea trout fingerling (Salmo trutta m. trutta) diets on growth performance, microbiota and biochemical blood parameters. Animals 2020, 10, 1031. [Google Scholar] [CrossRef]
- Zarantoniello, M.; Randazzo, B.; Gioacchini, G.; Truzzi, C.; Giorgini, E.; Riolo, P.; Gioia, G.; Bertolucci, C.; Osimani, A.; Cardinaletti, G.; et al. Zebrafish (Danio rerio) physiological and behavioural responses to insect-based diets: A multidisciplinary approach. Sci. Rep. 2020, 10, 10648. [Google Scholar] [CrossRef] [PubMed]
- Gaudioso, G.; Marzorati, G.; Faccenda, F.; Weil, T.; Lunelli, F.; Cardinaletti, G.; Marino, G.; Olivotto, I.; Parisi, G.; Tibaldi, E.; et al. Processed animal proteins from insect and poultry by-products in a fish meal-free diet for rainbow trout: Impact on intestinal microbiota and inflammatory markers. Int. J. Mol. Sci. 2021, 22, 5454. [Google Scholar] [CrossRef] [PubMed]
- Rimoldi, S.; Antonini, M.; Gasco, L.; Moroni, F.; Terova, G. Intestinal microbial communities of rainbow trout (Oncorhynchus mykiss) may be improved by feeding a Hermetia illucens meal/low-fishmeal diet. Fish Physiol. Biochem. 2021, 47, 365–380. [Google Scholar] [CrossRef] [PubMed]
- Tran, H.Q.; Prokešová, M.; Zare, M.; Gebauer, T.; Elia, A.C.; Colombino, E.; Ferrocino, I.; Caimi, C.; Gai, F.; Gasco, L.; et al. How does pikeperch Sander lucioperca respond to dietary insect meal Hermetia illucens? Investigation on gut microbiota, histomorphology, and antioxidant biomarkers. Front. Mar. Sci. 2021, 8, 680942. [Google Scholar] [CrossRef]
- Rimoldi, S.; Gini, E.; Iannini, F.; Gasco, L.; Terova, G. The effects of dietary insect meal from Hermetia illucens prepupae on autochthonous gut microbiota of rainbow trout (Oncorhynchus mykiss). Animals 2019, 9, 143. [Google Scholar] [CrossRef]
- Biasato, I.; Chemello, G.; Oddon, S.B.; Ferrocino, I.; Corvaglia, M.R.; Caimi, C.; Resconi, A.; Paul, A.; van Spankeren, M.; Capucchio, M.T.; et al. Hermetia illucens meal inclusion in low-fishmeal diets for rainbow trout (Oncorhynchus mykiss): Effects on the growth performance, nutrient digestibility coefficients, selected gut health traits, and health status indices. Anim. Feed Sci. Technol. 2022, 290, 115341. [Google Scholar] [CrossRef]
- Terova, G.; Gini, E.; Gasco, L.; Moroni, F.; Antonini, M.; Rimoldi, S. Effects of full replacement of dietary fishmeal with insect meal from Tenebrio molitor on rainbow trout gut and skin microbiota. J. Anim. Sci. Biotechnol. 2021, 12, 30. [Google Scholar] [CrossRef]
- Antonopoulou, E.; Nikouli, E.; Piccolo, G.; Gasco, L.; Gai, F.; Chatzifotis, S.; Mente, E.; Kormas, K.A. Reshaping gut bacterial communities after dietary Tenebrio molitor larvae meal supplementation in three fish species. Aquaculture 2019, 503, 628–635. [Google Scholar] [CrossRef]
- Piazzon, M.C.; Naya-Català, F.; Pereira, G.V.; Estensoro, I.; del Pozo, R.; Calduch-Giner, J.A.; Nuez-Ortín, W.G.; Palenzuela, O.; Sitjà-Bobadilla, A.; Dias, J.; et al. A novel fish meal-free diet formulation supports proper growth and does not impair intestinal parasite susceptibility in gilthead sea bream (Sparus aurata) with a reshape of gut microbiota and tissue-specific gene expression patterns. Aquaculture 2022, 558, 738362. [Google Scholar] [CrossRef]
- Chang, C.-I.; Liu, W.-Y. An evaluation of two probiotic bacterial strains, Enterococcus faecium SF68 and Bacillus toyoi, for reducing edwardsiellosis in cultured European eel, Anguilla anguilla L. J. Fish Dis. 2002, 25, 311–315. [Google Scholar] [CrossRef]
- Capkin, E.; Altinok, I. Effects of dietary probiotic supplementations on prevention⁄treatment of yersiniosis disease. J. Appl. Microbiol. 2009, 106, 1147–1153. [Google Scholar] [CrossRef] [PubMed]
- Dimitroglou, A.; Merrifield, D.L.; Carnevali, O.; Picchietti, S.; Avella, M.; Daniels, C.; Güroy, D.; Davies, S.J. Microbial manipulations to improve fish health and production—A Mediterranean perspective. Fish Shellfish Immunol. 2011, 30, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Józefiak, A.; Nogales-Mérida, S.; Mikołajczak, Z.; Rawski, M.; Kierończyk, B.; Mazurkiewicz, J. The utilization of full-fat insect meal in rainbow trout (Oncorhynchus mykiss) nutrition: The effects on growth performance, intestinal microbiota and gastrointestinal tract histomorphology. Ann. Anim. Sci. 2019, 19, 747–765. [Google Scholar] [CrossRef]
- Smith, E.A.; Macfarlane, G.T. Enumeration of human colonic bacteria producing phenolic and indolic compounds: Effects of pH, carbohydrate availability and retention time on dissimilatory aromatic amino acid metabolism. J. Appl. Bacteriol. 1996, 81, 288–302. [Google Scholar] [CrossRef]
- Neis, E.P.J.G.; Dejong, C.H.C.; Rensen, S.S. The role of microbial amino acid metabolism in host metabolism. Nutrients 2015, 7, 2930–2946. [Google Scholar] [CrossRef]
- Chapagain, P.; Arivett, B.; Cleveland, B.M.; Walker, D.M.; Salem, M. Analysis of the fecal microbiota of fast- and slow-growing rainbow trout (Oncorhynchus mykiss). BMC Genom. 2019, 20, 788. [Google Scholar] [CrossRef]
- Oren, A.; Garrity, G.M. Valid publication of the names of forty-two phyla of prokaryotes. Int. J. Syst. Evol. Microbiol. 2021, 71, 005056. [Google Scholar] [CrossRef]



| Ingredients (%; on dry basis) | C | T | dT | TO1 | TO2 |
| Fishmeal LT94 1 | 36.78 | 18.28 | 18.48 | 18.28 | 18.28 |
| Yellow mealworm meal 2 | - | 19.05 | - | 19.05 | 19.05 |
| Defatted yellow mealworm meal 3 | - | - | 18.29 | - | - |
| Enriched omega–3 oil | - | - | - | 3.09 | 7.24 |
| Wheat gluten 4 | 11.05 | 12.41 | 10.65 | 12.41 | 12.41 |
| Soybean protein concentrate 5 | 15.09 | 16.4 | 15.41 | 16.4 | 16.4 |
| Wheat meal 6 | 16.16 | 16.98 | 16.19 | 16.98 | 16.98 |
| Soybean lecithin 7 | 1.27 | 0.48 | 1.27 | 0.48 | 0.48 |
| Fish oil 8 | 11.87 | 8.68 | 11.9 | 5.6 | 1.45 |
| Vitamin and mineral premix 9 | 1.95 | 1.93 | 1.95 | 1.93 | 1.93 |
| Goma guar 10 | 1.95 | 1.93 | 1.95 | 1.93 | 1.93 |
| Blood meal 10 | 3.89 | 3.86 | 3.90 | 3.86 | 3.86 |
| Methionine 10 | 0.2 | 0.5 | 0.5 | 0.5 | 0.5 |
| Lysine 10 | - | 0.1 | 0.1 | 0.1 | 0.1 |
| Proximate composition (%; on dry basis) | C | T | dT | TO1 | TO2 |
| Crude protein | 49.19 | 48.76 | 48.98 | 49.25 | 48.23 |
| Crude lipid | 17.80 | 18.58 | 17.86 | 18.99 | 19.12 |
| Crude fibre | 0.97 | 2.15 | 2.35 | 2.25 | 2.47 |
| Ash | 8.63 | 8.49 | 6.31 | 6.22 | 6.12 |
| Calcium (Ca) | 0.43 | 0.24 | 0.17 | 0.18 | 0.20 |
| Phosphorus (P) | 0.31 | 0.24 | 0.24 | 0.24 | 0.24 |
| Ca:P ratio | 1.38 | 1.0 | 0.73 | 0.77 | 0.86 |
| Amino Acid Composition (g/100 g Wet Feed) | C | T | dT | TO1 | TO2 |
|---|---|---|---|---|---|
| Essential amino acids | |||||
| Arg | 2.10 | 2.35 | 2.08 | 2.01 | 2.00 |
| His | 1.09 | 1.02 | 1.05 | 1.05 | 1.04 |
| Ile | 1.50 | 1.69 | 1.45 | 1.56 | 1.44 |
| Leu | 3.00 | 2.99 | 3.02 | 2.90 | 2.88 |
| Lys | 2.94 | 3.66 | 2.77 | 2.70 | 2.66 |
| Met | 1.01 | 1.15 | 1.23 | 1.18 | 1.13 |
| Phe | 2.07 | 2.19 | 2.01 | 1.96 | 1.95 |
| Thr | 1.48 | 1.56 | 1.48 | 1.31 | 1.36 |
| Val | 2.10 | 2.07 | 2.16 | 2.25 | 2.10 |
| Non-essential amino acids | |||||
| Ala | 2.01 | 2.06 | 2.18 | 1.95 | 1.94 |
| Asp | 3.32 | 3.56 | 3.35 | 3.10 | 3.12 |
| Cys | 0.46 | 0.66 | 0.46 | 0.43 | 0.44 |
| Glu | 7.15 | 7.64 | 7.41 | 7.44 | 7.44 |
| Gly | 1.92 | 2.20 | 1.91 | 1.81 | 1.80 |
| Pro | 2.46 | 2.51 | 2.67 | 2.74 | 2.61 |
| Ser | 1.76 | 1.87 | 1.91 | 1.65 | 1.79 |
| Tyr | 1.63 | 1.78 | 2.94 | 2.12 | 2.42 |
| Fatty Acid Composition (g/100 Total Fatty Acids) | C | T | dT | TO1 | TO2 |
|---|---|---|---|---|---|
| MYR; C14:0 | 3.68 | 3.26 | 2.55 | 3.16 | 3.20 |
| PA; C16:0 | 21.13 | 19.37 | 17.35 | 20.75 | 22.74 |
| POA; C16:1n–7 | 4.68 | 2.97 | 3.56 | 3.02 | 2.00 |
| STE; C18:0 | 5.67 | 4.78 | 4.96 | 3.95 | 2.87 |
| OA; C18:1n–9 | 13.67 | 23.05 | 16.17 | 20.47 | 17.14 |
| VA; C18:1n–7 | 2.60 | 1.54 | 1.42 | 0.00 | 0.00 |
| LA; C18:2n–6 | 8.77 | 18.69 | 13.75 | 15.31 | 15.21 |
| ALA; C18:3n–3 | 1.12 | 1.33 | 1.56 | 0.95 | 0.87 |
| SDA; C18:4n–3 | 0.82 | 0.44 | 0.89 | 0.61 | 0.51 |
| GOA; C20:1n–9 | 0.98 | 0.76 | 1.75 | 1.14 | 0.56 |
| ARA; C20:4n–6 | 1.66 | 1.22 | 1.56 | 1.17 | 1.26 |
| EPA; C20:5n–3 | 8.31 | 3.92 | 6.42 | 6.33 | 8.51 |
| DPA; C22:5n–3 | 1.33 | 0.89 | 1.49 | 1.08 | 1.12 |
| DHA; C22:6n–3 | 18.79 | 14.05 | 19.07 | 16.10 | 19.20 |
| Other (up to 100%) | 6.79 | 3.73 | 7.50 | 5.96 | 4.81 |
| ∑SFA | 30.48 | 27.41 | 24.87 | 27.86 | 28.81 |
| ∑MUFA | 21.93 | 28.32 | 22.91 | 24.63 | 19.70 |
| ∑PUFA | 40.80 | 40.54 | 44.73 | 41.56 | 46.67 |
| ∑n–3 | 30.37 | 20.63 | 29.42 | 25.07 | 30.20 |
| ∑n–6 | 10.43 | 19.91 | 15.31 | 16.49 | 16.47 |
| ∑n–6/∑n–3 | 0.34 | 0.97 | 0.52 | 0.66 | 0.55 |
| ∑Total fatty acids | 100 | 100 | 100 | 100 | 100 |
| Growth performance | C | T | dT | TO1 | TO2 | SEM | p-value | F-value | DF |
| IBW (g) | 46.11 | 46.25 | 46.08 | 46.2 | 46.05 | 0.11 | 0.6461 | 0.92 | 4 |
| Final length (cm) | 29.94 | 29.86 | 30.06 | 30.01 | 29.84 | 0.12 | 0.6787 | 0.58 | 4 |
| SGR (%/day) | 2.47 | 2.46 | 2.48 | 2.47 | 2.46 | 0.01 | 0.592 | 0.72 | 4 |
| FCR | 0.86 | 0.85 | 0.86 | 0.85 | 0.85 | 0.005 | 0.4568 | 0.96 | 4 |
| Protein utilization | C | T | dT | TO1 | TO2 | SEM | p-value | F-value | DF |
| PER | 2.63 ab | 2.68 a | 2.6 b | 2.64 ab | 2.67 a | 0.02 | 0.0142 | 4.47 | 4 |
| PPV (%) | 46.89 | 48.83 | 46.66 | 47.05 | 47.1 | 0.68 | 0.2165 | 1.64 | 4 |
| ADCprot (%) | 88.31 b | 88.86 b | 88.18 b | 90.5 a | 91.1 a | 0.22 | <0.0001 | 36.07 | 4 |
| Biometric indices | C | T | dT | TO1 | TO2 | SEM | p-value | F-value | DF |
| CF (g/cm3) | 1.55 | 1.55 | 1.54 | 1.55 | 1.55 | 0.02 | 0.9407 | 0.19 | 4 |
| HSI (%) | 1.33 | 1.3 | 1.2 | 1.17 | 1.14 | 0.15 | 0.1209 | 2.18 | 4 |
| VSI (%) | 11.14 | 10.45 | 10.02 | 10.13 | 9.94 | 0.36 | 0.1652 | 1.89 | 4 |
| ISI (%) | 4.58 a | 4.3 abc | 4.33 ab | 3.82 c | 3.99 bc | 0.11 | 0.0021 | 7.09 | 4 |
| IL/FL (%) | 70.28 | 66.07 | 67.16 | 65 | 64.21 | 1.69 | 0.1524 | 1.96 | 4 |
| Butchering yield | C | T | dT | TO1 | TO2 | SEM | p-value | F-value | DF |
| Dressing yield (%) | 89.95 | 90.6 | 90.54 | 90.87 | 91.28 | 0.32 | 0.0969 | 2.39 | 4 |
| Filleting yield (%) | 57.91 | 58.73 | 59.5 | 60.3 | 60.16 | 0.59 | 0.0597 | 2.87 | 4 |
| Immunological system (plasma) | C | T | dT | TO1 | TO2 | SEM | p-value | F-value | DF |
| Lysozyme | 2.22 | 2.22 | 1.99 | 1.68 | 2.62 | 0.21 | 0.0726 | 2.68 | 4 |
| Antiprotease | 163.75 | 168.86 | 164.46 | 164.12 | 130.77 | 9.21 | 0.0605 | 2.86 | 4 |
| Acid phosphatase | 973.55 | 919.88 | 746.49 | 1019.55 | 926.23 | 80.57 | 0.2133 | 1.65 | 4 |
| Alkaline phosphatase | 1040.85 | 834.1 | 1054.88 | 1036.89 | 930.7 | 63.17 | 0.1097 | 2.27 | 4 |
| Peroxidase | 0.82 | 0.94 | 0.76 | 0.81 | 0.76 | 0.07 | 0.3856 | 1.12 | 4 |
| Immunoglobulins | 17.48 | 18.18 | 16.19 | 15.1 | 16.25 | 0.86 | 0.1558 | 1.94 | 4 |
| Immunological system (skin mucus) | C | T | dT | TO1 | TO2 | SEM | p-value | F-value | DF |
| Acid phosphatase | 1256.02 b | 1095.21 b | 1376.64 b | 2471.43 a | 1642.04 ab | 207.11 | 0.0024 | 6.87 | 4 |
| Alkaline phosphatase | 1286.65 b | 2196.06 ab | 2781.82 ab | 3320.95 a | 2694.87 ab | 431.58 | 0.0457 | 3.15 | 4 |
| Peroxidase | 16.83 | 23.09 | 13.85 | 20.41 | 11.52 | 3.49 | 0.1771 | 1.82 | 4 |
| Esterase | 11.91 | 13.03 | 9.67 | 11.7 | 9.44 | 1.24 | 0.2426 | 1.53 | 4 |
| Carbonic anhydrase | 685.83 | 485.73 | 429.89 | 131.3 | 460.53 | 153.97 | 0.1608 | 1.96 | 4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Melenchón, F.; Larrán, A.M.; Hernández, M.; Abad, D.; Morales, A.E.; Pula, H.J.; Fabrikov, D.; Sánchez-Muros, M.J.; Galafat, A.; Alarcón, F.J.; et al. Different Diets Based on Yellow Mealworm (Tenebrio molitor)—Part B: Modification of the Intestinal Inflammatory Response and the Microbiota Composition of Rainbow Trout (Oncorhynchus mykiss). Fishes 2023, 8, 284. https://doi.org/10.3390/fishes8060284
Melenchón F, Larrán AM, Hernández M, Abad D, Morales AE, Pula HJ, Fabrikov D, Sánchez-Muros MJ, Galafat A, Alarcón FJ, et al. Different Diets Based on Yellow Mealworm (Tenebrio molitor)—Part B: Modification of the Intestinal Inflammatory Response and the Microbiota Composition of Rainbow Trout (Oncorhynchus mykiss). Fishes. 2023; 8(6):284. https://doi.org/10.3390/fishes8060284
Chicago/Turabian StyleMelenchón, Federico, Ana María Larrán, Marta Hernández, David Abad, Amalia E. Morales, Héctor J. Pula, Dmitri Fabrikov, María José Sánchez-Muros, Alba Galafat, Francisco Javier Alarcón, and et al. 2023. "Different Diets Based on Yellow Mealworm (Tenebrio molitor)—Part B: Modification of the Intestinal Inflammatory Response and the Microbiota Composition of Rainbow Trout (Oncorhynchus mykiss)" Fishes 8, no. 6: 284. https://doi.org/10.3390/fishes8060284