Molecular Characterization and Expression Response of Ghrelin, GLP-1 and PYY to Fasting, Dietary Lipid, and Fatty Acids in Silver Pomfret (Pampus argenteus)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Fish
2.2. Open Reading Frame (ORF) Cloning of Ghrelin, glp-1, pyya, and pyyb
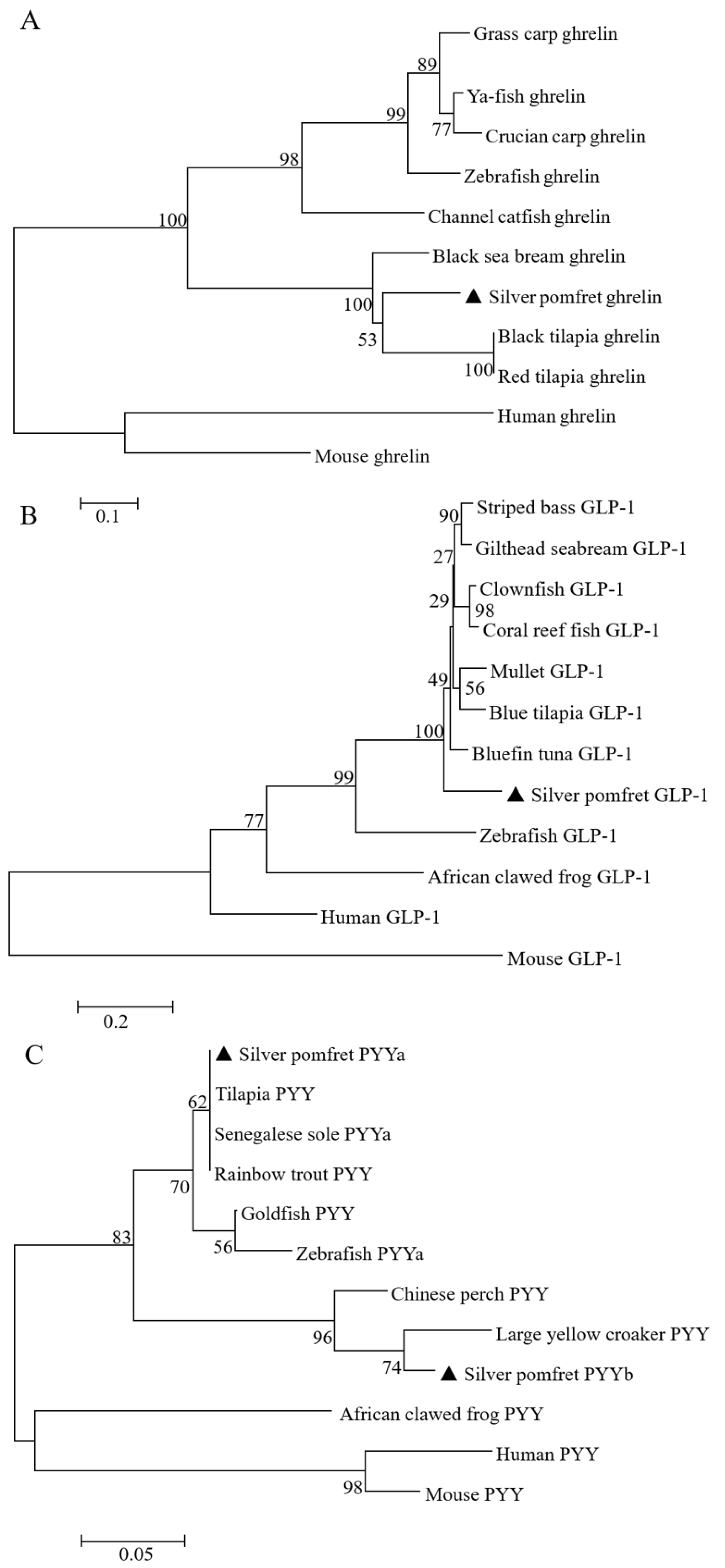
2.3. Sequences Analysis
2.4. Feeding Experiment
2.5. Lipid Concentrition Experiment
2.6. FAs Incubation
2.7. Quantitative Real-Time PCR (qPCR)
2.8. Data Analysis and Statistics
3. Results
3.1. Molecular Characterization of Silver Pomfret Ghrelin, glp-1, pyya, and pyyb
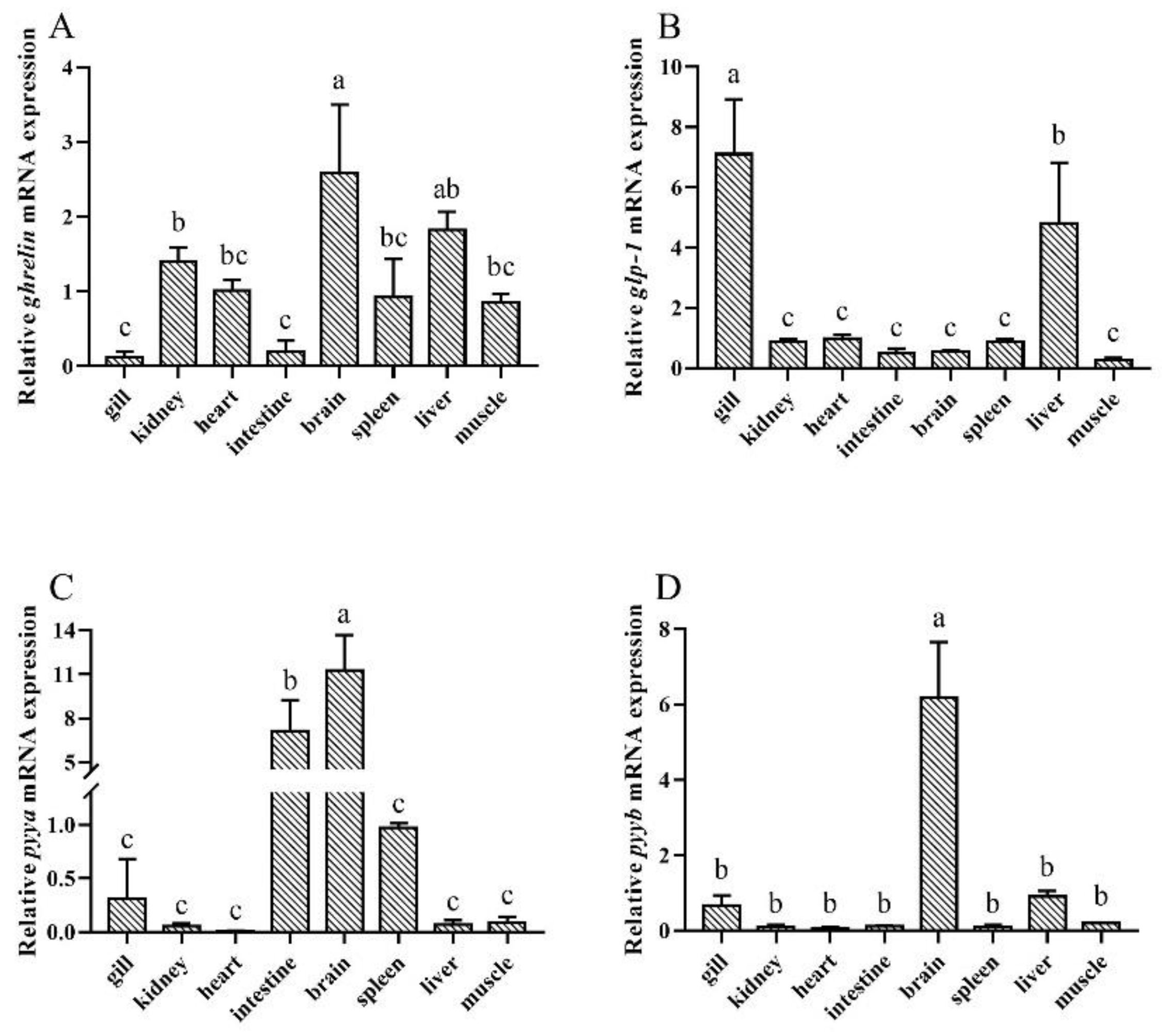
3.2. Tissue Distribution of Silver Pomfret Ghrelin, glp-1, pyya, and pyyb
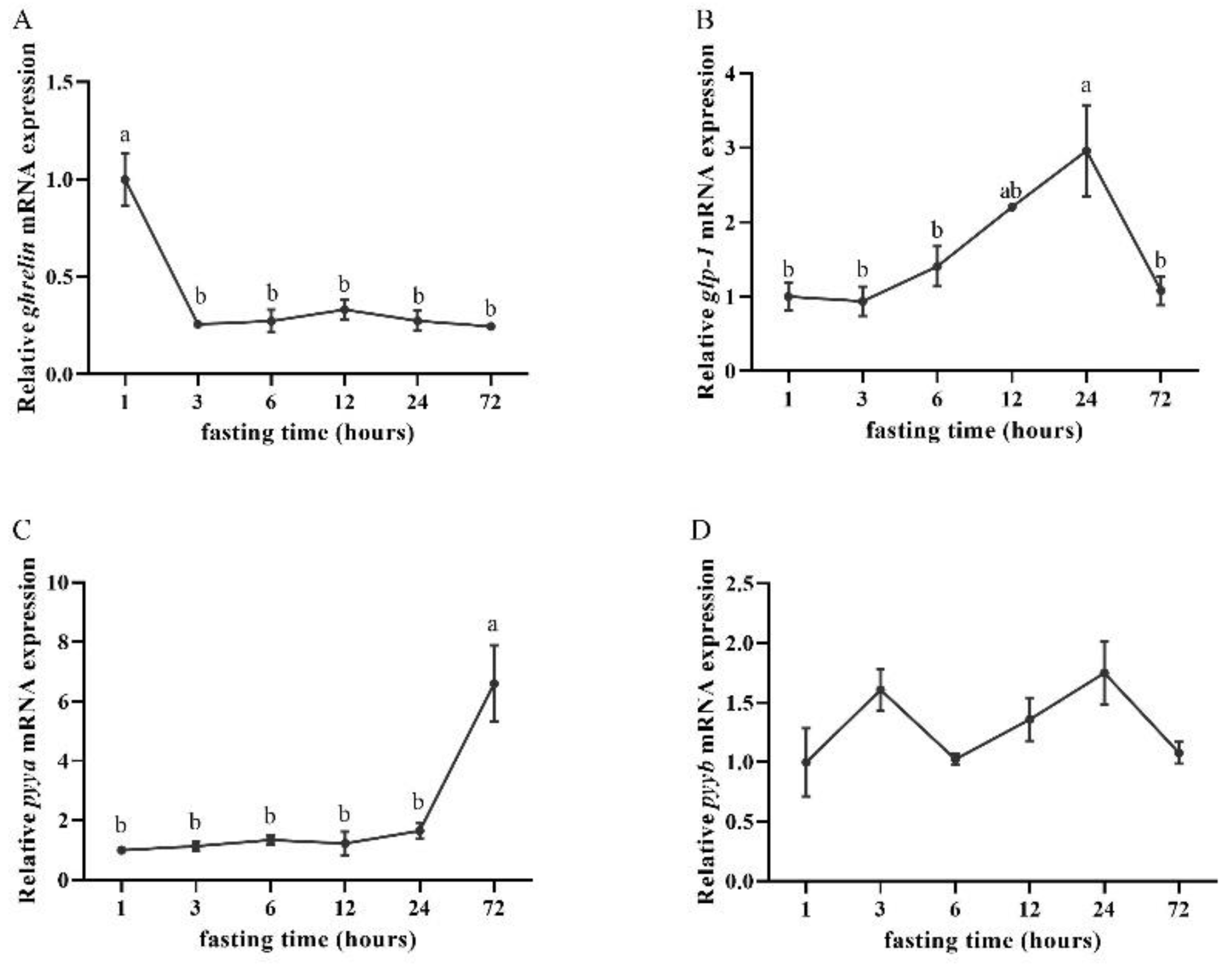
3.3. Effects of Fasting on Ghrelin, glp-1, pyya, and pyyb Expression
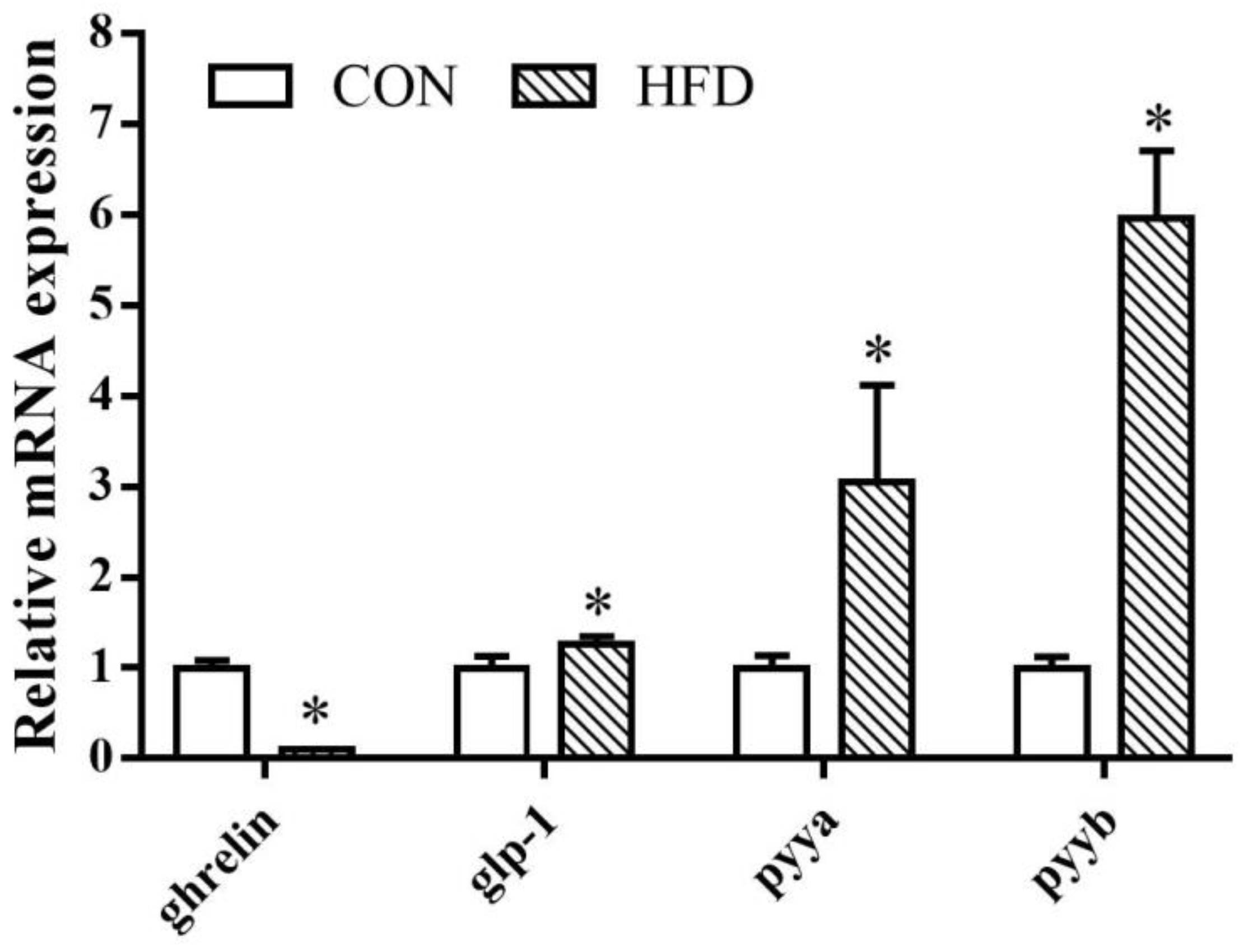
3.4. Effects of Dietary Lipid Concentration on Ghrelin, glp-1, pyya, and pyyb Expression
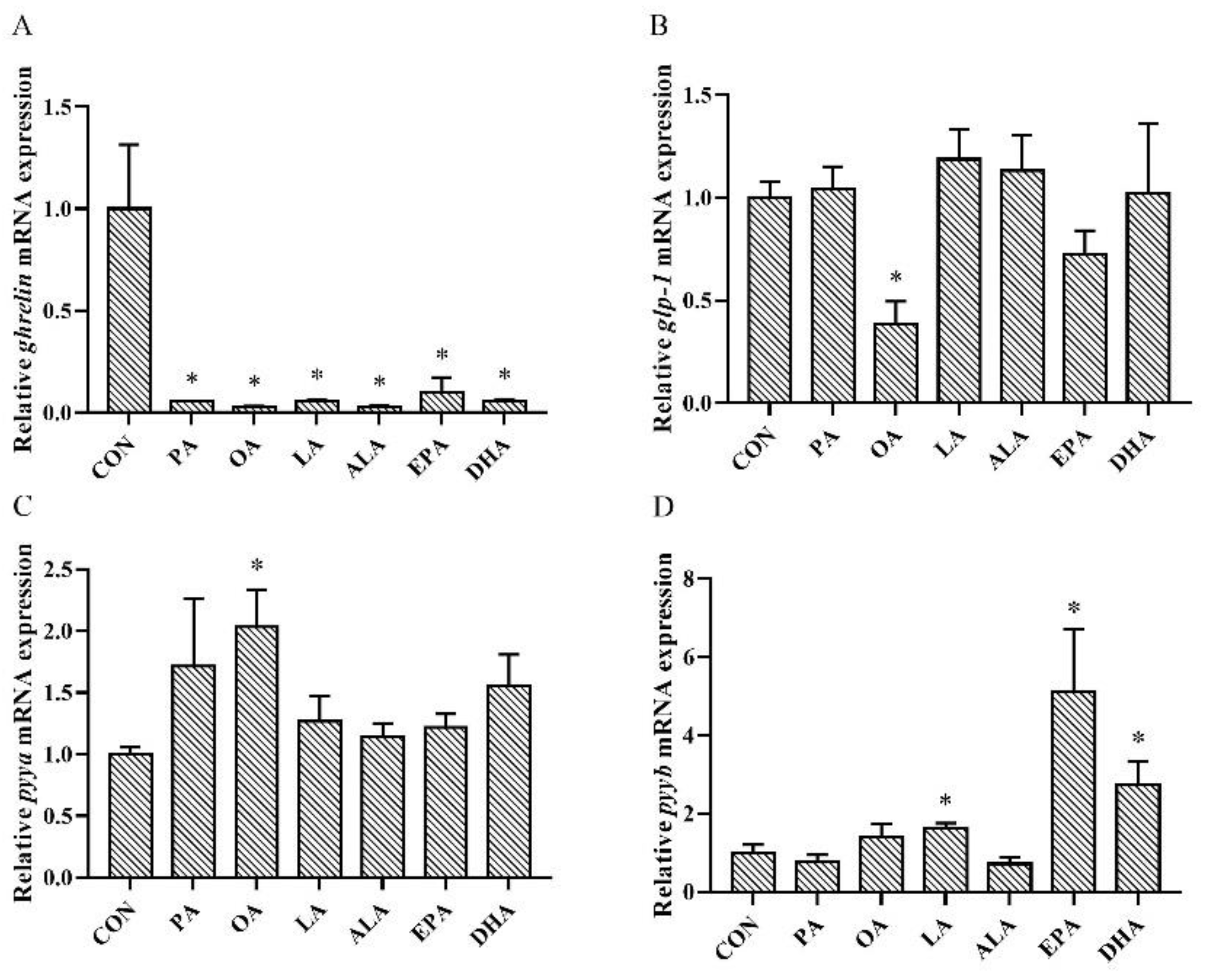
3.5. Effects of FAs Incubation on Ghrelin, glp-1, pyya, and pyyb Expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rehfeld, J.F. The New Biology of Gastrointestinal Hormones. Physiol. Rev. 1998, 78, 1087–1108. [Google Scholar] [CrossRef]
- Gribble, F.M.; Reimann, F. Function and Mechanisms of Enteroendocrine Cells and Gut Hormones in Metabolism. Nat. Rev. Endocrinol. 2019, 15, 226–237. [Google Scholar] [CrossRef] [PubMed]
- Delfini, M.; Stakenborg, N.; Viola, M.F.; Boeckxstaens, G. Macrophages in the Gut: Masters in Multitasking. Immunity 2022, 55, 1530–1548. [Google Scholar] [CrossRef] [PubMed]
- McCarty, T.R.; Jirapinyo, P.; Thompson, C.C. Effect of Sleeve Gastrectomy on Ghrelin, GLP-1, PYY, and GIP Gut Hormones: A Systematic Review and Meta-Analysis. Ann. Surg. 2020, 272, 72. [Google Scholar] [CrossRef] [PubMed]
- Freire, R.H.; Alvarez-Leite, J.I. Appetite Control: Hormones or Diet Strategies? Curr. Opin. Clin. Nutr. Metab. Care 2020, 23, 328–335. [Google Scholar] [CrossRef] [PubMed]
- Kojima, M.; Hosoda, H.; Date, Y.; Nakazato, M.; Matsuo, H.; Kangawa, K. Ghrelin Is a Growth-Hormone-Releasing Acylated Peptide from Stomach. Nature 1999, 402, 656–660. [Google Scholar] [CrossRef]
- Nunez-Salces, M.; Li, H.; Feinle-Bisset, C.; Young, R.L.; Page, A.J. The Regulation of Gastric Ghrelin Secretion. Acta Physiol. 2021, 231, e13588. [Google Scholar] [CrossRef]
- Zhong, H.; Hu, Y.; Yu, F. A Review on Ghrelin and Fish Reproduction. Reprod. Breed. 2021, 1, 128–135. [Google Scholar] [CrossRef]
- Del Vecchio, G.; Lai, F.; Gomes, A.S.; Verri, T.; Kalananthan, T.; Barca, A.; Handeland, S.; Rønnestad, I. Effects of Short-Term Fasting on MRNA Expression of Ghrelin and the Peptide Transporters PepT1 and 2 in Atlantic Salmon (Salmo Salar). Front. Physiol. 2021, 12, 666670. [Google Scholar] [CrossRef]
- Calo, J.; Soengas, J.L.; Pastor, J.J.; Blanco, A.M.; Morais, S. Evidence of Gastrointestinal Sensing and Gut-Brain Communication in Rainbow Trout (Oncorhynchus mykiss) in Response to the Aqueous Extract of Fishmeal and Its Free Amino Acid Fraction. Aquaculture 2023, 567, 739264. [Google Scholar] [CrossRef]
- Panaro, B.L.; Yusta, B.; Matthews, D.; Koehler, J.A.; Song, Y.; Sandoval, D.A.; Drucker, D.J. Intestine-Selective Reduction of Gcg Expression Reveals the Importance of the Distal Gut for GLP-1 Secretion. Mol. Metab. 2020, 37, 100990. [Google Scholar] [CrossRef] [PubMed]
- Hira, T.; Pinyo, J.; Hara, H. What Is GLP-1 Really Doing in Obesity? Trends Endocrinol. Metab. 2020, 31, 71–80. [Google Scholar] [CrossRef]
- Xie, Y.; Cheng, D.; Wang, L.; Chen, F.; Chen, H.; Ma, H.; Yang, Y.; Yuan, X. GLP-1 Responds to Postprandial Hyperglycemia by Reducing Transcription Level in Grass Carp (Ctenopharyngodon idella). Aquac. Rep. 2022, 23, 101045. [Google Scholar] [CrossRef]
- White, S.L.; Volkoff, H.; Devlin, R.H. Regulation of Feeding Behavior and Food Intake by Appetite-Regulating Peptides in Wild-Type and Growth Hormone-Transgenic Coho Salmon. Horm. Behav. 2016, 84, 18–28. [Google Scholar] [CrossRef] [PubMed]
- Assan, D.; Mustapha, U.F.; Ndandala, C.B.; Li, Z.; Li, G.; Huang, C.; Zhao, G.; Chen, H.; Jiang, M. Molecular Cloning, Identification, and Expression Analysis of Pyy and Its Effect on Mechanisms Regulating Appetite in Spotted Scat, Scatophagus argus. Aquac. Rep. 2023, 28, 101465. [Google Scholar] [CrossRef]
- Assan, D.; Mustapha, U.F.; Chen, H.; Li, Z.; Peng, Y.; Li, G. The Roles of Neuropeptide Y (Npy) and Peptide YY (Pyy) in Teleost Food Intake: A Mini Review. Life 2021, 11, 547. [Google Scholar] [CrossRef]
- Nadermann, N.; Seward, R.K.; Volkoff, H. Effects of Potential Climate Change-Induced Environmental Modifications on Food Intake and the Expression of Appetite Regulators in Goldfish. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2019, 235, 138–147. [Google Scholar] [CrossRef]
- Gomes, A.S.; Lygre, E.; Harboe, T.; Zimmermann, F.; Jordal, A.-E.O.; Hamre, K.; Rønnestad, I. The Role of Cholecystokinin and Peptide YY in Feed Intake in Atlantic Halibut (Hippoglossus hippoglossus) Larvae. Neuropeptides 2022, 91, 102202. [Google Scholar] [CrossRef]
- Zhou, R.; Wu, G.; Qu, L.; Zhong, X.; Gao, Y.; Ding, Z.; Xu, J.; Chen, X.; Cheng, H. Effect of Starvation on Intestinal Morphology, Digestive Enzyme Activity and Expression of Lipid Metabolism-Related Genes in Javelin Goby (Synechogobius hasta). Aquac. Res. 2022, 53, 87–97. [Google Scholar] [CrossRef]
- Kalananthan, T.; Folkedal, O.; Gomes, A.S.; Lai, F.; Handeland, S.O.; Tolås, I.; Gelebart, V.; Rønnestad, I. Impact of Long-Term Fasting on the Stomach-Hypothalamus Appetite Regulating Genes in Atlantic Salmon Postsmolts. Aquaculture 2023, 563, 738917. [Google Scholar] [CrossRef]
- Zhang, X.; Chen, S.; Tang, N.; Wang, S.; Liu, Y.; Li, Y.; Liu, Y.; Zhang, S.; Li, Y.; Chen, D.; et al. Neuromedin U Regulates Food Intake of Siberian Sturgeon through the Modulation of Central and Peripheral Appetite Factors. Br. J. Nutr. 2023, 129, 904–918. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.-O.; Oh, S.-Y.; Kim, T. Effect of Fasting and Refeeding on Juvenile Leopard Mandarin Fish Siniperca scherzeri. Animals 2022, 12, 889. [Google Scholar] [CrossRef]
- Mankiewicz, J.L.; Cleveland, B.M. Characterization of a Leptin Receptor Paralog and Its Response to Fasting in Rainbow Trout (Oncorhynchus mykiss). Int. J. Mol. Sci. 2021, 22, 7732. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Wang, M.; Li, Y.; Tang, N.; Zhang, X.; Chen, H.; Zhang, S.; Liu, Y.; Wang, J.; Chen, D.; et al. Cloning and Expression of Kiss Genes and Regulation of Feeding in Siberian Sturgeon (Acipenser baerii). Fish Physiol. Biochem. 2022, 48, 419–436. [Google Scholar] [CrossRef] [PubMed]
- Liao, K.; Meng, R.; Ran, Z.; Cheng, G.; Wang, Y.; Xu, J.; Xu, S.; Yan, X. Short-Term Starvation in Silver Pomfret (Pampus argenteus): Molecular Effects on Lipid Mobilization and Utilization. Aquac. Res. 2017, 48, 4874–4885. [Google Scholar] [CrossRef]
- Li, Y.; Liang, S.; Shao, Y.; Li, Y.; Chen, C.; You, C.; Monroig, Ó.; Rahimnejad, S.; Tocher, D.R.; Wang, S. Impacts of Dietary Konjac Glucomannan Supplementation on Growth, Antioxidant Capacity, Hepatic Lipid Metabolism and Inflammatory Response in Golden Pompano (Trachinotus ovatus) Fed a High Fat Diet. Aquaculture 2021, 545, 737113. [Google Scholar] [CrossRef]
- Tang, T.; Hu, Y.; Peng, M.; Chu, W.; Hu, Y.; Zhong, L. Effects of High-Fat Diet on Growth Performance, Lipid Accumulation and Lipid Metabolism-Related MicroRNA/Gene Expression in the Liver of Grass Carp (Ctenopharyngodon idella). Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2019, 234, 34–40. [Google Scholar] [CrossRef]
- Gélineau, A.; Corraze, G.; Boujard, T.; Larroquet, L.; Kaushik, S. Relation between Dietary Lipid Level and Voluntary Feed Intake, Growth, Nutrient Gain, Lipid Deposition and Hepatic Lipogenesis in Rainbow Trout. Reprod. Nutr. Dev. 2001, 41, 487–503. [Google Scholar] [CrossRef]
- López, L.M.; Torres, A.L.; Durazo, E.; Drawbridge, M.; Bureau, D.P. Effects of Lipid on Growth and Feed Utilization of White Seabass (Atractoscion nobilis) Fingerlings. Aquaculture 2006, 253, 557–563. [Google Scholar] [CrossRef]
- Huang, C.; Sun, J.; Ji, H.; Oku, H.; Chang, Z.; Tian, J.; Yu, E.; Xie, J. Influence of Dietary Alpha-Lipoic Acid and Lipid Level on the Growth Performance, Food Intake and Gene Expression of Peripheral Appetite Regulating Factors in Juvenile Grass Carp (Ctenopharyngodon idellus). Aquaculture 2019, 505, 412–422. [Google Scholar] [CrossRef]
- Dai, Y.-J.; Jiang, G.-Z.; Yuan, X.-Y.; Liu, W.-B. High-Fat-Diet-Induced Inflammation Depresses the Appetite of Blunt Snout Bream (Megalobrama amblycephala) through the Transcriptional Regulation of Leptin/Mammalian Target of Rapamycin. Br. J. Nutr. 2018, 120, 1422–1431. [Google Scholar] [CrossRef]
- Hossain, M.A.; Almatar, S.M.; James, C.M. Effect of Varying Dietary Lipid Levels and Protein to Energy (P:E) Ratios on Growth Performance, Feed Utilization and Body Composition of Sub-Adult Silver Pomfrets, Pampus Argenteus (Euphrasen, 1788); Asian Network for Scientific Information: Punjab, Pakistan, 2011. [Google Scholar]
- Calo, J.; Conde-Sieira, M.; Comesaña, S.; Soengas, J.L.; Blanco, A.M. Fatty Acids of Different Nature Differentially Modulate Feed Intake in Rainbow Trout. Aquaculture 2023, 563, 738961. [Google Scholar] [CrossRef]
- Graybeal, A.J.; Shah, M.; Willis, J.L. Manipulation of Fatty Acid Composition in a High-Fat Meal Does Not Result in Differential Alterations in Appetite or Food Intake in Normal Weight Females: A Single-Blind Randomized Crossover Study. Appetite 2021, 160, 105085. [Google Scholar] [CrossRef]
- Behrouz, V.; Yari, Z. A Review on Differential Effects of Dietary Fatty Acids on Weight, Appetite and Energy Expenditure. Crit. Rev. Food Sci. Nutr. 2022, 62, 2235–2249. [Google Scholar] [CrossRef]
- Librán-Pérez, M.; Otero-Rodiño, C.; López-Patiño, M.A.; Míguez, J.M.; Soengas, J.L. Central Administration of Oleate or Octanoate Activates Hypothalamic Fatty Acid Sensing and Inhibits Food Intake in Rainbow Trout. Physiol. Behav. 2014, 129, 272–279. [Google Scholar] [CrossRef]
- Luo, H.; Liang, X.-F.; Li, J.; Zhang, Y.; Xiao, Q.; Peng, B.; Zhang, Z. Effect of Long-Chain Saturated and Unsaturated Fatty Acids on Hypothalamic Fatty Acid Sensing in Chinese Perch (Siniperca chuatsi). Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2020, 241, 110395. [Google Scholar] [CrossRef]
- Lan, W.; Che, X.; Xu, Q.; Wang, T.; Du, R.; Xie, J.; Hou, M.; Lei, H. Sensory and Chemical Assessment of Silver Pomfret (Pampus argenteus) Treated with Ginkgo Biloba Leaf Extract Treatment during Storage in Ice. Aquac. Fish. 2018, 3, 30–37. [Google Scholar] [CrossRef]
- Huang, X.; Yin, Y.; Shi, Z.; Li, W.; Zhou, H.; Lv, W. Lipid Content and Fatty Acid Composition in Wild-Caught Silver Pomfret (Pampus argenteus) Broodstocks: Effects on Gonad Development. Aquaculture 2010, 310, 192–199. [Google Scholar] [CrossRef]
- Hossain, M.A.; Almatar, S.M.; James, C.M. Effects of Varying Dietary Docosahexaenoic Acid Levels on Growth, Proximate Composition and Tissue Fatty Acid Profile of Juvenile Silver Pomfrets, Pampus Argenteus (Euphrasen, 1788). Aquac. Res. 2012, 43, 1599–1610. [Google Scholar] [CrossRef]
- Zhang, Y.; Hu, J.; Li, Y.; Zhang, M.; Jacques, K.J.; Gu, W.; Sun, Y.; Sun, J.; Yang, Y.; Xu, S.; et al. Immune Response of Silver Pomfret (Pampus argenteus) to Amyloodinium Ocellatum Infection. J. Fish Dis. 2021, 44, 2111–2123. [Google Scholar] [CrossRef]
- Xiong, X.; Yang, H.; Ding, C.; Qin, B.; Deng, Y.; Xiong, L.; Liu, X.; Li, Y.; Xiao, T.; Lv, Z. Functional and Expression Analysis Reveals the Involvement of Integrin Avβ3 in Antiviral Immunity of Grass Carp (Ctenopharyngodon idella). Fish Shellfish Immunol. 2022, 129, 52–63. [Google Scholar] [CrossRef]
- Shi, P.; Liao, K.; Xu, J.; Wang, Y.; Xu, S.; Yan, X. Eicosapentaenoic Acid Mitigates Palmitic Acid-Induced Heat Shock Response, Inflammation and Repair Processes in Fish Intestine. Fish Shellfish Immunol. 2022, 124, 362–371. [Google Scholar] [CrossRef] [PubMed]
- Mul, J.D.; Begg, D.P.; Barrera, J.G.; Li, B.; Matter, E.K.; D’Alessio, D.A.; Woods, S.C.; Seeley, R.J.; Sandoval, D.A. High-Fat Diet Changes the Temporal Profile of GLP-1 Receptor-Mediated Hypophagia in Rats. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2013, 305, R68–R77. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Tamura, K.; Nei, M. MEGA3: Integrated Software for Molecular Evolutionary Genetics Analysis and Sequence Alignment. Brief. Bioinform. 2004, 5, 150–163. [Google Scholar] [CrossRef]
- Arias-Jayo, N.; Abecia, L.; Lavín, J.L.; Tueros, I.; Arranz, S.; Ramírez-García, A.; Pardo, M.A. Host-Microbiome Interactions in Response to a High-Saturated Fat Diet and Fish-Oil Supplementation in Zebrafish Adult. J. Funct. Foods 2019, 60, 103416. [Google Scholar] [CrossRef]
- Arlington, V. Official Methods of Analysis of Official Analytical Chemists International; Association of Official Analytical Chemists (AOAC) International: Gaithersburg, MD, USA, 1995. [Google Scholar]
- Blanco, A.M.; Bertucci, J.I.; Valenciano, A.I.; Delgado, M.J.; Unniappan, S. Ghrelin Suppresses Cholecystokinin (CCK), Peptide YY (PYY) and Glucagon-like Peptide-1 (GLP-1) in the Intestine, and Attenuates the Anorectic Effects of CCK, PYY and GLP-1 in Goldfish (Carassius auratus). Horm. Behav. 2017, 93, 62–71. [Google Scholar] [CrossRef]
- Chen, D.; Li, Y.; Wu, H.; Wu, Y.; Tang, N.; Chen, S.; Liu, Y.; Wang, J.; Zhang, X.; Li, Z. Ghrelin-Ghrelin Receptor (GSHR) Pathway via Endocannabinoid Signal Affects the Expression of NPY to Promote the Food Intake of Siberian Sturgeon (Acipenser baerii). Horm. Behav. 2022, 143, 105199. [Google Scholar] [CrossRef] [PubMed]
- Skovgaard, M.; Kodra, J.T.; Gram, D.X.; Knudsen, S.M.; Madsen, D.; Liberles, D.A. Using Evolutionary Information and Ancestral Sequences to Understand the Sequence–Function Relationship in GLP-1 Agonists. J. Mol. Biol. 2006, 363, 977–988. [Google Scholar] [CrossRef]
- Cheng, X.; Pan, M.; E, Z.; Zhou, Y.; Niu, B.; Chen, C. Functional Divergence of Two Duplicated Fertilization Independent Endosperm Genes in Rice with Respect to Seed Development. Plant J. 2020, 104, 124–137. [Google Scholar] [CrossRef]
- Dar, S.A.; Srivastava, P.P.; Varghese, T.; Gupta, S.; Gireesh-Babu, P.; Krishna, G. Effects of Starvation and Refeeding on Expression of Ghrelin and Leptin Gene with Variations in Metabolic Parameters in Labeo Rohita Fingerlings. Aquaculture 2018, 484, 219–227. [Google Scholar] [CrossRef]
- Muta, H.; Sugita, Y.; Furuta, T.; Shiimura, Y.; Ohshima, K.; Nakashima, K.; Sato, K.; Morioka, M.; Abe, H.; Nozawa, T.; et al. Expression of the Ghrelin/Growth Hormone Secretagogue Receptor Axis and Its Functional Role in Promoting Tumor Growth in Primary Central Nervous System Lymphomas. Neuropathology 2020, 40, 232–239. [Google Scholar] [CrossRef]
- Enes, P.; Panserat, S.; Kaushik, S.; Oliva-Teles, A. Nutritional Regulation of Hepatic Glucose Metabolism in Fish. Fish Physiol. Biochem. 2009, 35, 519–539. [Google Scholar] [CrossRef]
- Hindsø, M.; Svane, M.S.; Hedbäck, N.; Holst, J.J.; Madsbad, S.; Bojsen-Møller, K.N. The Role of GLP-1 in Postprandial Glucose Metabolism after Bariatric Surgery: A Narrative Review of Human GLP-1 Receptor Antagonist Studies. Surg. Obes. Relat. Dis. 2021, 17, 1383–1391. [Google Scholar] [CrossRef]
- Liu, Y.; Wei, J.; Luo, Z.; Cui, J.; Luo, Y.; Mak, S.O.K.; Wang, S.; Zhang, F.; Yang, Y.; So, K.-F.; et al. A Gut-Brain Axis Mediates Sodium Appetite via Gastrointestinal Peptide Regulation on a Medulla-Hypothalamic Circuit. Sci. Adv. 2023, 9, eadd5330. [Google Scholar] [CrossRef]
- Chen, Y.; Shen, Y.; Pandit, N.P.; Fu, J.; Li, D.; Li, J. Molecular Cloning, Expression Analysis, and Potential Food Intake Attenuation Effect of Peptide YY in Grass Carp (Ctenopharyngodon idellus). Gen. Comp. Endocrinol. 2013, 187, 66–73. [Google Scholar] [CrossRef]
- Gonzalez, R.; Unniappan, S. Molecular Characterization, Appetite Regulatory Effects and Feeding Related Changes of Peptide YY in Goldfish. Gen. Comp. Endocrinol. 2010, 166, 273–279. [Google Scholar] [CrossRef]
- Unniappan, S.; Canosa, L.F.; Peter, R.E. Orexigenic Actions of Ghrelin in Goldfish: Feeding-Induced Changes in Brain and Gut MRNA Expression and Serum Levels, and Responses to Central and Peripheral Injections. Neuroendocrinology 2004, 79, 100–108. [Google Scholar] [CrossRef]
- Hevrøy, E.M.; Azpeleta, C.; Shimizu, M.; Lanzén, A.; Kaiya, H.; Espe, M.; Olsvik, P.A. Effects of Short-Term Starvation on Ghrelin, GH-IGF System, and IGF-Binding Proteins in Atlantic Salmon. Fish Physiol. Biochem. 2011, 37, 217–232. [Google Scholar] [CrossRef]
- Cai, W.; Yuan, X.; Yuan, Y.; Xie, S.; Gong, Y.; Su, H.; Qiao, Y. Sequence, Genomic Organization and Expression of Ghrelin Receptor in Grass Carp, Ctenopharyngodon idellus. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2015, 179, 54–61. [Google Scholar] [CrossRef]
- D’Alessio, D.; Lu, W.; Sun, W.; Zheng, S.; Yang, Q.; Seeley, R.; Woods, S.C.; Tso, P. Fasting and Postprandial Concentrations of GLP-1 in Intestinal Lymph and Portal Plasma: Evidence for Selective Release of GLP-1 in the Lymph System. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2007, 293, R2163–R2169. [Google Scholar] [CrossRef]
- Yuan, D.; Zhou, C.; Wang, T.; Lin, F.; Chen, H.; Wu, H.; Wei, R.; Xin, Z.; Liu, J.; Gao, Y.; et al. Molecular Characterization and Tissue Expression of Peptide YY in Schizothorax Prenanti: Effects of Periprandial Changes and Fasting on Expression in the Hypothalamus. Regul. Pept. 2014, 190–191, 32–38. [Google Scholar] [CrossRef]
- Zhou, C.; Lei, L.; Yuan, D.; Deng, X.; Ye, H.; Luo, H.; Fang, J.; Yang, M.; Li, Y.; Zhang, C.; et al. Structural and Functional Characterization of Peptide YY on Feeding in Schizothorax Davidi. J. Exp. Zool. Part Ecol. Integr. Physiol. 2018, 329, 55–61. [Google Scholar] [CrossRef]
- Campos, I.; Matos, E.; Maia, M.R.G.; Marques, A.; Valente, L.M.P. Partial and Total Replacement of Fish Oil by Poultry Fat in Diets for European Seabass (Dicentrarchus labrax) Juveniles: Effects on Nutrient Utilization, Growth Performance, Tissue Composition and Lipid Metabolism. Aquaculture 2019, 502, 107–120. [Google Scholar] [CrossRef]
- Lee, H.-M.; Wang, G.; Englander, E.W.; Kojima, M.; Greeley, G.H., Jr. Ghrelin, A New Gastrointestinal Endocrine Peptide That Stimulates Insulin Secretion: Enteric Distribution, Ontogeny, Influence of Endocrine, and Dietary Manipulations. Endocrinology 2002, 143, 185–190. [Google Scholar] [CrossRef]
- Bonacic, K.; Martínez, A.; Gisbert, E.; Estévez, A.; Morais, S. Effect of Alternative Oil Sources at Different Dietary Inclusion Levels on Food Intake and Appetite Regulation via Enteroendocrine and Central Factors in Juvenile Solea Senegalensis (Kaup, 1858). Aquaculture 2017, 470, 169–181. [Google Scholar] [CrossRef]
- Charrier, J.A.; Martin, R.J.; McCutcheon, K.L.; Raggio, A.M.; Goldsmith, F.; Goita, M.; Senevirathne, R.N.; Brown, I.L.; Pelkman, C.; Zhou, J.; et al. High Fat Diet Partially Attenuates Fermentation Responses in Rats Fed Resistant Starch from High-Amylose Maize. Obesity 2013, 21, 2350–2355. [Google Scholar] [CrossRef]
- Essah, P.A.; Levy, J.R.; Sistrun, S.N.; Kelly, S.M.; Nestler, J.E. Effect of Macronutrient Composition on Postprandial Peptide YY Levels. J. Clin. Endocrinol. Metab. 2007, 92, 4052–4055. [Google Scholar] [CrossRef]
- Yu, J.; Wen, X.; You, C.; Wang, S.; Chen, C.; Tocher, D.R.; Li, Y. Comparison of the Growth Performance and Long-Chain Polyunsaturated Fatty Acids (LC-PUFA) Biosynthetic Ability of Red Tilapia (Oreochromis Mossambicus♀ × O. Niloticus♂) Fed Fish Oil or Vegetable Oil Diet at Different Salinities. Aquaculture 2021, 542, 736899. [Google Scholar] [CrossRef]
- Nickles, K.R.; Hamer, L.; Coleman, D.N.; Relling, A.E. Supplementation with Eicosapentaenoic and Docosahexaenoic Acids in Late Gestation in Ewes Changes Adipose Tissue Gene Expression in the Ewe and Growth and Plasma Concentration of Ghrelin in the Offspring1. J. Anim. Sci. 2019, 97, 2631–2643. [Google Scholar] [CrossRef]
- Xie, P.; Wan, X.P.; Bu, Z.; Zou, X.T. Molecular Cloning, Characterization, and Expression Analysis of Ghrelin and Cholecystokinin in the Pigeon (Columba Livia). Poult. Sci. 2016, 95, 2655–2666. [Google Scholar] [CrossRef]
- Bertucci, J.I.; Blanco, A.M.; Canosa, L.F.; Unniappan, S. Glucose, Amino Acids and Fatty Acids Directly Regulate Ghrelin and NUCB2/Nesfatin-1 in the Intestine and Hepatopancreas of Goldfish (Carassius Auratus) in Vitro. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2017, 206, 24–35. [Google Scholar] [CrossRef]
- Bala, V.; Rajagopal, S.; Kumar, D.; Nalli, A.; Mahavadi, S.; Sanyal, A.; Grider, J.; Murthy, K. Release of GLP-1 and PYY in Response to the Activation of G Protein-Coupled Bile Acid Receptor TGR5 Is Mediated by Epac/PLC-ε Pathway and Modulated by Endogenous H2S. Front. Physiol. 2014, 5, 420. [Google Scholar] [CrossRef]
- Hand, K.V.; Bruen, C.M.; O’Halloran, F.; Panwar, H.; Calderwood, D.; Giblin, L.; Green, B.D. Examining Acute and Chronic Effects of Short- and Long-Chain Fatty Acids on Peptide YY (PYY) Gene Expression, Cellular Storage and Secretion in STC-1 Cells. Eur. J. Nutr. 2013, 52, 1303–1313. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequence (5′-3′) | Applications | Reference |
|---|---|---|---|
| ghrelin-F1 | ATGTTGTTGAAGAGAAATACATTTTG | Cloning of ORF | Designed by author |
| ghrelin-R1 | TCAAAGCTGTGATGGTTTCTC | Cloning of ORF | Designed by author |
| glp-1-F2 | ATGGCATCTTTCCTTTCTGG | Cloning of ORF | Designed by author |
| glp-1-R2 | TCATTGGTCCGCTTGC | Cloning of ORF | Designed by author |
| pyya-F3 | ATGGCCGTGATGCTGAA | Cloning of ORF | Designed by author |
| pyya-R3 | TCACCACATGTAGGAGTCTTCATAT | Cloning of ORF | Designed by author |
| pyyb-F4 | ATGGAGCTTCTATTTCAGATTTTTT | Cloning of ORF | Designed by author |
| pyyb-R4 | TCACCACATCGGCAGCT | Cloning of ORF | Designed by author |
| pyya-F5 | CACTGCCCTGAGACACTAC | qPCR | Designed by author |
| pyya-R5 | TCTCTGTTGCTGTCGCCAC | qPCR | Designed by author |
| pyyb-F6 | AGCACAGTGATGTCGCTGAG | qPCR | Designed by author |
| pyyb-R6 | GCTTGGCTGGGTATGCATTC | qPCR | Designed by author |
| ghrelin-F7 | TTGCTGGTGTTGCTGTTGTG | qPCR | Designed by author |
| ghrelin-R7 | GCACTTATTGTGATGTGGTC | qPCR | Designed by author |
| glp-1-F8 | ACTTCGTCCACAGCAGCTG | qPCR | Designed by author |
| glp-1-R8 | ACTTCGTCCACAGCAGCTG | qPCR | Designed by author |
| β-actin-F9 | ACCCTGAAGTACCCCATCGA | qPCR | [43] |
| β-actin-R9 | GGCCACTCTCAGCTCATTGT | qPCR | [43] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, L.; Shi, P.; Zhang, L.; Wang, Y.; Xu, J.; Yan, X.; Liao, K. Molecular Characterization and Expression Response of Ghrelin, GLP-1 and PYY to Fasting, Dietary Lipid, and Fatty Acids in Silver Pomfret (Pampus argenteus). Fishes 2023, 8, 170. https://doi.org/10.3390/fishes8030170
Zhang L, Shi P, Zhang L, Wang Y, Xu J, Yan X, Liao K. Molecular Characterization and Expression Response of Ghrelin, GLP-1 and PYY to Fasting, Dietary Lipid, and Fatty Acids in Silver Pomfret (Pampus argenteus). Fishes. 2023; 8(3):170. https://doi.org/10.3390/fishes8030170
Chicago/Turabian StyleZhang, Lei, Peng Shi, Lin Zhang, Yajun Wang, Jilin Xu, Xiaojun Yan, and Kai Liao. 2023. "Molecular Characterization and Expression Response of Ghrelin, GLP-1 and PYY to Fasting, Dietary Lipid, and Fatty Acids in Silver Pomfret (Pampus argenteus)" Fishes 8, no. 3: 170. https://doi.org/10.3390/fishes8030170