
Comparison of Immune Response of Litopenaeus vannamei Shrimp Naturally Infected with Vibrio Species, and after Being Fed with Florfenicol

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
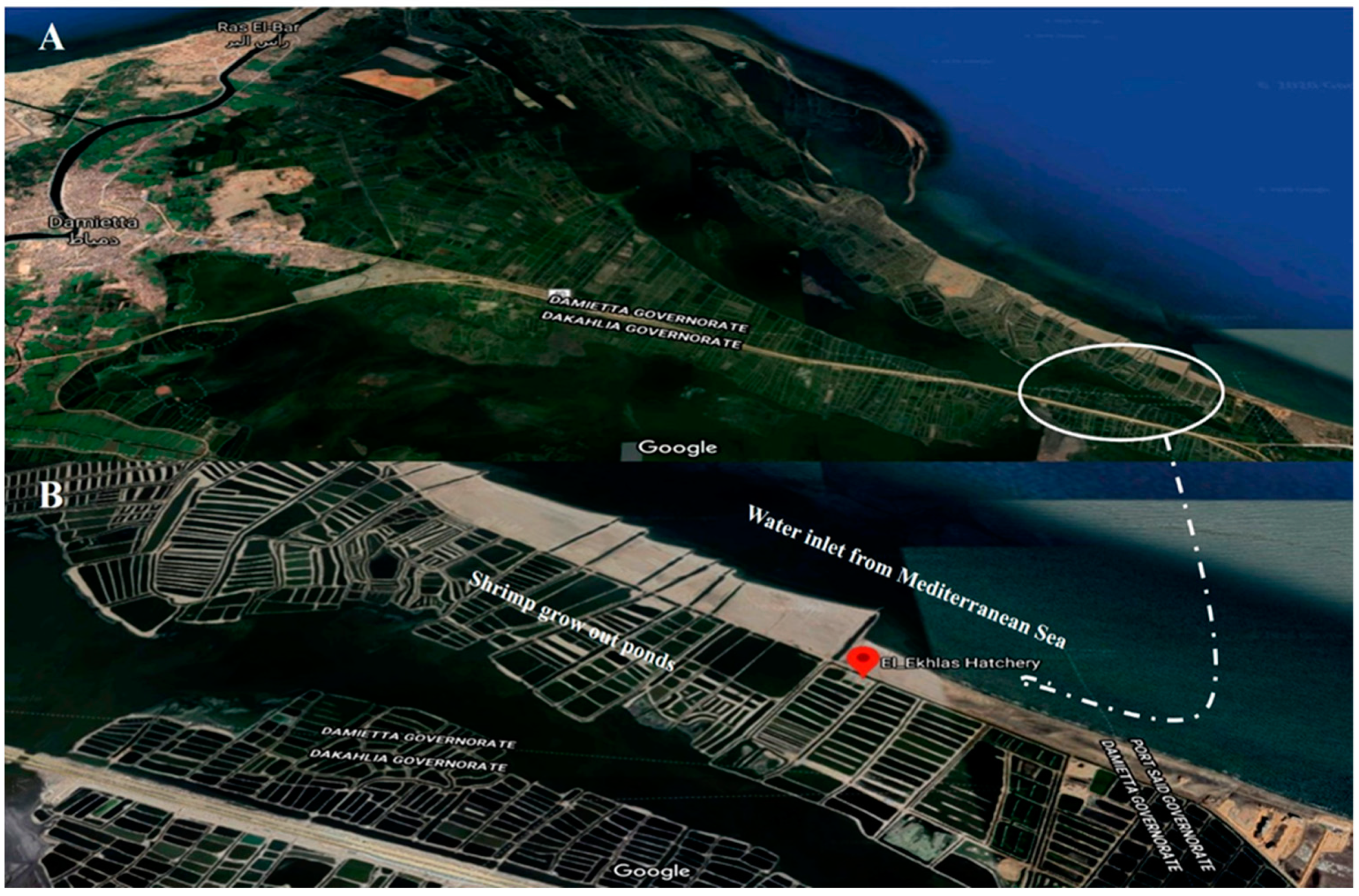
2.1. Collection of Shrimp Samples from Study Area
2.2. Isolation and Identification of Vibrio spp. Using 16S Ribosomal RNA Sequence Analysis
2.3. Susceptibility of Pathogenic Vibrio Strains to Antibiotics
2.4. Experimental Control of Vibrio Infection by Antibiotic Application
2.5. Total Hemocyte Count
2.6. Enzyme Activity Assays and Antioxidant Index
2.6.1. Hepatopancreatic Enzymes
2.6.2. Alkaline Phosphatase and Acid Phosphatase Enzymes and Antioxidants
2.7. Non-Specific Immune Response Assays
2.7.1. Phagocytosis Percent Assay
2.7.2. Respiratory Burst Assay
2.7.3. Bactericidal Activity
2.8. Total RNA Extraction, cDNA Synthesis, and Real-Time Quantitative PCR (qRT-PCR) Assay
2.9. Data Analysis
3. Results
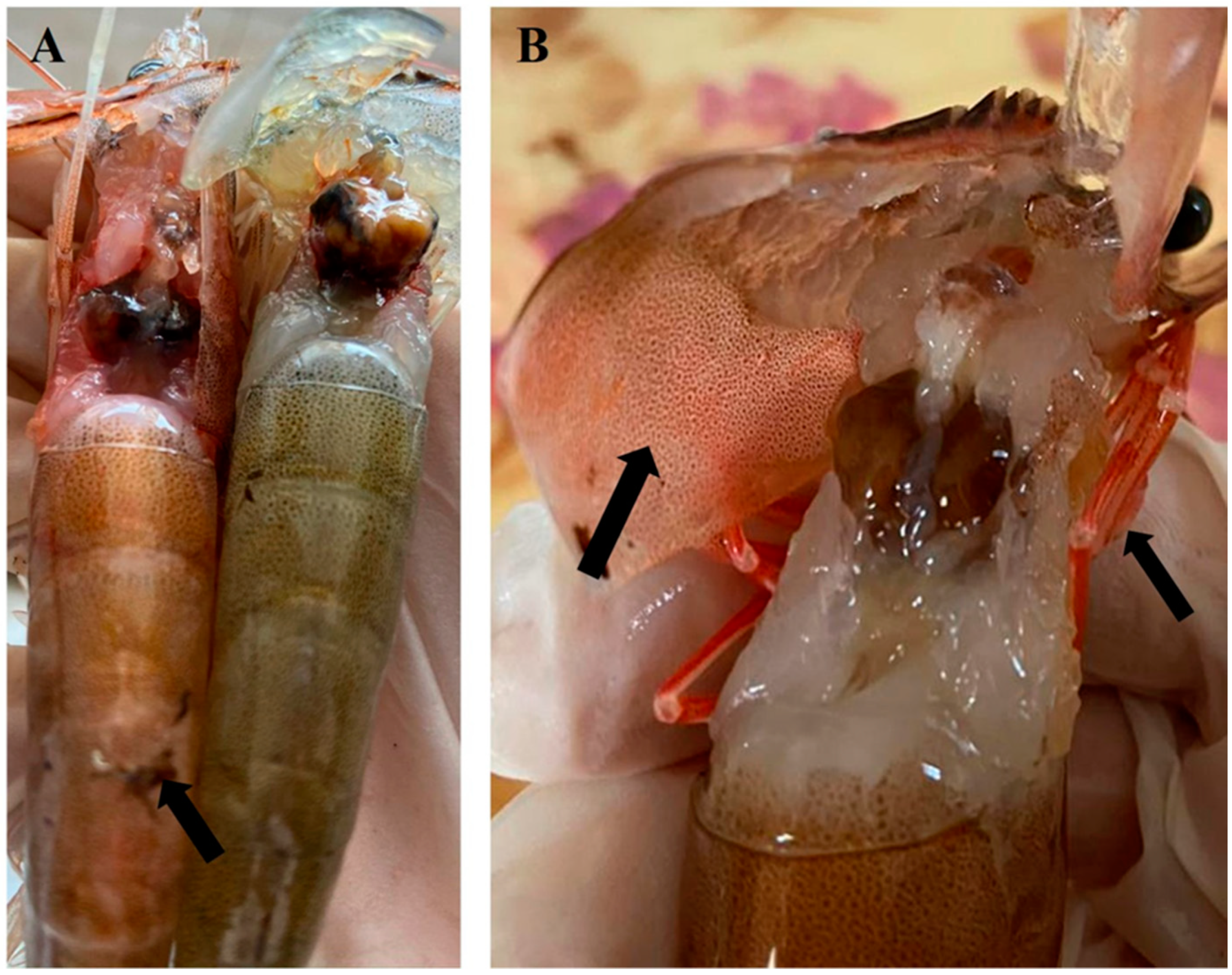
3.1. Clinical Signs of Diseased Shrimp
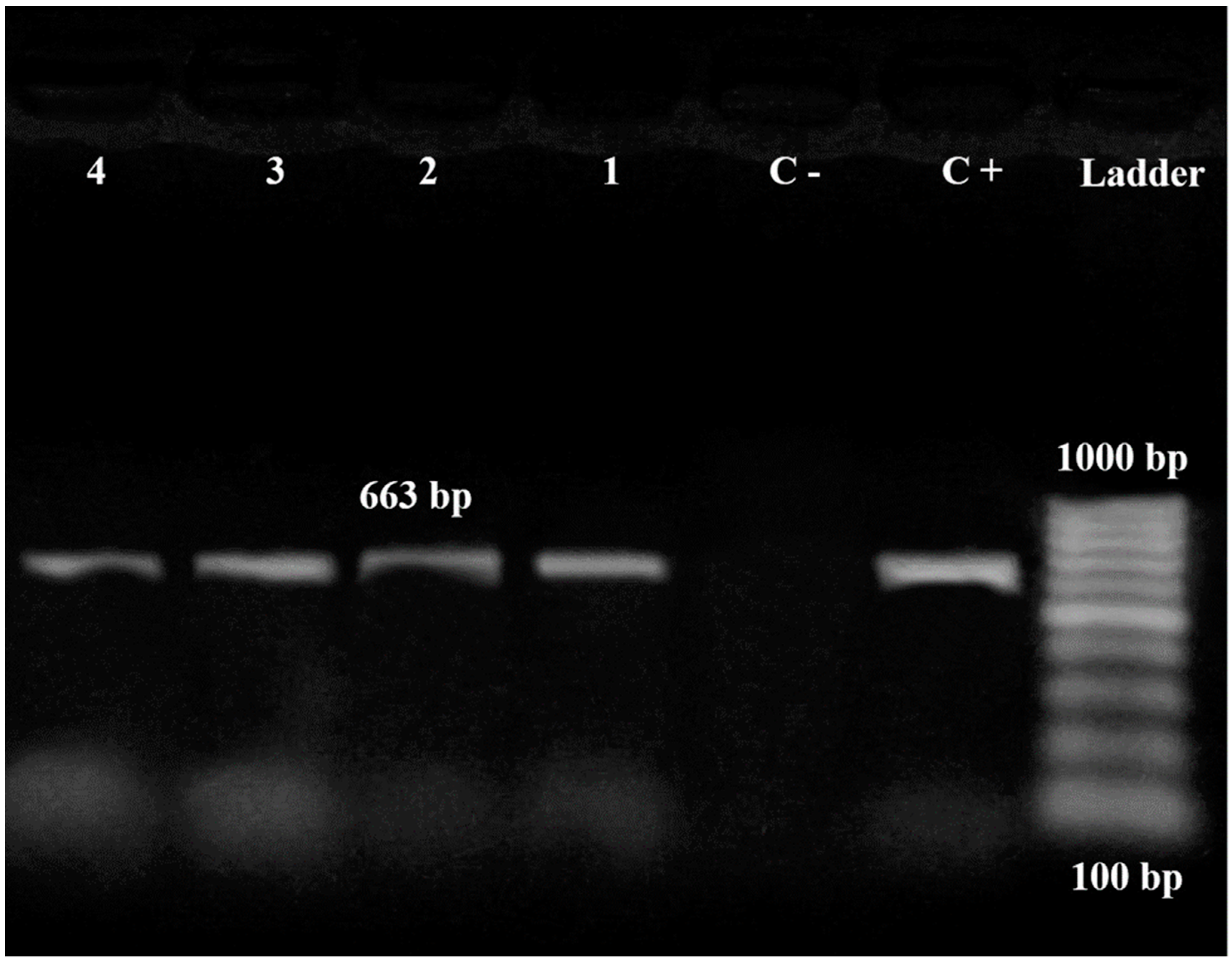
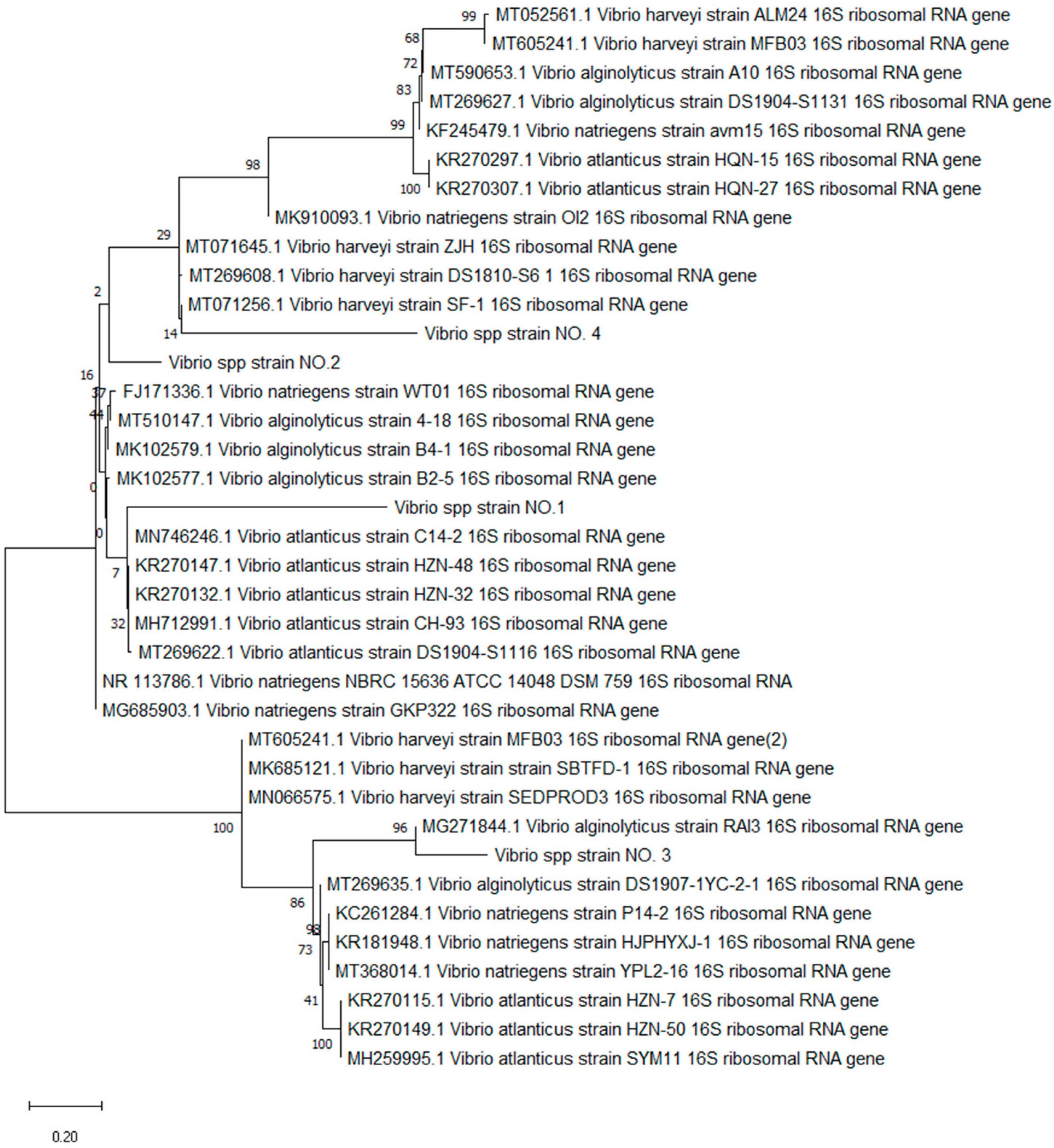
3.2. Characterization of Bacterial Vibrio Strains
3.3. Susceptibility of Pathogenic Vibrio Strains to Antibiotics
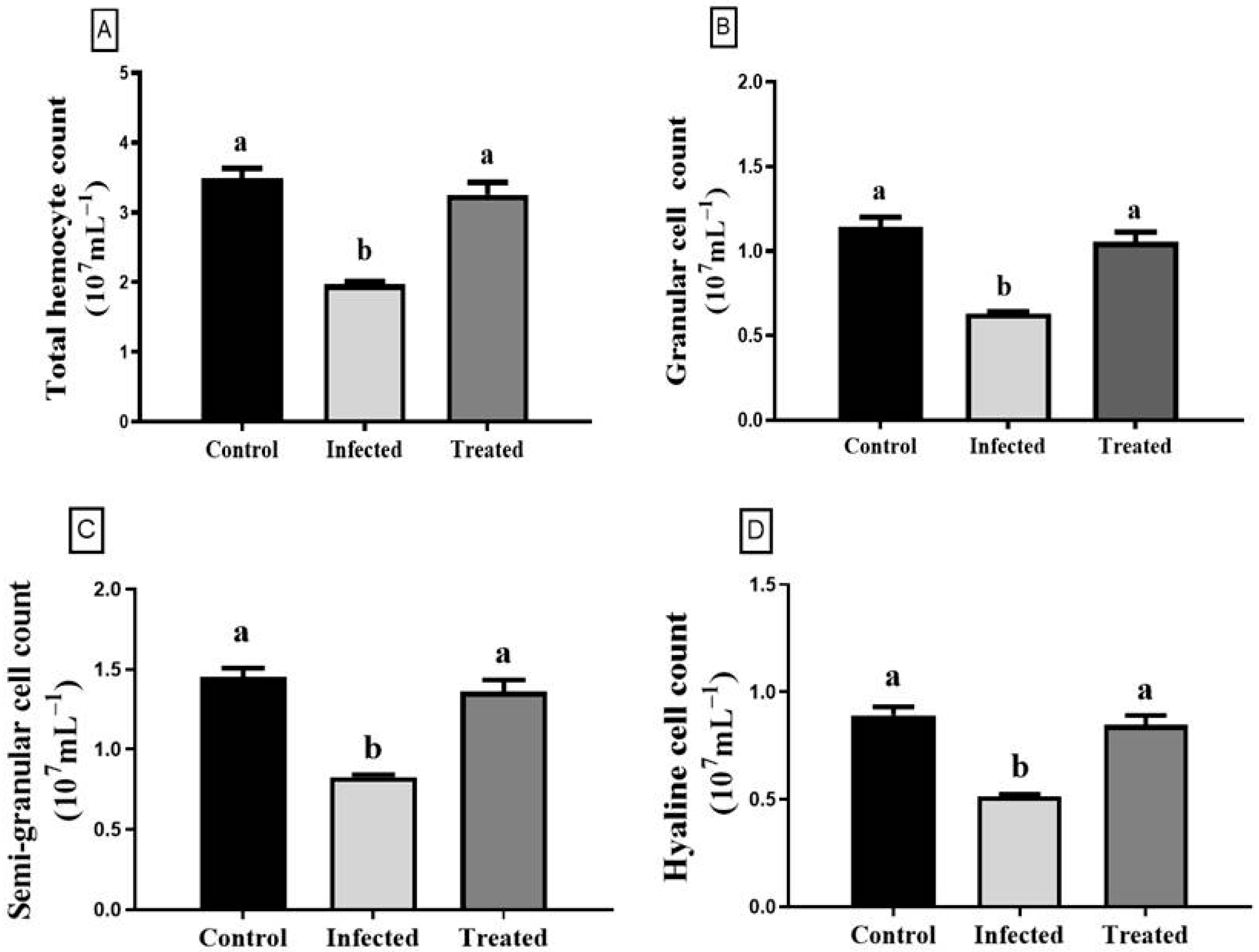
3.4. Total Hemocyte Count
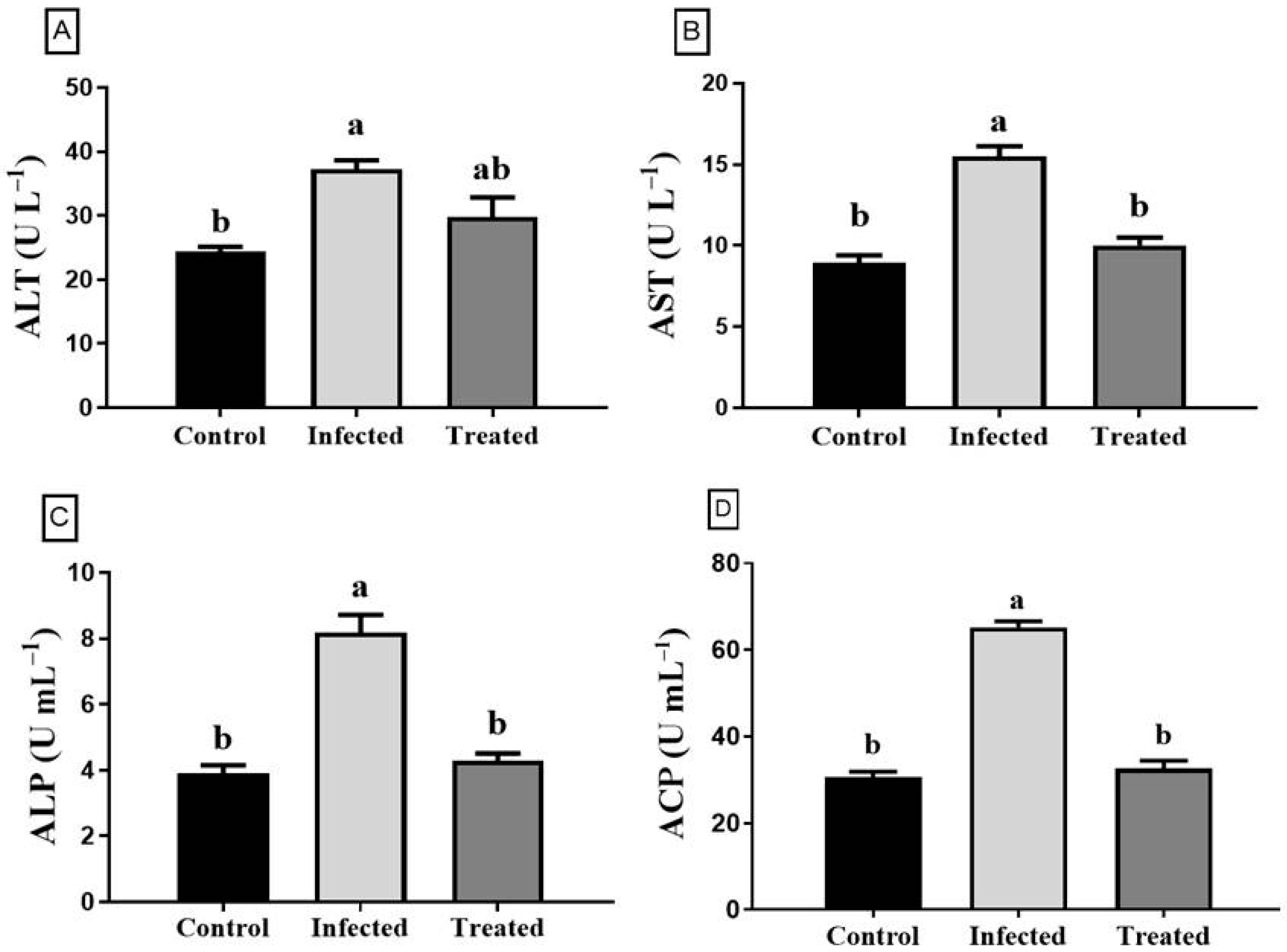
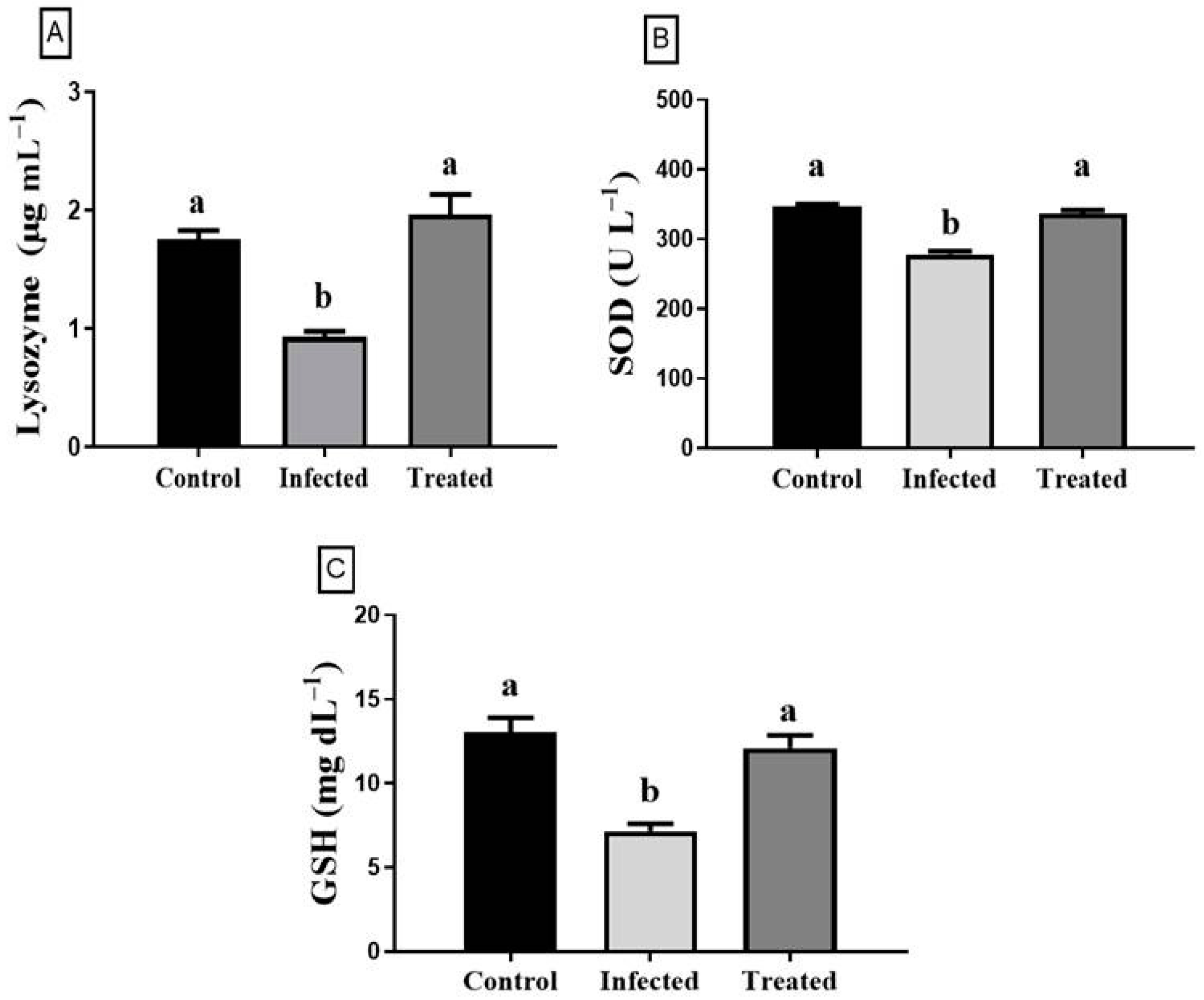
3.5. Enzyme Activity Assays and Antioxidant Index
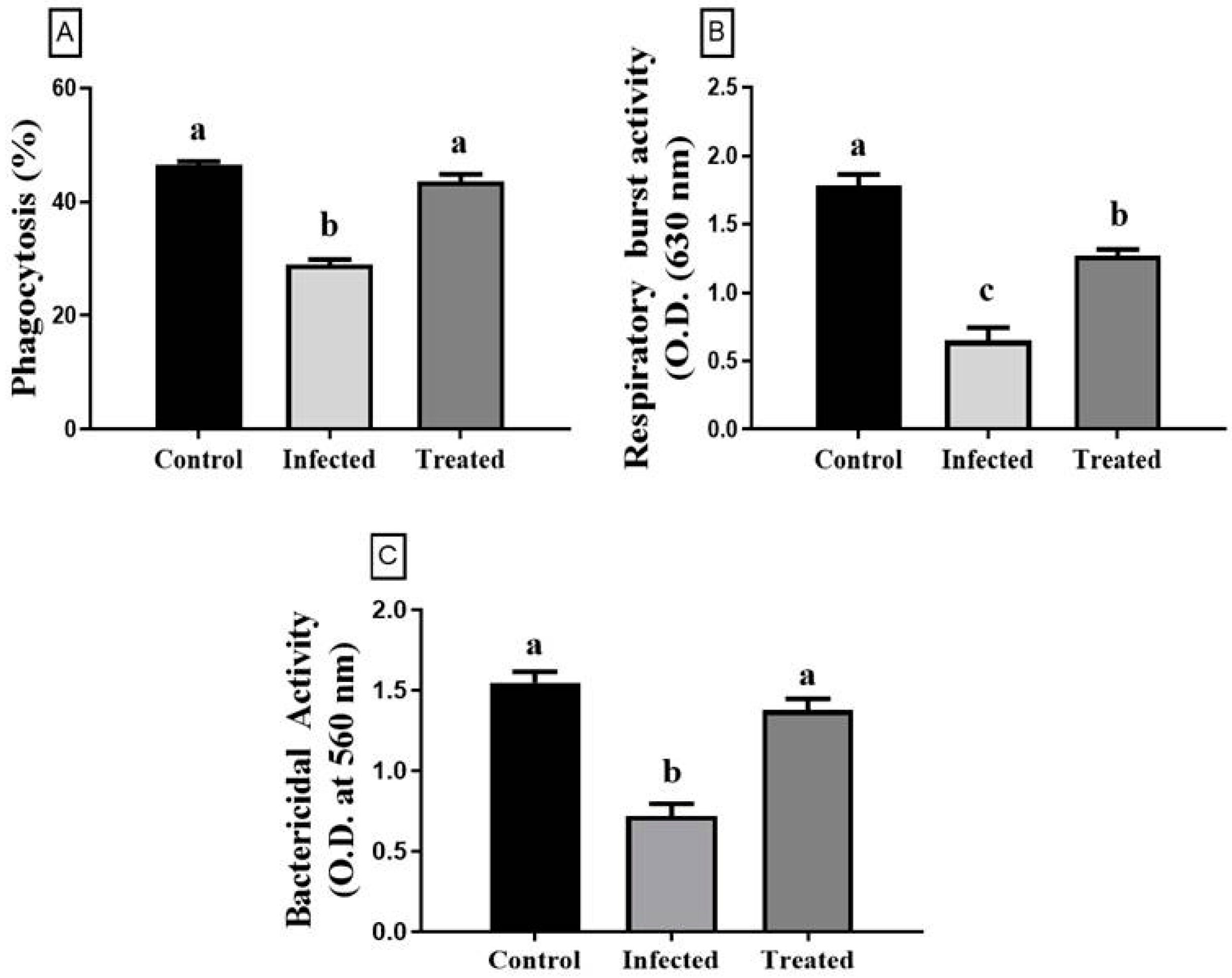
3.6. Immune Parameters
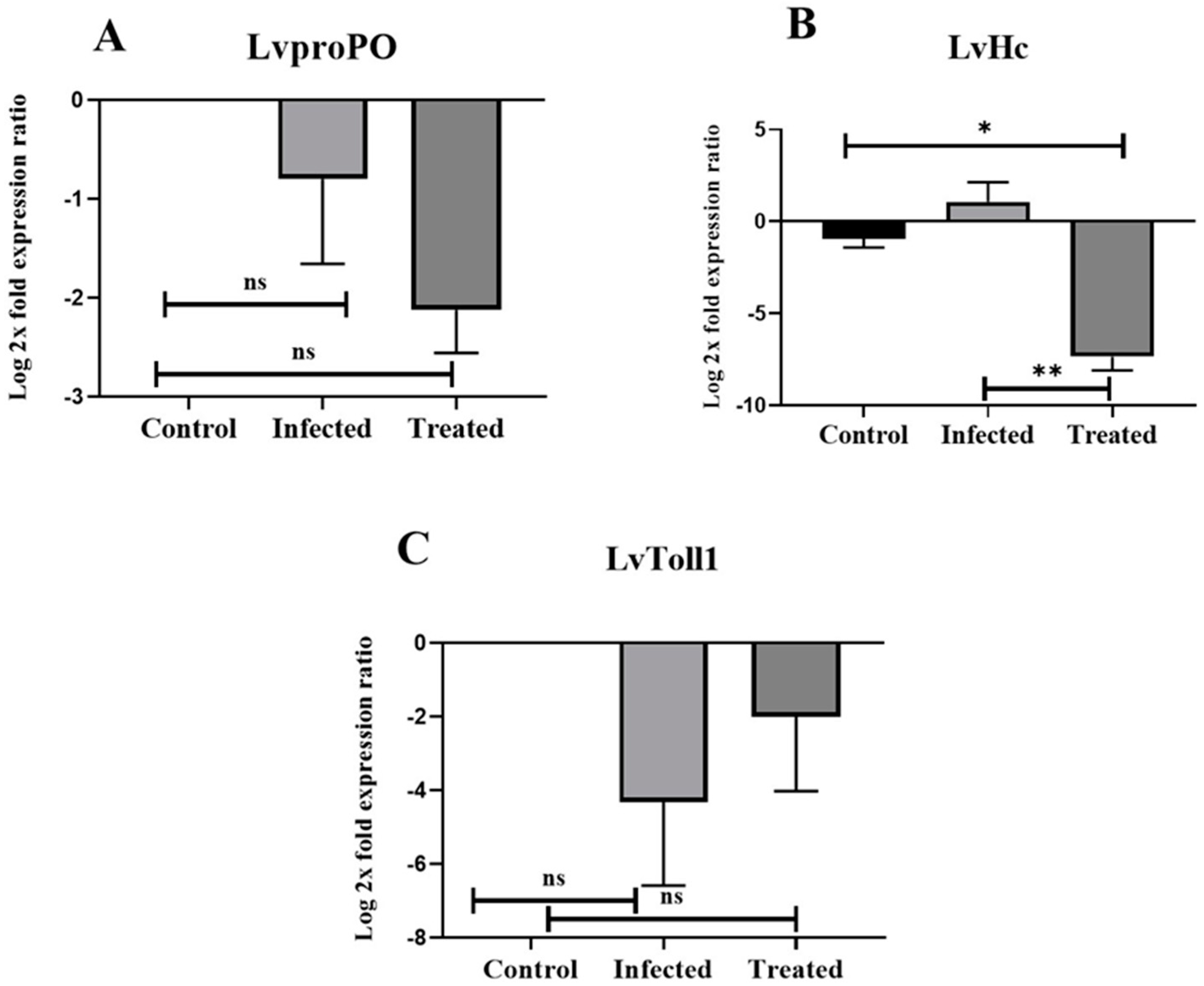
3.7. Expression Profiles of LvHc, LvToll1, and LvproPO
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Okamura, Y.; Inada, M.; Elshopakey, G.E.; Itami, T. Characterization of xanthine dehydrogenase and aldehyde oxidase of Marsupenaeus japonicus and their response to microbial pathogen. Mol. Biol. Rep. 2018, 45, 419–432. [Google Scholar] [CrossRef] [PubMed]
- Thirugnanasambandam, R.; Inbakandan, D.; Kumar, C.; Subashni, B.; Vasantharaja, R.; Abraham, L.S.; Ayyadurai, N.; Murthy, P.S.; Kirubagaran, R.; Khan, S.A. Genomic insights of Vibrio harveyi RT-6 strain, from infected “Whiteleg shrimp”(Litopenaeus vannamei) using Illumina platform. Mol. Phylogenetics Evol. 2019, 130, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Adeleke, B.; Robertson-Andersson, D.; Moodley, G.; Taylor, S. Aquaculture in Africa: A Comparative Review of Egypt, Nigeria, and Uganda Vis-À-Vis South Africa. Rev. Fish. Sci. Aquac. 2020, 29, 1–31. [Google Scholar] [CrossRef]
- Megahed, M.E. A comparison of the severity of white spot disease in cultured shrimp (Fenneropenaeus indicus) at a farm level in Egypt. I-Molecular, histopathological and field observations. Egypt. J. Aquat. Biol. Fish. 2019, 23, 613–637. [Google Scholar] [CrossRef] [Green Version]
- Feng, B.; Liu, H.; Wang, M.; Sun, X.; Pan, Y.; Zhao, Y. Diversity analysis of acute hepatopancreatic necrosis disease-positive Vibrio parahaemolyticus strains. Aquac. Fish. 2017, 2, 278–285. [Google Scholar] [CrossRef]
- Culot, A.; Grosset, N.; Bruey, Q.; Auzou, M.; Giard, J.-C.; Favard, B.; Wakatsuki, A.; Baron, S.; Frouel, S.; Techer, C.; et al. Isolation of Harveyi clade Vibrio spp. collected in aquaculture farms: How can the identification issue be addressed? J. Microbiol. Methods 2021, 180, 106106. [Google Scholar] [CrossRef] [PubMed]
- Dubert, J.; Nelson, D.R.; Spinard, E.J.; Kessner, L.; Gomez-Chiarri, M.; da Costa, F.; Prado, S.; Barja, J.L. Following the infection process of vibriosis in Manila clam (Ruditapes philippinarum) larvae through GFP-tagged pathogenic Vibrio species. J. Invertebr. Pathol. 2016, 133, 27–33. [Google Scholar] [CrossRef] [Green Version]
- Chatterjee, S.; Haldar, S. Vibrio related diseases in aquaculture and development of rapid and accurate identification methods. J. Mar. Sci. Res. Dev. S 2012, 1, 1–7. [Google Scholar]
- Mastan, S.; Begum, S.A. Vibriosis in farm reared white shrimp, Litopenaeus vannamei in Andhra Pradesh-natural occurrence and artificial challenge. Int. J. Appl. Sci. Biotechnol. 2016, 4, 217–222. [Google Scholar] [CrossRef] [Green Version]
- Baker-Austin, C.; Oliver, J.D.; Alam, M.; Ali, A.; Waldor, M.K.; Qadri, F.; Martinez-Urtaza, J. Vibrio spp. infections. Nat. Rev. Dis. Prim. 2018, 4, 1–19. [Google Scholar] [CrossRef]
- Kumaran, T.; Citarasu, T. Isolation and characterization of Vibrio species from shrimp and Artemia culture and evaluation of the potential virulence factor. Intel. Prop. Rights 2016, 4, 2. [Google Scholar]
- Zhang, X.-H.; He, X.; Austin, B. Vibrio harveyi: A serious pathogen of fish and invertebrates in mariculture. Mar. Life Sci. Technol. 2020, 2, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darshanee Ruwandeepika, H.A.; Sanjeewa Prasad Jayaweera, T.; Paban Bhowmick, P.; Karunasagar, I.; Bossier, P.; Defoirdt, T. Pathogenesis, virulence factors and virulence regulation of vibrios belonging to the Harveyi clade. Rev. Aquac. 2012, 4, 59–74. [Google Scholar] [CrossRef]
- Hasan, M.A.R.; Siddique, M.A.; Hasan, M.; Hossain, M.A.; Rahman, M.S. 16S rRNA gene sequence based identification of Vibrio spp. in shrimp and tilapia hatcheries of Bangladesh. Dhaka Univ. J. Biol. Sci. 2017, 26, 45–58. [Google Scholar] [CrossRef] [Green Version]
- Janda, J.M.; Abbott, S.L. 16S rRNA gene sequencing for bacterial identification in the diagnostic laboratory: Pluses, perils, and pitfalls. J. Clin. Microbiol. 2007, 45, 2761–2764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karim, M.; Uddin, M.; Rahman, M. Microbiological study of costal shrimp aquaculture production system of Bangladesh. J. Biol. Life Sci. 2018, 9, 17–30. [Google Scholar]
- Thornber, K.; Verner-Jeffreys, D.; Hinchliffe, S.; Rahman, M.M.; Bass, D.; Tyler, C.R. Evaluating antimicrobial resistance in the global shrimp industry. Rev. Aquac. 2020, 12, 966–986. [Google Scholar] [CrossRef] [Green Version]
- Cabello, F.C. Heavy use of prophylactic antibiotics in aquaculture: A growing problem for human and animal health and for the environment. Environ. Microbiol. 2006, 8, 1137–1144. [Google Scholar] [CrossRef]
- De Schryver, P.; Defoirdt, T.; Sorgeloos, P. Early mortality syndrome outbreaks: A microbial management issue in shrimp farming? PLoS Pathog. 2014, 10, e1003919. [Google Scholar] [CrossRef]
- Yanong, R.P.; Curtis, E.W.; Simmons, R.; Bhattaram, V.A.; Gopalakrishnan, M.; Ketabi, N.; Nagaraja, N.V.; Derendorf, H. Pharmacokinetic studies of florfenicol in koi carp and threespot gourami Trichogaster trichopterus after oral and intramuscular treatment. J. Aquat. Anim. Health 2005, 17, 129–137. [Google Scholar] [CrossRef]
- Brausch, J.M.; Connors, K.A.; Brooks, B.W.; Rand, G.M. Human pharmaceuticals in the aquatic environment: A review of recent toxicological studies and considerations for toxicity testing. Rev. Environ. Contam. Toxicol. 2012, 218, 1–99. [Google Scholar] [PubMed]
- Bowker, J.D.; Ostland, V.E.; Carty, D.; Bowman, M.P. Effectiveness of Aquaflor (50% florfenicol) to control mortality associated with Streptococcus iniae in freshwater-reared subadult sunshine bass. J. Aquat. Anim. Health 2010, 22, 254–265. [Google Scholar] [CrossRef]
- Carty, D.; Bowker, J.; Bowman, M.; Erdahl, D. Calculate Amount of Aquaflor (Florfenicol, 501‰) to Add to Fish Feed; AADAP Drug Research Information Bulletin of the US Fish and Wildlife Services; U.S. Fish & Wildlife Service: Washington, DC, USA, 2007.
- de Oliveira, T.F.; Queiroz, G.A.; Teixeira, J.P.; Figueiredo, H.C.P.; Leal, C.A.G. Recurrent Streptoccoccus agalactiae infection in Nile tilapia (Oreochromis niloticus) treated with florfenicol. Aquaculture 2018, 493, 51–60. [Google Scholar] [CrossRef]
- Samuelsen, O.B.; Bergh, Ø.; Ervik, A. Pharmacokinetics of florfenicol in cod Gadus morhua and in vitro antibacterial activity against Vibrio anguillarum. Dis. Aquat. Org. 2003, 56, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Sotomayor, M.A.; Reyes, J.K.; Restrepo, L.; Domínguez-Borbor, C.; Maldonado, M.; Bayot, B. Efficacy assessment of commercially available natural products and antibiotics, commonly used for mitigation of pathogenic Vibrio outbreaks in Ecuadorian Penaeus (Litopenaeus) vannamei hatcheries. PloS ONE 2019, 14, e0210478. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Córdova, L.R.; Gollas-Galván, T.; Garibay-Valdez, E.; Valenzuela-Gutiérrez, R.; Porchas, M.M.; Porchas-Cornejo, M.A.; Sánchez-Paz, A.; Mendoza-Cano, F. Physiological and immune response of Litopenaeus vannamei undergoing the acute phase of the necrotizing hepatopancreatitis disease and after being treated with oxytetracycline and FF. Lat. Am. J. Aquat. Res. 2016, 44, 535–545. [Google Scholar] [CrossRef]
- Soto-Rodríguez, S.; Armenta, M.; Gomez-Gil, B. Effects of enrofloxacin and florfenicol on survival and bacterial population in an experimental infection with luminescent Vibrio campbellii in shrimp larvae of Litopenaeus vannamei. Aquaculture 2006, 255, 48–54. [Google Scholar] [CrossRef]
- Hauton, C. The scope of the crustacean immune system for disease control. J. Invertebr. Pathol. 2012, 110, 251–260. [Google Scholar] [CrossRef] [Green Version]
- Okamura, Y.; Mekata, T.; Elshopakey, G.E.; Itami, T. Molecular characterization and gene expression analysis of hypoxia-inducible factor and its inhibitory factors in kuruma shrimp Marsupenaeus japonicus. Fish Shellfish. Immunol. 2018, 79, 168–174. [Google Scholar] [CrossRef]
- Elbahnaswy, S.; Koiwai, K.; Zaki, V.H.; Shaheen, A.A.; Kondo, H.; Hirono, I. A novel viral responsive protein (MjVRP) from Marsupenaeus japonicus haemocytes is involved in white spot syndrome virus infection. Fish Shellfish. Immunol. 2017, 70, 638–647. [Google Scholar] [CrossRef]
- Lin, X.; Söderhäll, I. Crustacean hematopoiesis and the astakine cytokines. Blood J. Am. Soc. Hematol. 2011, 117, 6417–6424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tassanakajon, A.; Rimphanitchayakit, V.; Visetnan, S.; Amparyup, P.; Somboonwiwat, K.; Charoensapsri, W.; Tang, S. Shrimp humoral responses against pathogens: Antimicrobial peptides and melanization. Dev. Comp. Immunol. 2018, 80, 81–93. [Google Scholar] [CrossRef] [PubMed]
- Jiravanichpaisal, P.; Lee, B.L.; Söderhäll, K. Cell-mediated immunity in arthropods: Hematopoiesis, coagulation, melanization and opsonization. Immunobiology 2006, 211, 213–236. [Google Scholar] [CrossRef]
- Bergey, D.H. Bergey’s Manual of Determinative Bacteriology; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 1994. [Google Scholar]
- Buller, N.B. Bacteria from Fish and Other Aquatic Animals: A Practical Identification Manual; CABI Publishing: Wallingford, UK, 2004. [Google Scholar]
- Costinar, L.; Herman, V.; Pascu, C.; Marcu, A.; Marcu, A.; Faur, B. Isolation and characterization of Vibrio alginolyticus and Pasteurella spp. from Siberian Sturgeon (Acipenser Baerii). Lucr. Stiinłifice Med. Vet. 2010, 43, 125–127. [Google Scholar]
- Tarr, C.L.; Patel, J.S.; Puhr, N.D.; Sowers, E.G.; Bopp, C.A.; Strockbine, N.A. Identification of Vibrio isolates by a multiplex PCR assay and rpoB sequence determination. J. Clin. Microbiol. 2007, 45, 134–140. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Zidour, M.; Chevalier, M.; Belguesmia, Y.; Cudennec, B.; Grard, T.; Drider, D.; Souissi, S.; Flahaut, C. Isolation and characterization of bacteria colonizing Acartia tonsa copepod eggs and displaying antagonist effects against Vibrio anguillarum, Vibrio alginolyticus and other pathogenic strains. Front. Microbiol. 2017, 8, 1919. [Google Scholar] [CrossRef]
- Balouiri, M.; Sadiki, M.; Ibnsouda, S.K. Methods for in vitro evaluating antimicrobial activity: A review. J. Pharm. Anal. 2016, 6, 71–79. [Google Scholar] [CrossRef] [Green Version]
- El-Son, M.A.; Elbahnaswy, S.; Ibrahim, I. Molecular and histopathological characterization of Photobacterium damselae in naturally and experimentally infected Nile tilapia (Oreochromis niloticus). J. Fish Dis. 2020, 43, 1505–1517. [Google Scholar] [CrossRef]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing; Twenty-Fifth Informational Supplement M100-S25; Clinical and Laboratory Standards Institute (CLSI): Wayne, PA, USA, 2015. [Google Scholar]
- Liu, C.-H.; Yeh, S.-T.; Cheng, S.-Y.; Chen, J.-C. The immune response of the white shrimp Litopenaeus vannamei and its susceptibility to Vibrio infection in relation with the moult cycle. Fish Shellfish. Immunol. 2004, 16, 151–161. [Google Scholar] [CrossRef]
- Elshopakey, G.E.; Risha, E.F.; Abdalla, O.A.; Okamura, Y.; Hanh, V.D.; Ibuki, M.; Sudhakaran, R.; Itami, T. Enhancement of immune response and resistance against Vibrio parahaemolyticus in kuruma shrimp (Marsupenaeus japonicus) by dietary supplementation of β-1, 4-mannobiose. Fish Shellfish. Immunol. 2018, 74, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Vaseeharan, B.; Ramasamy, P.; Wesley, S.G.; Chen, J.C. Influence of acute salinity changes on biochemical, hematological and immune characteristics of Fenneropenaeus indicus during white spot syndrome virus challenge. Microbiol. Immunol. 2013, 57, 463–469. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.-C.; Tayag, C.M.; Huang, C.-L.; Tsui, W.-C.; Chen, J.-C. White shrimp Litopenaeus vannamei that had received the hot-water extract of Spirulina platensis showed earlier recovery in immunity and up-regulation of gene expressions after pH stress. Fish Shellfish. Immunol. 2010, 29, 1092–1098. [Google Scholar] [CrossRef] [PubMed]
- Biagini, G.; Sala, D.; Zini, I. Diethyldithiocarbamate, a superoxide dismutase inhibitor, counteracts the maturation of ischemic-like lesions caused by endothelin-1 intrastriatal injection. Neurosci. Lett. 1995, 190, 212–216. [Google Scholar] [CrossRef]
- Jollow, D.; Mitchell, J.; Zampaglione, N.a.; Gillette, J. Bromobenzene-induced liver necrosis. Protective role of glutathione and evidence for 3, 4-bromobenzene oxide as the hepatotoxic metabolite. Pharmacology 1974, 11, 151–169. [Google Scholar] [CrossRef]
- Elshopakey, G.E.; Risha, E.F.; Abdalla, O.A.; Okamura, Y.; Harada, S.; Kishida, S.; Matsuura, Y.; Sudhakaran, R.; Itami, T. Efficacy of dietary fermented vegetable product on immune response, up-regulation of immune-related genes and protection of kuruma shrimp (Marsupenaeus japonicus) against Vibrio parahaemolyticus. Aquaculture 2018, 497, 431–439. [Google Scholar] [CrossRef]
- Elbahnaswy, S.; Elshopakey, G.E.; Ibrahim, I.; Habotta, O.A. Potential role of dietary chitosan nanoparticles against immunosuppression, inflammation, oxidative stress, and histopathological alterations induced by pendimethalin toxicity in Nile tilapia. Fish Shellfish. Immunol. 2021, 118, 270–282. [Google Scholar] [CrossRef]
- Andrino, K.G.S.; Apines-Amar, M.J.S.; Janeo, R.L.; Corre, V.L., Jr. Effects of dietary mannan oligosaccharide (MOS) and β-glucan on growth, immune response and survival against white spot syndrome virus (WSSV) infection of juvenile tiger shrimp Penaeus monodon. Aquac. Aquar. Conserv. Legis. 2014, 7, 321–332. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Wang, Y.-C.; Chang, P.-S.; Chen, H.-Y. Tissue distribution of prophenoloxidase transcript in the Pacific white shrimp Litopenaeus vannamei. Fish Shellfish. Immunol. 2006, 20, 414–418. [Google Scholar] [CrossRef]
- Soto-Alcalá, J.; Álvarez-Ruiz, P.; Audelo-Naranjo, J.M.; Esparza-Leal, H.M.; Luis-Villaseñor, J.; Estrada-Godínez, A.; Luna-González, A.; Gámez-Jiménez, C.; Diarte-Plata, G. Transcriptional response of immune-related genes in Litopenaeus vannamei post-larvae cultured in recirculating aquaculture systems with and without biofloc. Aquacult. Int. 2019, 27, 209–225. [Google Scholar] [CrossRef]
- Aguilera-Rivera, D.; Escalante-Herrera, K.; Gaxiola, G.; Prieto-Davó, A.; Rodríguez-Fuentes, G.; Guerra-Castro, E.; Hernández-López, J.; Chávez-Sánchez, M.C.; Rodríguez-Canul, R. Immune response of the Pacific white shrimp, Litopenaeus vannamei, previously reared in biofloc and after an infection assay with Vibrio harveyi. J. World Aquac. Soc. 2019, 50, 119–136. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.-H.; Liang, J.-P.; Gu, Z.-H.; Wan, D.-H.; Weng, S.-P.; Yu, X.-Q.; He, J.-G. Molecular cloning, characterization and expression analysis of two novel Tolls (LvToll2 and LvToll3) and three putative Spätzle-like Toll ligands (LvSpz1–3) from Litopenaeus vannamei. Dev. Comp. Immunol. 2012, 36, 359–371. [Google Scholar] [CrossRef] [PubMed]
- Soto-Rodriguez, S.A.; Gomez-Gil, B.; Lozano, R. ‘Bright-red’syndrome in Pacific white shrimp Litopenaeus vannamei is caused by Vibrio harveyi. Dis. Aquat. Org. 2010, 92, 11–19. [Google Scholar] [CrossRef] [Green Version]
- Soto-Rodriguez, S.A.; Gomez-Gil, B.; Lozano, R.; del Rio-Rodríguez, R.; Diéguez, A.L.; Romalde, J.L. Virulence of Vibrio harveyi responsible for the “Bright-red” Syndrome in the Pacific white shrimp Litopenaeus vannamei. J. Invertebr. Pathol. 2012, 109, 307–317. [Google Scholar] [CrossRef]
- Xiao, J.; Liu, L.; Ke, Y.; Li, X.; Liu, Y.; Pan, Y.; Yan, S.; Wang, Y. Shrimp AHPND-causing plasmids encoding the PirAB toxins as mediated by pirAB-Tn903 are prevalent in various Vibrio species. Sci. Rep. 2017, 7, 42177. [Google Scholar] [CrossRef] [Green Version]
- Fu, S.; Song, B.; Liu, Y.; Liu, Z. Screening of probiotic strains and their protection against vibriosis in juvenile shrimp (Litopenaeus vannamei). China Environ. Sci. 2009, 29, 867–872. [Google Scholar]
- Cano-Gomez, A.; Bourne, D.G.; Hall, M.R.; Owens, L.; Høj, L. Molecular identification, typing and tracking of Vibrio harveyi in aquaculture systems: Current methods and future prospects. Aquaculture 2009, 287, 1–10. [Google Scholar] [CrossRef]
- Tendencia, E.A.; de la Peña, L.D. Antibiotic resistance of bacteria from shrimp ponds. Aquaculture 2001, 195, 193–204. [Google Scholar] [CrossRef]
- Chiou, J.; Li, R.; Chen, S. CARB-17 family of β-lactamases mediates intrinsic resistance to penicillins in Vibrio parahaemolyticus. Antimicrob. Agents Chemother. 2015, 59, 3593–3595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morales-Covarrubias, M.S.; Tlahuel-Vargas, L.; Martínez-Rodríguez, I.E.; Lozano-Olvera, R.; Palacios-Arriaga, J.M. Necrotising hepatobacterium (NHPB) infection in Penaeus vannamei with florfenicol and oxytetracycline: A comparative experimental study. Rev. Científica 2012, 22, 72–80. [Google Scholar]
- Tipmongkolsilp, N.; Limpanon, Y.; Patamalai, B.; Lusanandana, P.; Wongtavatchai, J. Oral medication with florfenicol for black tiger shrimps Penaeus monodon. Thai J. Vet. Med. 2006, 36, 39–47. [Google Scholar]
- Roque, A.; Molina-Aja, A.; Bolán-Mejıa, C.; Gomez-Gil, B. In vitro susceptibility to 15 antibiotics of Vibrios isolated from penaeid shrimps in Northwestern Mexico. Int. J. Antimicrob. Agents 2001, 17, 383–387. [Google Scholar] [CrossRef]
- Fang, W.; Li, G.; Zhou, S.; Li, X.; Hu, L.; Zhou, J. Pharmacokinetics and tissue distribution of thiamphenicol and florfenicol in Pacific white shrimp Litopenaeus vannamei in freshwater following oral administration. J. Aquat. Anim. Health 2013, 25, 83–89. [Google Scholar] [CrossRef]
- Ren, X.; Pan, L.; Wang, L. Effect of florfenicol on selected parameters of immune and antioxidant systems, and damage indexes of juvenile Litopenaeus vannamei following oral administration. Aquaculture 2014, 432, 106–113. [Google Scholar] [CrossRef]
- Ramalingam, K.; Shyamala, D.; Kumaran, N.S.; Karthik, R.; Vanitha, M. Manifestations Studies on Enzyme Profile of Vibrio parahaemolyticus MTCC451 Inoculated Black Tiger Prawn Penaeus monodon. J. Fish. Aquat. Sci. 2015, 10, 477. [Google Scholar] [CrossRef]
- Chang, Z.Q.; Ge, Q.Q.; Sun, M.; Wang, Q.; Lv, H.Y.; Li, J. Immune responses by dietary supplement with Astragalus polysaccharides in the Pacific white shrimp, Litopenaeus vannamei. Aquac. Nutr. 2018, 24, 702–711. [Google Scholar] [CrossRef]
- Araya, M.T.; Siah, A.; Mateo, D.R.; Markham, F.; McKenna, P.; Johnson, G.R.; Berthe, F.C. Morphological and molecular effects of Vibrio splendidus on hemocytes of softshell clams, Mya arenaria. J. Shellfish. Res. 2009, 28, 751–758. [Google Scholar] [CrossRef]
- Vieira, F.; Buglione, C.; Mourino, J.; Jatobá, A.; Martins, M.; Schleder, D.; Andreatta, E.; Barraco, M.; Vinatea, L. Effect of probiotic supplemented diet on marine shrimp survival after challenge with Vibrio harveyi. Arq. Bras. De Med. Veterinária E Zootec. 2010, 62, 631–638. [Google Scholar] [CrossRef] [Green Version]
- Van de Braak, C.; Botterblom, M.; Taverne, N.v.; Van Muiswinkel, W.; Rombout, J.; Van der Knaap, W. The roles of haemocytes and the lymphoid organ in the clearance of injected Vibrio bacteria in Penaeus monodon shrimp. Fish Shellfish. Immunol. 2002, 13, 293–309. [Google Scholar] [CrossRef]
- Liang, F.; Li, C.; Hou, T.; Wen, C.; Kong, S.; Ma, D.; Sun, C.; Li, S. Effects of chitosan–gentamicin conjugate supplement on non-specific immunity, aquaculture water, intestinal histology and microbiota of pacific white shrimp (Litopenaeus vannamei). Mar. Drugs 2020, 18, 419. [Google Scholar] [CrossRef]
- Zhai, Q.; Li, J. Effectiveness of traditional Chinese herbal medicine, San-Huang-San, in combination with enrofloxacin to treat AHPND-causing strain of Vibrio parahaemolyticus infection in Litopenaeus vannamei. Fish Shellfish. Immunol. 2019, 87, 360–370. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zhuang, Z.; Yin, P.; Chen, X.; Zhang, Y.; Tian, L.; Niu, J.; Liu, Y. Changes in growth performance, haematological parameters, hepatopancreas histopathology and antioxidant status of pacific white shrimp (Litopenaeus vannamei) fed oxidized fish oil: Regulation by dietary myo-inositol. Fish Shellfish. Immunol. 2019, 88, 53–64. [Google Scholar] [CrossRef]
- Zhai, Q.; Li, J.; Feng, Y.; Ge, Q. Evaluation of combination effects of Astragalus polysaccharides and florfenicol against acute hepatopancreatic necrosis disease-causing strain of Vibrio parahaemolyticus in Litopenaeus vannamei. Fish Shellfish. Immunol. 2019, 86, 374–383. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.-L.; Xi, Q.-Y.; Yang, L.; Li, H.-Y.; Jiang, Q.-Y.; Shu, G.; Wang, S.-B.; Gao, P.; Zhu, X.-T.; Zhang, Y.-L. The effect of dietary Panax ginseng polysaccharide extract on the immune responses in white shrimp, Litopenaeus vannamei. Fish Shellfish. Immunol. 2011, 30, 495–500. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.-L.; Li, Z.-J.; Yang, K.; Lin, H.-Z.; Guo, Z.-X. Effect of guava leaves on growth and the non-specific immune response of Penaeus monodon. Fish Shellfish. Immunol. 2014, 40, 190–196. [Google Scholar] [CrossRef]
- Sarlin, P.a.; Philip, R. Efficacy of marine yeasts and baker’s yeast as immunostimulants in Fenneropenaeus indicus: A comparative study. Aquaculture 2011, 321, 173–178. [Google Scholar] [CrossRef]
- Deng, B.; Wang, Z.; Tao, W.; Li, W.; Wang, C.; Wang, M.; Ye, S.; Du, Y.; Wu, X.; Wu, D. Effects of polysaccharides from mycelia of Cordyceps sinensis on growth performance, immunity and antioxidant indicators of the white shrimp Litopenaeus vannamei. Aquac. Nutr. 2015, 21, 173–179. [Google Scholar] [CrossRef]
- Wang, X.; Wang, L.; Zhang, H.; Ji, Q.; Song, L.; Qiu, L.; Zhou, Z.; Wang, M.; Wang, L. Immune response and energy metabolism of Chlamys farreri under Vibrio anguillarum challenge and high temperature exposure. Fish Shellfish. Immunol. 2012, 33, 1016–1026. [Google Scholar] [CrossRef]
- Pang, H.; Wang, G.; Zhou, S.; Wang, J.; Zhao, J.; Hoare, R.; Monaghan, S.J.; Wang, Z.; Sun, C. Survival and immune response of white shrimp Litopenaeus vannamei following single and concurrent infections with WSSV and Vibrio parahaemolyticus. Fish Shellfish. Immunol. 2019, 92, 712–718. [Google Scholar] [CrossRef]
- Zhao, Y.; Yuan, L.; Wan, J.; Sun, H.; Wang, Y.; Zhang, Q. Effects of a potential autochthonous probiotic Bacillus subtilis 2-1 on the growth and intestinal microbiota of juvenile sea cucumber, Apostichopus japonicus Selenka. J. Ocean. Univ. China 2018, 17, 363–370. [Google Scholar] [CrossRef]
- Kohen, R.; Nyska, A. Invited review: Oxidation of biological systems: Oxidative stress phenomena, antioxidants, redox reactions, and methods for their quantification. Toxicol. Pathol. 2002, 30, 620–650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Couto, N.; Malys, N.; Gaskell, S.J.; Barber, J. Partition and turnover of glutathione reductase from Saccharomyces cerevisiae: A proteomic approach. J. Proteome Res. 2013, 12, 2885–2894. [Google Scholar] [CrossRef] [PubMed]
- Li, C.-C.; Yeh, S.-T.; Chen, J.-C. The immune response of white shrimp Litopenaeus vannamei following Vibrio alginolyticus injection. Fish Shellfish. Immunol. 2008, 25, 853–860. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Wang, Q.; Li, J.; Li, J.T.; Liu, Q.; Liu, P. Effects on lysozyme and phosphatase activities of Portunus trituberculatus infected by Vibrio alginolyticus. Prog. Fish. Sci. 2009, 30, 78–82. [Google Scholar]
- Munoz, M.; Cedeño, R.; Rodríguez, J.; van der Knaap, W.P.; Mialhe, E.; Bachère, E. Measurement of reactive oxygen intermediate production in haemocytes of the penaeid shrimp, Penaeus vannamei. Aquaculture 2000, 191, 89–107. [Google Scholar] [CrossRef] [Green Version]
- Saptiani, G.; Sidik, A.S.; Ardhani, F.; Hardi, E.H. Response of hemocytes profile in the black tiger shrimp (Penaeus monodon) against Vibrio harveyi induced by Xylocarpus granatum leaves extract. Vet. World 2019, 13, 751. [Google Scholar] [CrossRef] [Green Version]
- Cheng, A.-C.; Lin, H.-L.; Shiu, Y.-L.; Tyan, Y.-C.; Liu, C.-H. Isolation and characterization of antimicrobial peptides derived from Bacillus subtilis E20-fermented soybean meal and its use for preventing Vibrio infection in shrimp aquaculture. Fish Shellfish. Immunol. 2017, 67, 270–279. [Google Scholar] [CrossRef]
- Tseng, D.-Y.; Ho, P.-L.; Huang, S.-Y.; Cheng, S.-C.; Shiu, Y.-L.; Chiu, C.-S.; Liu, C.-H. Enhancement of immunity and disease resistance in the white shrimp, Litopenaeus vannamei, by the probiotic, Bacillus subtilis E20. Fish Shellfish. Immunol. 2009, 26, 339–344. [Google Scholar] [CrossRef]
- Allam, B.; Paillard, C.; Auffret, M.; Ford, S.E. Effects of the pathogenic Vibrio tapetis on defence factors of susceptible and non-susceptible bivalve species: II. Cellular and biochemical changes following in vivo challenge. Fish Shellfish. Immunol. 2006, 20, 384–397. [Google Scholar] [CrossRef]
- Baucheun, P.; Somboonwiwat, K.; Tassanakajon, A. Inhibitory activity of alpha-2-macroglobulin from Pacific white shrimp Litopenaeus vannamei against Vibrio harveyi proteases. In Proceedings of the The 26th annual meeting of the Thai Society for Biotechnology and International Conference, Chiang Rai, Thailand, 26–29 November 2014; pp. 559–566. [Google Scholar]
- Tassanakajon, A.; Somboonwiwat, K.; Supungul, P.; Tang, S. Discovery of immune molecules and their crucial functions in shrimp immunity. Fish Shellfish. Immunol. 2013, 34, 954–967. [Google Scholar] [CrossRef] [PubMed]
- Ai, H.-S.; Liao, J.-X.; Huang, X.-D.; Yin, Z.-X.; Weng, S.-P.; Zhao, Z.-Y.; Li, S.-D.; Yu, X.-Q.; He, J.-G. A novel prophenoloxidase 2 exists in shrimp hemocytes. Dev. Comp. Immunol. 2009, 33, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Coates, C.J.; Talbot, J. Hemocyanin-derived phenoloxidase reaction products display anti-infective properties. Dev. Comp. Immunol. 2018, 86, 47–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aoki, T.; Wang, H.-C.; Unajak, S.; Santos, M.D.; Kondo, H.; Hirono, I. Microarray analyses of shrimp immune responses. Mar. Biotechnol. 2011, 13, 629–638. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | qPCR Primers (5′–3′) | Reference |
|---|---|---|
| β-Actin-F | CCACGAGACCACCTACAAC | [54] |
| β-Actin-R | AGCGAGGGCAGTGATTTC | |
| Lv PPO-F | CTGGGCCCGGGAACTCAAG | [55] |
| Lv PPO-R | GGTGAGCATGAAGAAGAGCTGGA | |
| Lv Hc-F | GTCTTAGTGGTTCTTGGGCTTGTC | [56] |
| Lv Hc-R | GGTCTCCGTCCTGAATGTCTCC | |
| LvToll1-F | TCGACCATCCCTTTTACACC | [57] |
| LvToll1-R | TTGCCTGGAAGGTCTGATTC |
| Antibiotic | Concentration (µg) | Vibrio harveyi | Vibrio alginolyticus |
|---|---|---|---|
| amoxicillin (AML) | 25 | R | R |
| erythromycin I | 15 | s | R |
| chloramphenicI(C) | 30 | S | S |
| doxycycline (DO) | 30 | S | S |
| florfenicol (FFC) | 30 | S | S |
| ciprofloxacin (CIP) | 5 | s | S |
| norfloxacin (NOR) | 10 | s | s |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shakweer, M.S.; Elshopakey, G.E.; Abdelwarith, A.A.; Younis, E.M.; Davies, S.J.; Elbahnaswy, S. Comparison of Immune Response of Litopenaeus vannamei Shrimp Naturally Infected with Vibrio Species, and after Being Fed with Florfenicol. Fishes 2023, 8, 148. https://doi.org/10.3390/fishes8030148
Shakweer MS, Elshopakey GE, Abdelwarith AA, Younis EM, Davies SJ, Elbahnaswy S. Comparison of Immune Response of Litopenaeus vannamei Shrimp Naturally Infected with Vibrio Species, and after Being Fed with Florfenicol. Fishes. 2023; 8(3):148. https://doi.org/10.3390/fishes8030148
Chicago/Turabian StyleShakweer, Medhat S., Gehad E. Elshopakey, Abdelwahab A. Abdelwarith, Elsayed M. Younis, Simon John Davies, and Samia Elbahnaswy. 2023. "Comparison of Immune Response of Litopenaeus vannamei Shrimp Naturally Infected with Vibrio Species, and after Being Fed with Florfenicol" Fishes 8, no. 3: 148. https://doi.org/10.3390/fishes8030148