

Population and Transcriptomic Changes of the Tropical Fish Parasite Caligus confusus (Copepoda: Caligidae) with Seasonal Variations in Sea Temperature

 , ,
, ,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Fish Sampling
2.2. Changes in Infection Levels
2.3. Transcriptomic Responses
2.3.1. Acquisition of Copepod Specimens
2.3.2. RNA Extraction, Library Preparation, and Sequencing
2.3.3. De Novo Transcriptome Assembly and Functional Annotation
2.3.4. Evaluation of Assembly
2.3.5. Differentially Expressed Transcripts and Enrichment Analysis
3. Results
3.1. Changes in Infection Levels
3.2. Transcriptomic Responses
De Novo Transcriptome Assembly of C. confusus
3.3. DET and Enrichment Analysis
4. Discussion
4.1. Changes in Infection Levels and Transcriptomic Response
4.2. De Novo Transcriptome Quality
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Correction Statement
References
- Walter, T.C.; Boxshall, G. World of Copepods Database. Available online: https://www.marinespecies.org/copepoda (accessed on 11 May 2023).
- Boxshall, G.A.; Halsey, S.H. An Introduction to Copepod Diversity; Natural History Museum: London, UK, 2004; Volume 15, p. 996. [Google Scholar]
- Abolofia, J.; Asche, F.; Wilen, J.E. The Cost of Lice: Quantifying the Impacts of Parasitic Sea Lice on Farmed Salmon. Mar. Resour. Econ. 2017, 32, 329–349. [Google Scholar] [CrossRef]
- Costello, M.J. The Global Economic Cost of Sea Lice to the Salmonid Farming Industry. J. Fish Dis. 2009, 32, 115–118. [Google Scholar] [CrossRef] [PubMed]
- Hemmingsen, W.; MacKenzie, K.; Sagerup, K.; Remen, M.; Bloch-Hansen, K.; Dagbjartarson Imsland, A.K. Caligus elongatus and other Sea Lice of the Genus Caligus as Parasites of Farmed Salmonids: A Review. Aquaculture 2020, 522, 735160. [Google Scholar] [CrossRef]
- Schram, T. Seasonal Occurrence of Lepeophtheirus salmonis and Caligus elongatus (Copepoda: Caligidae) on Sea Trout (Salmo Trutta), off Southern Norway. ICES J. Mar. Sci. 1998, 55, 163–175. [Google Scholar] [CrossRef]
- Samsing, F.; Oppedal, F.; Dalvin, S.; Johnsen, I.; Vågseth, T.; Dempster, T. Salmon Lice (Lepeophtheirus salmonis) Development Times, Body Size, and Reproductive Outputs Follow Universal Models of Temperature Dependence. Can. J. Fish. Aquat. Sci. 2016, 73, 1841–1851. [Google Scholar] [CrossRef]
- Hamre, L.; Bui, S.; Oppedal, F.; Skern-Mauritzen, R.; Dalvin, S. Development of the Salmon Louse Lepeophtheirus salmonis Parasitic Stages in Temperatures Ranging from 3 to 24 °C. Aquac. Environ. Interact. 2019, 11, 429–443. [Google Scholar] [CrossRef]
- Montory, J.A.; Cumillaf, J.P.; Gebauer, P.; Urbina, M.; Cubillos, V.M.; Navarro, J.M.; Marín, S.L.; Cruces, E. Early Development and Metabolic Rate of the Sea Louse Caligus rogercresseyi under Different Scenarios of Temperature and PCO2. Mar. Environ. Res. 2020, 162, 105154. [Google Scholar] [CrossRef]
- Morales-Serna, F.N.; Rubio-Godoy, M.; Gómez, S. Seasonality of Parasitic Copepods on Bullseye Puffer, Sphoeroides annulatus (Pisces: Tetraodontidae), From the Northwestern Coast of Mexico. J. Parasitol. 2011, 97, 565–573. [Google Scholar] [CrossRef]
- Morales-Serna, F.N.; Olivas-Padilla, L.; Marín-Enriquez, E.; Osuna-Cabanillas, J.M.; Aguirre-Villaseñor, H.; Hernández-Covarrubias, V. Copepod and Monogenean Infection of Fish under Sea Surface Temperature Anomalies. J. Sea Res. 2021, 175, 102098. [Google Scholar] [CrossRef]
- Lõhmus, M.; Björklund, M. Climate Change: What Will It Do to Fish-Parasite Interactions? Biol. J. Linn. Soc. 2015, 116, 397–411. [Google Scholar] [CrossRef]
- Eichner, C.; Frost, P.; Dysvik, B.; Jonassen, I.; Kristiansen, B.; Nilsen, F. Salmon Louse (Lepeophtheirus salmonis) Transcriptomes during Post Molting Maturation and Egg Production, Revealed Using EST-Sequencing and Microarray Analysis. BMC Genom. 2008, 9, 126. [Google Scholar] [CrossRef]
- Chávez-Mardones, J.; Gallardo-Escárate, C. Next-Generation Transcriptome Profiling of the Salmon Louse Caligus Exposed to Deltamethrin (AlphaMaxTM): Discovery of Relevant Genes and Sex-Related Differences. Mar. Biotechnol. 2015, 17, 793–810. [Google Scholar] [CrossRef] [PubMed]
- Poley, J.D.; Igboeli, O.O.; Fast, M.D. Towards a Consensus: Multiple Experiments Provide Evidence for Constitutive Expression Differences among Sexes and Populations of Sea Lice (Lepeophtheirus salmonis) Related to Emamectin Benzoate Resistance. Aquaculture 2015, 448, 445–450. [Google Scholar] [CrossRef]
- Gallardo-Escárate, C.; Valenzuela-Muñoz, V.; Boltaña, S.; Nuñez-Acuña, G.; Valenzuela-Miranda, D.; Gonçalves, A.T.; Détrée, C.; Tarifeño-Saldivia, E.; Farlora, R.; Roberts, S. The Caligus Rogercresseyi MiRNome: Discovery and Transcriptome Profiling during the Sea Lice Ontogeny. Agri Gene 2017, 4, 8–22. [Google Scholar] [CrossRef]
- Morales-Serna, F.N.; Medina-Guerrero, R.M.; Fajer-Avila, E.J. Sea Lice (Copepoda: Caligidae) Parasitic on Fishes Reported from the Neotropical Region. Neotrop. Biodivers. 2016, 2, 141–150. [Google Scholar] [CrossRef]
- Soler-Jiménez, L.C.; Morales-Serna, F.N.; Aguirre-Macedo, M.L.; Mclaughlin, J.P.; Jaramillo, A.G.; Shaw, J.C.; James, A.K.; Hechinger, R.F.; Kuris, A.M.; Lafferty, K.D.; et al. Parasitic Copepods (Crustacea, Hexanauplia) on Fishes from the Lagoon Flats of Palmyra Atoll, Central Pacific. ZooKeys 2019, 833, 85–106. [Google Scholar] [CrossRef]
- Sanchez-Cabeza, J.-A.; Herrera-Becerril, C.A.; Carballo, J.L.; Yáñez, B.; Álvarez-Sánchez, L.F.; Cardoso-Mohedano, J.-G.; Ruiz-Fernández, A.C. Rapid Surface Water Warming and Impact of the Recent (2013–2016) Temperature Anomaly in Shallow Coastal Waters at the Eastern Entrance of the Gulf of California. Prog. Oceanogr. 2022, 202, 102746. [Google Scholar] [CrossRef]
- Ho, J.S.; Lin, C.L. Sea Lice of Taiwan; The Sueichan Press: Keelung, Taiwan, 2004. [Google Scholar]
- Bush, A.O.; Lafferty, K.D.; Lotz, J.M.; Shostak, A.W. Parasitology Meets Ecology on its Own Terms: Margolis et al. Revisited. J. Parasitol. 1997, 83, 575–583. [Google Scholar] [CrossRef]
- Reiczigel, J.; Marozzi, M.; Fábián, I.; Rózsa, L. Biostatistics for Parasitologists–A Primer to Quantitative Parasitology. Trends Parasitol. 2019, 35, 277–281. [Google Scholar] [CrossRef]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. 2010. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 15 June 2023).
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-Length Transcriptome Assembly from RNA-Seq Data without a Reference Genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef]
- Fu, L.; Niu, B.; Zhu, Z.; Wu, S.; Li, W. CD-HIT: Accelerated for Clustering the next-Generation Sequencing Data. Bioinformatics 2012, 28, 3150–3152. [Google Scholar] [CrossRef]
- Haas, B.J.; Papanicolaou, A.; Yassour, M.; Grabherr, M.; Blood, P.D.; Bowden, J.; Couger, M.B.; Eccles, D.; Li, B.; Lieber, M.; et al. De Novo Transcript Sequence Reconstruction from RNA-Seq Using the Trinity Platform for Reference Generation and Analysis. Nat. Protoc. 2013, 8, 1494–1512. [Google Scholar] [CrossRef]
- Bryant, D.M.; Johnson, K.; DiTommaso, T.; Tickle, T.; Couger, M.B.; Payzin-Dogru, D.; Lee, T.J.; Leigh, N.D.; Kuo, T.-H.; Davis, F.G.; et al. A Tissue-Mapped Axolotl De Novo Transcriptome Enables Identification of Limb Regeneration Factors. Cell Rep. 2017, 18, 762–776. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, T.J.; Eddy, S.R. Nhmmer: DNA Homology Search with Profile HMMs. Bioinformatics 2013, 29, 2487–2489. [Google Scholar] [CrossRef]
- Petersen, T.N.; Brunak, S.; Heijne, G.; Nielsen, H. SignalP 4.0: Discriminating Signal Peptides from Transmembrane Regions. Nat. Methods 2011, 8, 785–786. [Google Scholar] [CrossRef] [PubMed]
- Krogh, A.; Larsson, B.; Von Heijne, G.; Sonnhammer, E.L.L. Predicting Transmembrane Protein Topology with a Hidden Markov Model: Application to Complete Genomes. J. Mol. Biol. 2001, 305, 567–580. [Google Scholar] [CrossRef] [PubMed]
- The UniProt Consortium. UniProt: The universal protein knowledgebase in 2021. Nucleic Acids Res. 2021, 49, D480–D489. [Google Scholar] [CrossRef]
- Buchfink, B.; Xie, C.; Huson, D.H. Fast and Sensitive Protein Alignment Using DIAMOND. Nat. Methods 2015, 12, 59–60. [Google Scholar] [CrossRef]
- Caña-Bozada, V.; Morales-Serna, F.N.; Fajer-Ávila, E.J.; Llera-Herrera, R. De Novo Transcriptome Assembly and Identification of G-Protein-Coupled-Receptors (GPCRs) in Two Species of Monogenean Parasites of Fish. Parasite 2022, 29, 51. [Google Scholar] [CrossRef]
- Manni, M.; Berkeley, M.R.; Seppey, M.; Zdobnov, E.M. BUSCO: Assessing Genomic Data Quality and Beyond. Curr. Protoc. 2021, 1, e323. [Google Scholar] [CrossRef] [PubMed]
- Bray, N.L.; Pimentel, H.; Melsted, P.; Pachter, L. Near-Optimal Probabilistic RNA-Seq Quantification. Nat. Biotechnol. 2016, 34, 525–527. [Google Scholar] [CrossRef] [PubMed]
- Baldoni, P.L.; Chen, Y.; Hediyeh-zadeh, S.; Liao, Y.; Dong, X.; Ritchie, M.E.; Shi, W.; Smyth, G.K. Dividing out Quantification Uncertainty Allows Efficient Assessment of Differential Transcript Expression. bioRxiv 2023. [Google Scholar] [CrossRef]
- Alexa, A.; Rahnenführer, J. Gene Set Enrichment Analysis with TopGO. Bioconductor Improv. 2009, 27, 1–26. [Google Scholar]
- Korotkevich, G.; Sukhov, V.; Budin, N.; Shpak, B.; Artyomov, M.N.; Sergushichev, A. Fast Gene Set Enrichment Analysis. bioXiv 2016. [Google Scholar] [CrossRef]
- Rikardsen, A.H. Seasonal Occurrence of Sea Lice Lepeophtheirus salmonis on Sea Trout in Two North Norwegian Fjords. J. Fish Biol. 2004, 65, 711–722. [Google Scholar] [CrossRef]
- Khamees, N.R.; Adday, T.K. Occurrence of Sea Lice Caligus Epinepheli Yamaguti, 1936 (Copepoda: Siphonostomatoida) on Gills of Nemipterus japonicus (Bloch, 1775) from Northwest of the Arabian Gulf. Basrah J. Agric. Sci. 2013, 26, 1–14. [Google Scholar] [CrossRef]
- Montes, R.M.; Quiñones, R.A.; Gallardo-Escárate, C. Disentangling the Effect of Sea Temperature and Stocking Density on Sea Louse Abundance Using Wavelets in a Highly Infested Salmon Farming Area. Aquaculture 2022, 546, 737246. [Google Scholar] [CrossRef]
- Nguyen, K.D.T.; Morley, S.A.; Lai, C.-H.; Clark, M.S.; Tan, K.S.; Bates, A.E.; Peck, L.S. Upper Temperature Limits of Tropical Marine Ectotherms: Global Warming Implications. PLoS ONE 2011, 6, e29340. [Google Scholar] [CrossRef]
- Doan, N.X.; Vu, M.T.T.; Pham, H.Q.; Wisz, M.S.; Nielsen, T.G.; Dinh, K.V. Extreme Temperature Impairs Growth and Productivity in a Common Tropical Marine Copepod. Sci. Rep. 2019, 9, 4550. [Google Scholar] [CrossRef]
- Bravo, S.; Erranz, F.; Lagos, C. A Comparison of Sea Lice, Caligus rogercresseyi, Fecundity in Four Areas in Southern Chile. J. Fish Dis. 2009, 32, 107–113. [Google Scholar] [CrossRef] [PubMed]
- King, H.R.; Pankhurst, N.W.; Watts, M.; Pankhurst, P.M. Effect of Elevated Summer Temperatures on Gonadal Steroid Production, Vitellogenesis and Egg Quality in Female Atlantic Salmon: Vitellogenesis in atlantic salmon. J. Fish Biol. 2003, 63, 153–167. [Google Scholar] [CrossRef]
- Lee, C.S.; O’Bryen, P.J.; Marcus, N.H. Copepods in Aquaculture; John Wiley & Sons: Hoboken, NJ, USA, 2008. [Google Scholar]
- Tarrant, A.M.; Nilsson, B.; Hansen, B.W. Molecular Physiology of Copepods-from Biomarkers to Transcriptomes and Back Again. Comp. Biochem. Physiol. Part D Genom. Proteom. 2019, 30, 230–247. [Google Scholar] [CrossRef] [PubMed]
- Semmouri, I.; Asselman, J.; Van Nieuwerburgh, F.; Deforce, D.; Janssen, C.R.; De Schamphelaere, K.A.C. The Transcriptome of the Marine Calanoid Copepod Temora longicornis under Heat Stress and Recovery. Mar. Environ. Res. 2019, 143, 10–23. [Google Scholar] [CrossRef]
- Dalvin, S.; Frost, P.; Loeffen, P.; Skern-Mauritzen, R.; Baban, J.; Rønnestad, I.; Nilsen, F. Characterisation of Two Vitellogenins in the Salmon Louse Lepeophtheirus salmonis: Molecular, Functional and Evolutional Analysis. Dis. Aquat. Organ. 2011, 94, 211–224. [Google Scholar] [CrossRef]
- Zhai, C.; Zhang, N.; Li, X.; Chen, X.; Sun, F.; Dong, M. Fusion and Expansion of Vitellogenin Vesicles during Caenorhabditis elegans Intestinal Senescence. Aging Cell 2022, 21, e13719. [Google Scholar] [CrossRef]
- Tang, J.; Ma, Y.-C.; Chen, Y.-L.; Yang, R.-Q.; Liu, H.-C.; Wang, X.; Ni, B.; Zou, C.-G.; Zhang, K.-Q. Vitellogenin Accumulation Leads to Reproductive Senescence by Impairing Lysosomal Function. Sci. China Life Sci. 2023, 66, 439–452. [Google Scholar] [CrossRef]
- Luchian, I.; Goriuc, A.; Sandu, D.; Covasa, M. The Role of Matrix Metalloproteinases (MMP-8, MMP-9, MMP-13) in Periodontal and Peri-Implant Pathological Processes. Int. J. Mol. Sci. 2022, 23, 1806. [Google Scholar] [CrossRef]
- Pagemccaw, A. Remodeling the Model Organism: Matrix Metalloproteinase Functions in Invertebrates. Semin. Cell Dev. Biol. 2008, 19, 14–23. [Google Scholar] [CrossRef]
- Li, R.; Meng, Q.; Huang, J.; Wang, S.; Sun, J. MMP-14 Regulates Innate Immune Responses to Eriocheir Sinensis via Tissue Degradation. Fish Shellfish Immunol. 2020, 99, 301–309. [Google Scholar] [CrossRef]
- Jia, Q.; Chen, X.; Wu, L.; Ruan, Z.; Li, K.; Li, S. Matrix Metalloproteinases Promote Fat Body Cell Dissociation and Ovary Development in Bombyx Mori. J. Insect Physiol. 2018, 111, 8–15. [Google Scholar] [CrossRef]
- Naqvi, M.A.-H.; Li, H.; Gao, W.; Naqvi, S.Z.; Jamil, T.; Aimulajiang, K.; Xu, L.; Song, X.; Li, X.; Yan, R. Haemonchus contortus: SiRNA Mediated Knockdown of Matrix Metalloproteinase 12A (MMP-12) Results in Reduction of Infectivity. Parasit. Vectors 2020, 13, 151. [Google Scholar] [CrossRef]
- Widlansky, M.J.; Long, X.; Schloesser, F. Increase in Sea Level Variability with Ocean Warming Associated with the Nonlinear Thermal Expansion of Seawater. Commun. Earth Environ. 2020, 1, 9. [Google Scholar] [CrossRef]
- Dalvin, S.; Are Hamre, L.; Skern-Mauritzen, R.; Vågseth, T.; Stien, L.; Oppedal, F.; Bui, S. The Effect of Temperature on Ability of Lepeophtheirus Salmonis to Infect and Persist on Atlantic Salmon. J. Fish Dis. 2020, 43, 1519–1529. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Egusa, T.; Fukushima, K.; Oda, T.; Ohte, N.; Tokuchi, N.; Watanabe, K.; Kanaiwa, M.; Murakami, I.; Lafferty, K.D. Nematomorph Parasites Indirectly Alter the Food Web and Ecosystem Function of Streams through Behavioural Manipulation of Their Cricket Hosts. Ecol. Lett. 2012, 15, 786–793. [Google Scholar] [CrossRef]
- Dobson, A.; Lafferty, K.D.; Kuris, A.M.; Hechinger, R.F.; Jetz, W. Homage to Linnaeus: How Many Parasites? How Many Hosts? Proc. Natl. Acad. Sci. USA 2008, 105 (Suppl. S1), 11482–11489. [Google Scholar] [CrossRef]
- Bowman, L.L.; Kondrateva, E.S.; Timofeyev, M.A.; Yampolsky, L.Y. Temperature Gradient Affects Differentiation of Gene Expression and SNP Allele Frequencies in the Dominant Lake Baikal Zooplankton Species. Mol. Ecol. 2018, 27, 2544–2559. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Ye, F.; Sheng, Q.; Clark, T.; Samuels, D.C. Three-Stage Quality Control Strategies for DNA Re-Sequencing Data. Brief. Bioinform. 2014, 15, 879–889. [Google Scholar] [CrossRef] [PubMed]
- Jørgensen, T.S.; Petersen, B.; Petersen, H.C.B.; Browne, P.D.; Prost, S.; Stillman, J.H.; Hansen, L.H.; Hansen, B.W. The Genome and MRNA Transcriptome of the Cosmopolitan Calanoid Copepod Acartia tonsa Dana Improve the Understanding of Copepod Genome Size Evolution. Genome Biol. Evol. 2019, 11, 1440–1450. [Google Scholar] [CrossRef]
- Mathieu-Bégné, E.; Loot, G.; Blanchet, S.; Toulza, E.; Genthon, C.; Rey, O. De Novo Transcriptome Assembly for Tracheliastes polycolpus, an Invasive Ectoparasite of Freshwater Fish in Western Europe. Mar. Genomics 2019, 46, 58–61. [Google Scholar] [CrossRef]
- Carmona-Antoñanzas, G.; Carmichael, S.N.; Heumann, J.; Taggart, J.B.; Gharbi, K.; Bron, J.E.; Bekaert, M.; Sturm, A. A Survey of the ATP-Binding Cassette (ABC) Gene Superfamily in the Salmon Louse (Lepeophtheirus salmonis). PLoS ONE 2015, 10, e0137394. [Google Scholar] [CrossRef] [PubMed]
- Pallavi, B.; Shankar, K.M.; Abhiman, P.B.; Iqlas, A. Identification of Putative Genes Involved in Parasitism in the Anchor Worm, Lernaea cyprinacea by de Novo Transcriptome Analysis. Exp. Parasitol. 2015, 153, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Gallardo-Escárate, C.; Valenzuela-Muñoz, V.; Nuñez-Acuña, G. RNA-Seq Analysis Using De Novo Transcriptome Assembly as a Reference for the Salmon Louse Caligus rogercresseyi. PLoS ONE 2014, 9, e92239. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
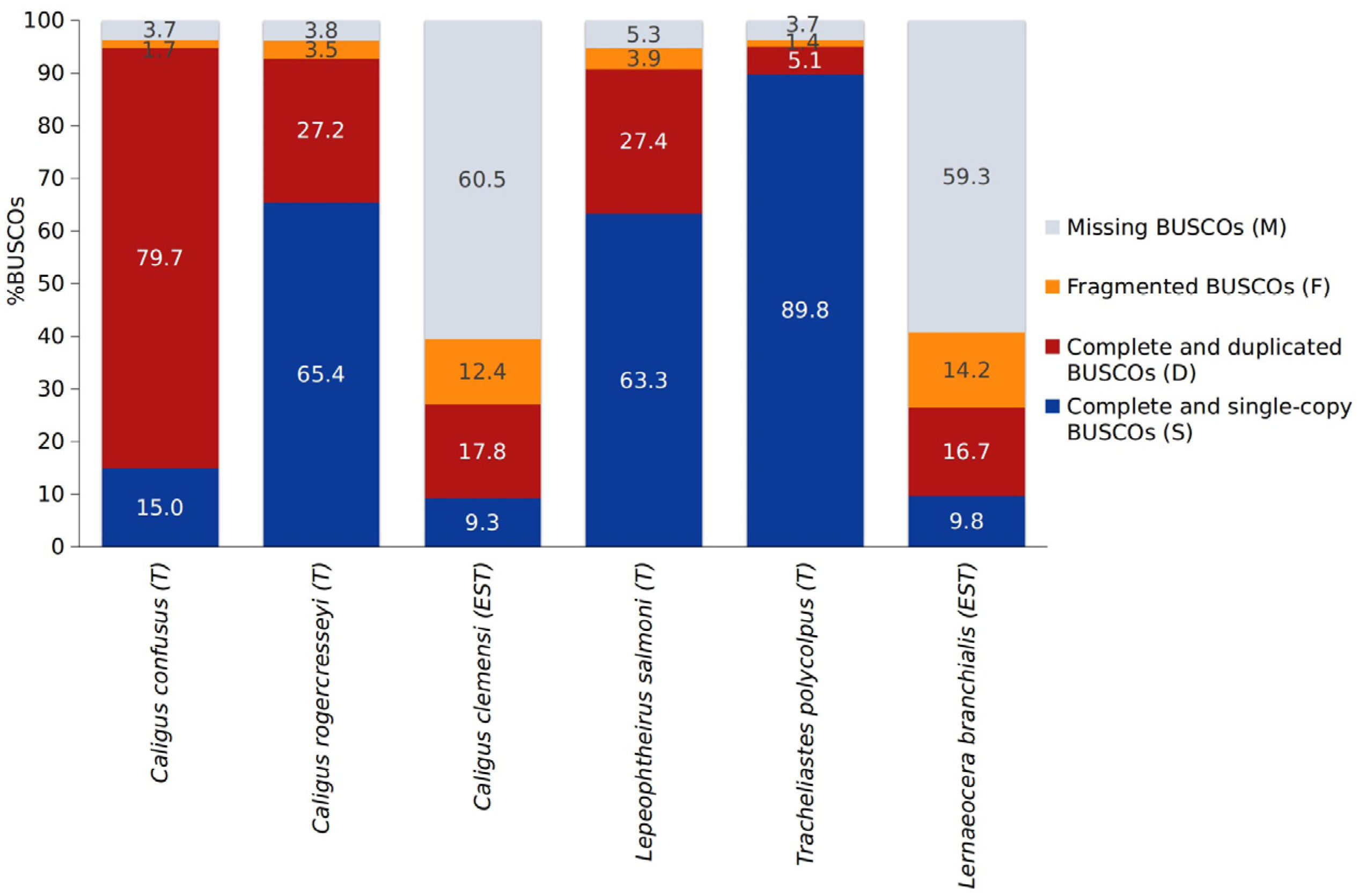{kind=link}
{kind=link}
{kind=link}
| Descriptors | De Novo Assembly |
|---|---|
| Reads (millions) | 893 |
| Trinity transcripts (n) | 230,861 |
| Trinity predicted genes (n) | 182,298 |
| Assembled bases | 233,706,455 |
| Average contig length (bp) | 1012.3 |
| Median contig length (bp) | 732 |
| N10 (bp) | 3841 |
| N50 (bp) | 1000 |
| GC content (%) | 36.96 |
| Overall mapping (%) | 35 |
| Annotation (total) | |
| Predicted ORF | 55,530 |
| Swiss-Prot top BLASTX hit | 44,902 |
| Gene Ontology in Blastx | 44,280 |
| Kegg | 39,021 |
| Swiss-Prot top BLASTP hit | 31,478 |
| Pfam | 30,596 |
| Gene Ontology in blastp | 31,050 |
| Gene Ontology in Pfam | 18,751 |
| SignalP | 5504 |
| Eggnog | 479 |
| Copepod Species | Caligus confusus | Tracheliastes polycolpus | Lepeophtheirus salmonis | Lernaea cyprinacea | Caligus rogercresseyi |
|---|---|---|---|---|---|
| Reference | This study | [65] | [66] | [67] | [68] |
| Sequencing platform | MGIseq 2000 | Illumina HiSeq 2000 | Illumina HiSeq 2000 | Illumina HiSeq 2000 | Illumina MiSeq |
| Assembly software | Trinity 2.6.6 | Trinity version 2014-07-17 | Trinity version 2012-06-08 | Velvet_1.2.10 and Oases_0.2.08 | CLC Genomics Workbench |
| Raw reads (millions) | 433.7 | 420 | 389.92 | 31.67 | 151.78 |
| Assembled bases | 233,706,455 | 20,355,465 | * | 50,792 | 83,444 |
| Transcripts filtered | 230,861 | 17,157 | 33,933 | 57,349 | 83,444 |
| Average contig length (bp) | 1012.3 | 1186 | 1469 | 1071.2 | 819 |
| Median contig length | 732 | * | 979 | 23,727 | * |
| N50 (bp) | 1000 | 1599 | 2100 | 1750 | * |
| Overall mapping (%) | 35 | 65.8 | * | * | * |
| GC content (%) | 36.96 | 38.55 | 36.9 | * | * |
| Transcripts with GO terms | 32,152 | 12,098 | 28,547 | 2439 | * |
| Transcripts with KEGG terms | 39,021 | 10,924 | * | * | * |
| Pfam | 30,596 | 11,751 | * | * | * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pimentel-Acosta, C.A.; Caña-Bozada, V.H.; Osuna-Cabanillas, J.M.; Fajer-Ávila, E.J.; Ovando-Vásquez, C.; Morales-Serna, F.N. Population and Transcriptomic Changes of the Tropical Fish Parasite Caligus confusus (Copepoda: Caligidae) with Seasonal Variations in Sea Temperature. Fishes 2023, 8, 475. https://doi.org/10.3390/fishes8100475
Pimentel-Acosta CA, Caña-Bozada VH, Osuna-Cabanillas JM, Fajer-Ávila EJ, Ovando-Vásquez C, Morales-Serna FN. Population and Transcriptomic Changes of the Tropical Fish Parasite Caligus confusus (Copepoda: Caligidae) with Seasonal Variations in Sea Temperature. Fishes. 2023; 8(10):475. https://doi.org/10.3390/fishes8100475
Chicago/Turabian StylePimentel-Acosta, Citlalic Altagracia, Víctor Hugo Caña-Bozada, Juan Manuel Osuna-Cabanillas, Emma Josefina Fajer-Ávila, Cesaré Ovando-Vásquez, and Francisco Neptalí Morales-Serna. 2023. "Population and Transcriptomic Changes of the Tropical Fish Parasite Caligus confusus (Copepoda: Caligidae) with Seasonal Variations in Sea Temperature" Fishes 8, no. 10: 475. https://doi.org/10.3390/fishes8100475