1. Introduction
Crustacean culture represents 12% of the aquaculture production by species. The shrimp species is one of the most important aquaculture species, with an estimated production in 2016 of 5,180,563 tons, of which, 4,155,827 tons were
Litopenaeus vannamei [
1]. Whiteleg shrimps need a high percentage of protein in their diet. Dietary protein in shrimp feed ranges from 25% to 33% [
2], of which, a minimum of 12% is fish meal [
3]. Fish meal is an essential ingredient in fish/crustaceans feed, but is an unsustainable source due to overfishing and the increased demand for livestock feed [
4]. Hence, a decrease in the fish meal included in shrimp feed and an increase in the employed alternative protein are foreseen [
5]. In fact, aquaculture depends increasingly less on fishmeal. The FIFO (fish in–fish out) lowered from 0.63 in 2000 to 0.22 in 2015, with 0.91 for crustaceans in 2000 and 0.46 in 2015 [
6].
The alternative proteins used in aquafeeds are mainly derived from plants. Vegetable sources have some advantages over animal-based sources, such as lower prices, more availability, and improved composition [
7,
8]. Of all plant sources, soy is the most widely used because of its excellent nutritional quality, high digestibility, high protein quality and quantity, and the best amino acid profile of available vegetable protein sources. Despite its good nutritional value, the inclusion percentages do not exceed 40% [
9] due to antinutritional factors that reduce feed utilization, absorption, and feed conversion ratio, as these sometimes affect the growth of shrimps. The bioprocessing of soy could eliminate these antinutritional factors. On the other hand, soy meal has a strong environmental impact, such as its contribution to deforestation, the use of water, and the utilization of GMOs and pesticides [
10]. Thus, the search for other sustainable sources that allow higher replacement percentages and diversify aquafeed ingredients remains as an important animal nutrition research goal.
Plukenetia volubilis (sacha inchi, SI) is a partly woody and perennial plant belongs to the Euphorbiaceous family, which is native to Peruvian jungles. The flour from the seeds contains approximately 48% oil and 27% proteins, which are rich in cysteine, tyrosine, threonine, and tryptophan [
11]. The fatty acids of sacha inchi are mostly unsaturated, with about 85% of polyunsaturation, and with an n6/n3 ratio, reported by Hamaker et al. [
12], of approximately 0.81, comprising approximately 33–34% linoleic acid and 50–51% linolenic acid [
12,
13]. These fatty acids are important for health and nutrition, and for the prevention of cardiovascular disease, rheumatoid arthritis, and cancer [
13,
14]. These unsaturated fatty acids are also important for the development of aquatic organisms, and are the main source of high amounts of unsaturated fatty acids for humans. Other bioactive compounds have also been observed in SI oil, such as carotenes [
15], polyphenolic compounds [
15,
16], tocopherols [
15,
17,
18], and phytosterols [
18]. For all these reasons, SI seeds should be considered as an important dietary source of health-promoting phytochemicals [
15]. Currently, SI oil is commercialized for medicinal applications for skin care and as a nutritional supplement for humans. SI oil is obtained by pressing nuts, separating oil, and obtaining a rich protein paste by-product which can be used in animal feed. The post-oil extraction residue from sacha inchi presents a high content of proteins (59%) [
19]. This cake represents 62–70% of seed and contains 12–17% of oil [
20]. SI cake has been examined as a fish meal substitute for several fish species, such as red tilapia (
Tilapia spp.) [
21],
Colossoma macropomum,
Brycon amazonicus [
22], and rainbow trout (
Oncorhynchus mykiss) [
23], all with promising results. In rainbow trout, SI digestibility is better than other vegetable sources [
23], and, depending on the animal species, SI can be included in the diet at 10–30% without affecting growth rates [
21,
22].
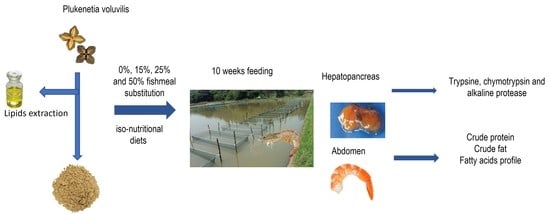
Given the above considerations, the aim of this work was to study the effect of the replacement of fish meal by Plukenetia volubilis on whiteleg shrimp growth, nutrition indices, digestive enzymes, and final body composition.
2. Materials and Methods
2.1. Sacha Inchi Cake
The sacha inchi (
P. volubilis) seeds were provided by the agricultural cooperative, “Aso-Inchi Cantón Las Lajas”, in the province of El Oro (Ecuador). To obtain sacha inchi cake, seeds were extruded at a low temperature (below 45 °C) to extract the sacha inchi oil. The amino acid profile of the sacha inchi cake was obtained through the Protein Chemistry Service of the Biological Research Centre (CSIC, Madrid, Spain), following the basic principle of operation developed by Spackman et al. [
24], by continuous flow chromatography (Biochrom 30 series). Tryptophan was not detected, and asparagine and glutamine were deaminated and detected as aspartic acid and glutamic acid, respectively (
Table 1,
Figure S1).
2.2. Experimental Diets
Four isonitrogenous (350 g crude protein kg
−1) diets were prepared in the Department of Aquaculture at the Technical University of Machala, Ecuador. Experimental diets were formulated, including sacha inchi cake as a partial substitute for fish meal. In order to formulate isonitrogenous diets, the quantity of soy meal and squid meal varied among the diets. A sacha-inchi-free diet was used as the control (CT), and three experimental diets were designed with increasing levels of fish meal replacement by sacha inchi cake: 15% (D-15), 25% (D-25), and 50% (D-50) regarding the fish meal. Briefly, feed ingredients, including vitamins and minerals, were finely ground, weighed, and mixed in a vertical helix ribbon mixer (Bathammex 178716, 10-L capacity, Bathammex, Mexico DF, Mexico). Then fish oil was added to the latter ingredients to be mixed together. Water was slowly added until the diets began to clump and form a homogeneous dough. The dough was then extruded by a mill machine (Torrey M-22R1, Torrey, Mexico DF, Mexico) to obtain pellets that were 4 mm in diameter. Finally, the pellets were dried at room temperature for 24 h. The pellets were broken and sieved with sieves of different mesh sizes, and kept in sealed plastic bags at −20 °C until their use. The formulation and chemical composition of the experimental diets are shown in
Table 2 and
Table 3, respectively.
2.3. Feeding Trial and Sampling
The feeding trial was carried out in the shrimp farm, “Noblecilla Salas”, located in Santa Rosa, El Oro Province, Ecuador (3°23′45″ S; 79°57′21″ W), using 1 m3 cages placed in a 1-hectare pond. Prior to the experiment, the shrimps were acclimatized to the culture environment for 3 weeks and fed a commercial diet (Nicovita Shrimp feed). Once acclimatized, 120 L. vannamei juveniles were used, with an average initial weight of 3.41 ± 0.31 g. The shrimps were randomly distributed and stocked in eight cubical 1 m3 cages at the bottom of pond at a density of 15 individuals per cage. Each experimental feed was tested in duplicate (4 diets × 2 cages each) for 10 weeks. All the shrimp groups were fed with the experimental diets 3 times daily (7% of their body weight) at 08:00 h, 12:00 h, and 16:00 h. Every week, the total weight of the shrimps in each cage was registered to adjust the daily amount of feed. The water quality parameters measured during the experimental trial were optimal for the growth and survival of white shrimp and averaged: temperature (26.6 °C), salinity (24.5‰), pH (7.19), and O2 (4–5 mg/mL). A natural light–dark cycle was used during the trial.
Weekly, the shrimps in each cage were individually weighed, and the standard length was measured to determine the growth and the feed utilization parameters. At the end of the feeding trial, the crude protein, total lipid content, and fatty acid profile of the abdomen samples were determined by standard methods, as described in
Section 2.6 Finally, for the enzymatic activity analysis, the hepatopancreas from 10 shrimps per treatment were pooled to obtain enzymatic extracts. Briefly, hepatopancreas were manually homogenized in distilled water at 4 °C to a final concentration of 0.5 g mL
−1, and supernatants were obtained after centrifugation (12,000 rpm, 12 min, 4 °C). The total soluble protein in the enzyme extracts was determined according to Bradford [
25], using bovine serum albumin as the standard.
2.4. Animals Ethics
Crustaceans are not regulated by European animal welfare legislation [
26]. However, the animals were kept and slaughtered under production conditions; they were immersed in ice and then frozen. During the experimental period, they were not subjected to any procedure, and all analyses were carried out post-mortem.
2.5. Growth Performance and Nutrient Utilization
Final weight and feed intake were used to calculate the different parameters: specific growth rate (SGR, %) = (Ln (Wf) − Ln (Wi)/days) × 100, where Wf and Wi were the final and the initial weight, respectively, (g); daily gain (DG, g day−1) = (Wf − Wi)/days; feed conversion ratio (FCR) = total feed intake as a dry basis (g)/weight gain (g).
Cephalotorax, abdomen, abdomen + exoskeleton, and hepatopancreas were weighted, and the total length of the cephalothorax and abdomen were measured.
2.6. Proximate Composition and Fatty Acids Analysis
Chemical analysis of the sacha inchi diets and the edible part of the abdomen (without the exoskeleton) samples were conducted by AOAC [
27] procedures. Dry matter and ash were determined gravimetrically after drying at 105 ± 0.5 °C and after combustion at 500 °C in a muffle furnace, respectively, until a constant weight. Crude protein content was determined by the Kjeldahl method (Nx.6.25). Total lipids were determined by ethyl ether extraction (Soxhlet technique). The fatty acid rofile of the sacha inchi cake, feeds, and abdomen without the exoskeleton were determined by gas chromatography using a modification of the direct transesterification method described by Lepage and Roy [
28] that requires no prior separation of the lipid fraction [
29].
All the analyses were performed in triplicate.
2.7. Digestive Enzymes Activity
The total alkaline protease activity in the hepatopancreas extracts was determined according to Alarcon et al. [
30], using 5 g L
−1 of casein in 50 mM of Tris HCl (pH 9.0) as the substrate. One unit of total protease activity was defined as the amount of enzyme that released 1 µg of tyrosine per min in the reaction mixture by considering an extinction coefficient for tyrosine of 0.008 µg
−1 mL
−1 cm
−1, measured at 280 nm. Trypsin and chymotrypsin activities were measured using 0.5 mM BAPNA (N-a-benzoyl-DL-arginine-4-nitroanilide) as the substrate according to Erlanger et al. [
31], and 0.2 mM SAPNA (N-succinyl-(Ala)2-Pro-Phe-P-nitroanilide) according to Del Mar et al. [
32], respectively, in 50 mM Tris-HCl buffer, pH 8.5, containing 10 mM CaCl
2. For trypsin and chymotrypsin, one unit of enzyme activity (U) was defined as the amount of enzyme that released 1 µmol of p-nitroanilide per min, using 8800 M cm
−1 as the extinction coefficient, measured at 405 nm. The amylase activity was determined according to the Somogyi–Nelson method using soluble starch (2%
w/
v) as the substrate, as described by Robyt and Whelan [
33]. One unit of activity (U) was defined as the amount of enzyme capable of producing 1 mg of maltose per minute.
All the assays were performed in triplicate, and the specific enzymatic activity was expressed as U g tissue−1.
2.8. Statistical Analysis
All the statistical analyses were performed with the Stagraphics Plus 4.0 (Rockville, MD, USA) software. The results were expressed as the mean ± standard deviation of at least three determinations. The data with parametric distribution were analyzed by a one-way analysis of variance (ANOVA), and the significant differences between treatments (p < 0.05) were determined by Tukey’s multiple comparison test. The data with nonparametric distribution were analyzed by the Kruskal–Wallis test, and significant differences were determined using box and whisker plot graphs.

 ,
, 


{kind=link}
{kind=link}