
Effects of Replacing Dietary Fish Meal by Soybean Meal Co-Fermented Using Bacillus subtilis and Enterococcus faecium on Serum Antioxidant Indices and Gut Microbiota of Crucian Carp Carassius auratus

Abstract
:1. Introduction
2. Materials and Methods
2.1. Source of Fermentation Strain and Fermentation Process
2.2. Preparation of FSM
2.3. Experimental Design and Experimental Feed Preparation
2.4. Experimental Conditions
2.5. Sample Collection
2.6. Determination of Serum Indices of C. auratus
2.7. Analysis of GM Composition
2.8. Data Analysis
3. Results
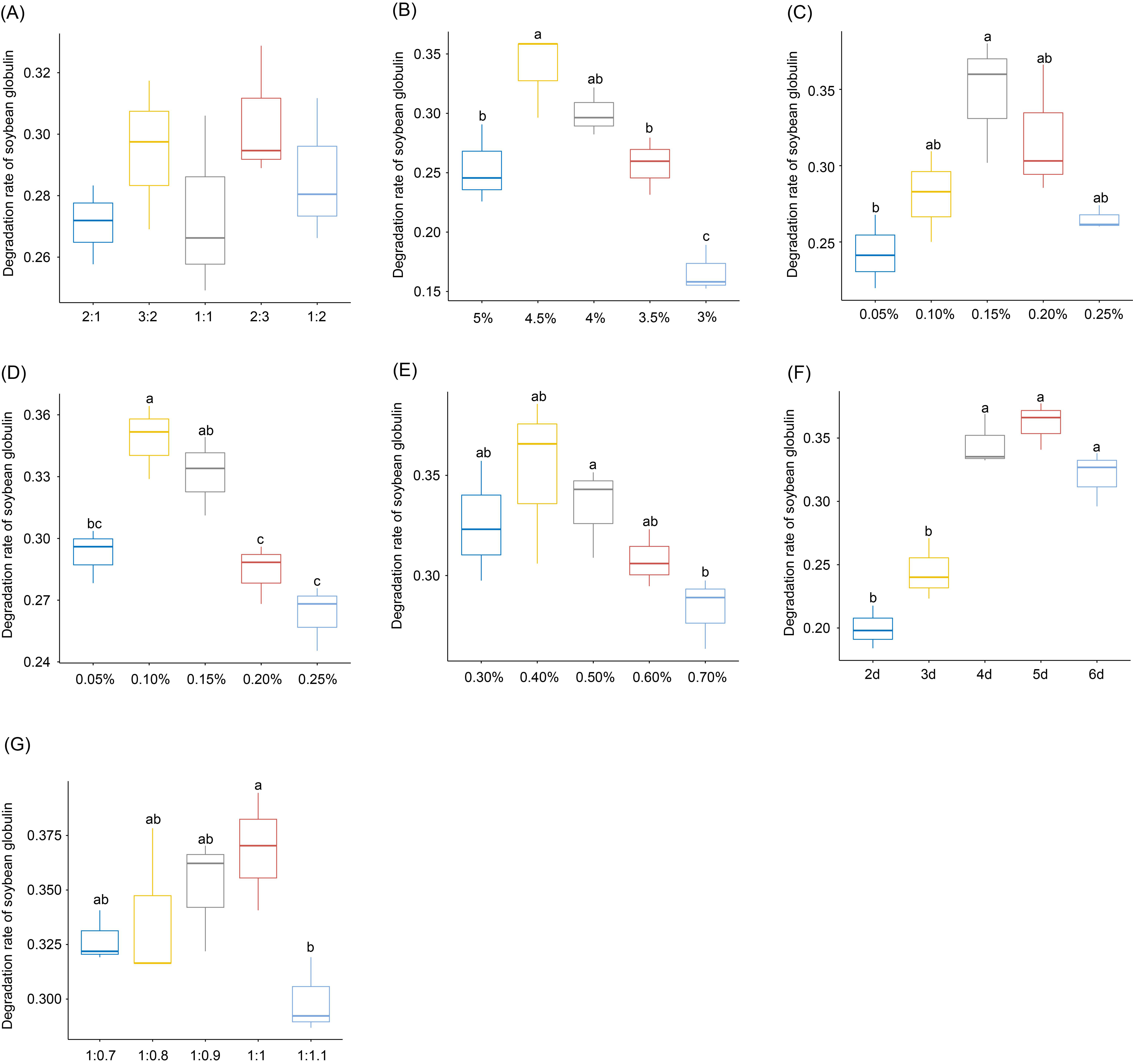
3.1. Optimization Results of Fermentation Conditions
3.2. Effects of Substituting FSM for Fish Meal on Serum Antioxidant Indices of C. auratus
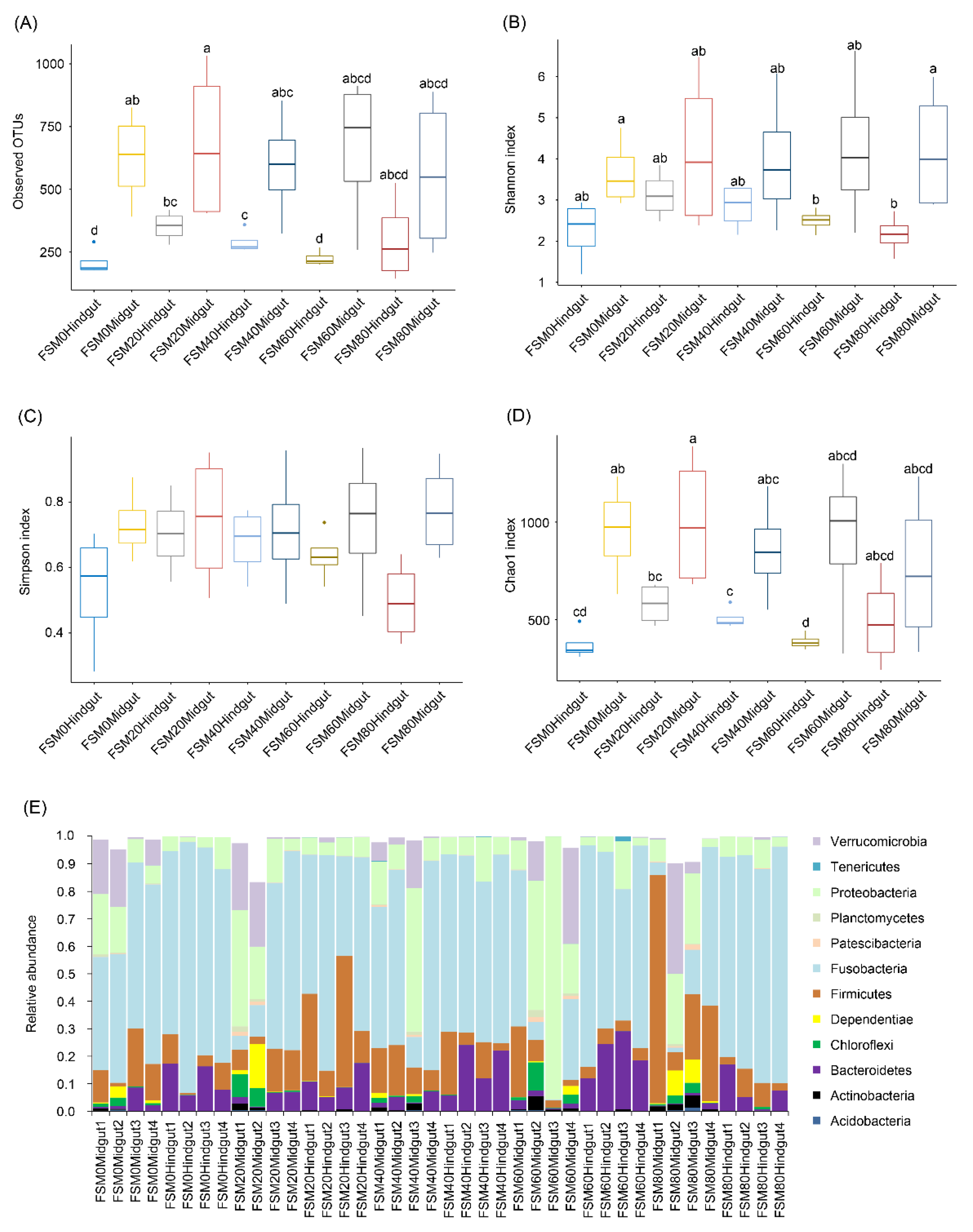
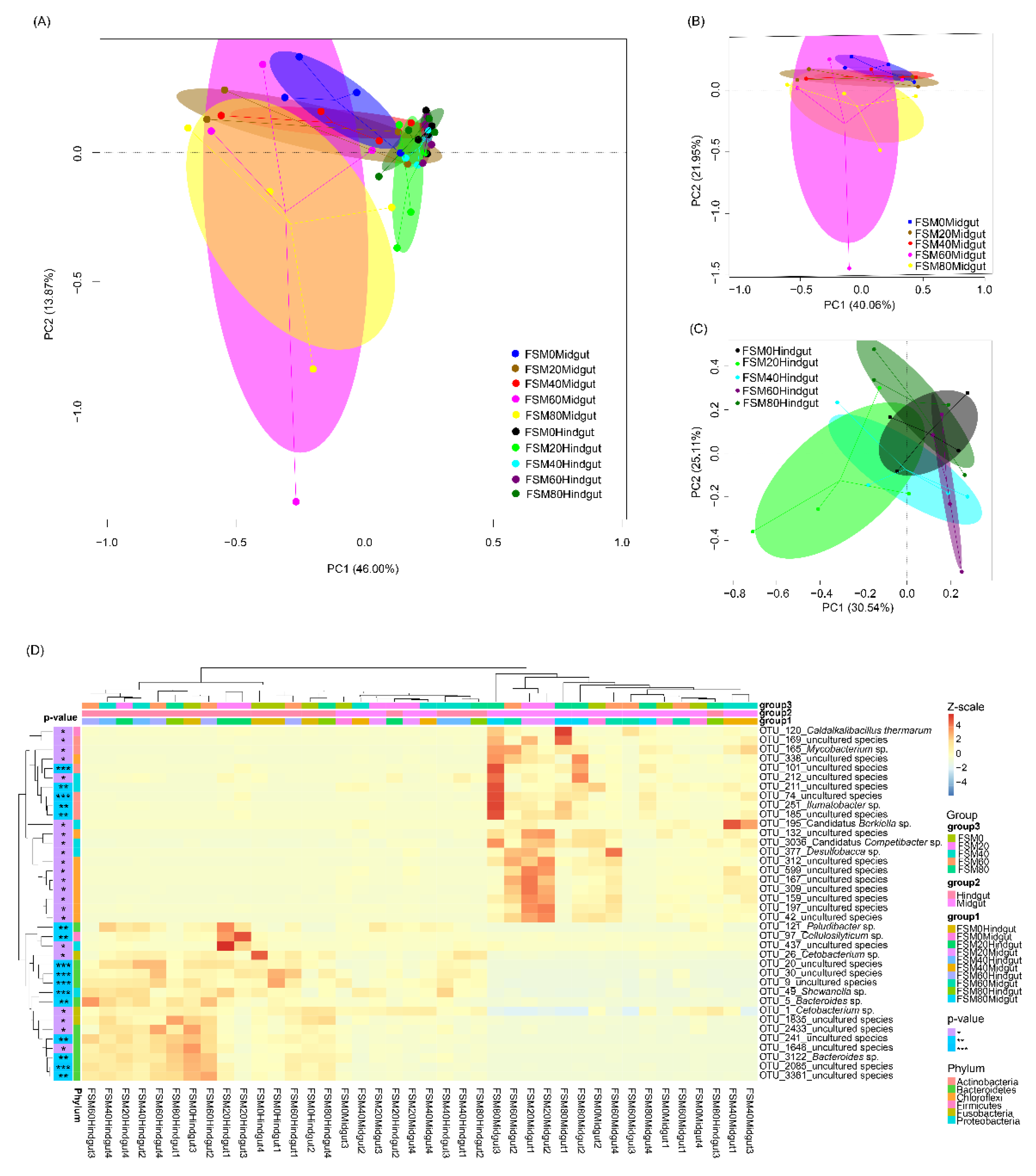
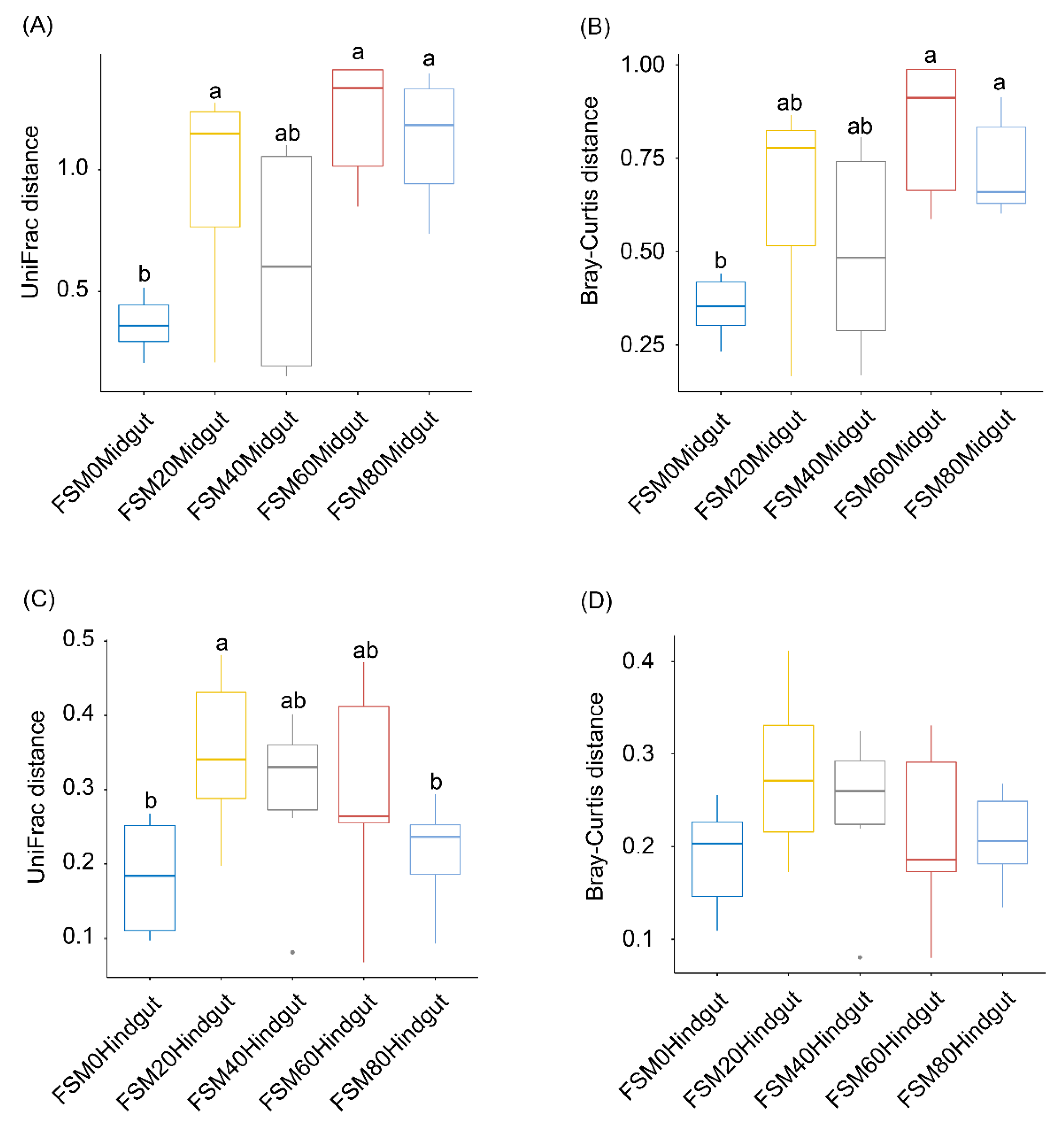
3.3. Effects of Replacing Fish Meal with FSM on GM Composition of C. auratus
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gong, W.; Gao, S.; Zhu, Y.; Wang, G.; Zhang, K.; Li, Z.; Yu, E.; Tian, J.; Xie, Y.; Xie, J.; et al. Effect of the aerobic denitrifying bacterium Pseudomonas furukawaii ZS1 on microbiota compositions in grass carp culture water. Water 2021, 13, 1329. [Google Scholar] [CrossRef]
- FAO. The State of World Fisheries and Aquaculture 2020. Sustainability in Action; FAO: Rome, Italy, 2020. [Google Scholar] [CrossRef]
- Ji, H.; Zhu, T.; Shan, S. Current status of dietary fish meal replacement research for non-carnivorous fish species. J. Dalian Fish. Univ. 2009, 24, 343–349. [Google Scholar]
- El-Saidy, D.M.S.D.; Gaber, M.M.A. Replacement of fish meal with a mixture of different plant protein sources in juvenile Nile tilapia, Oreochromis niloticus (L.) diets. Aquacul. Res. 2003, 34, 1119–1127. [Google Scholar] [CrossRef]
- Gaylord, T.G.; Barrows, F.T.; Teague, A.M.; Johansen, K.A.; Overturf, K.E.; Shepherd, B. Supplementation of taurine and methionine to all-plant protein diets for rainbow trout (Oncorhynchus mykiss). Aquaculture 2007, 269, 514–524. [Google Scholar] [CrossRef]
- Sakarika, M.; Delmoitié, B.; Ntagia, E.; Chatzigiannidou, I.; Gabet, X.; Ganigué, R.; Rabaey, K. Production of microbial protein from fermented grass. Chem. Eng. J. 2021, 433, 133631. [Google Scholar] [CrossRef]
- Zhu, W.; Xiao, S.; Chen, S.; Xu, Q.; Yang, Z.; Liu, J.; Wang, H.; Lan, S. Effects of fermented mulberry leaves on growth, serum antioxidant capacity, digestive enzyme activities and microbial compositions of the intestine in crucian (Carassius carassius). Aquacul. Res. 2021, 52, 6356–6366. [Google Scholar] [CrossRef]
- Tacon, A.G.; Metian, M. Global overview on the use of fish meal and fish oil in industrially compounded aquafeeds: Trends and future prospects. Aquaculture 2008, 285, 146–158. [Google Scholar] [CrossRef]
- Urán, P.A.; Gonçalves, A.A.; Taverne-Thiele, J.J.; Schrama, J.W.; Verreth, J.A.; Rombout, J.H.W.M. Soybean meal induces intestinal inflammation in common carp (Cyprinus carpio L.). Fish Shellfish Immunol. 2008, 25, 751–760. [Google Scholar] [CrossRef]
- Suárez, J.A.; Gaxiola, G.; Mendoza, R.; Cadavid, S.; Garcia, G.; Alanis, G.; Suárez, A.; Faillace, J.; Cuzon, G. Substitution of fish meal with plant protein sources and energy budget for white shrimp Litopenaeus vannamei (Boone, 1931). Aquaculture 2009, 289, 118–123. [Google Scholar] [CrossRef]
- Ding, Z.; Zhang, Y.; Ye, J.; Du, Z.; Kong, Y. An evaluation of replacing fish meal with fermented soybean meal in the diet of Macrobrachium nipponense: Growth, nonspecific immunity, and resistance to Aeromonas hydrophila. Fish Shellfish Immunol. 2015, 44, 295–301. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhou, R.-L.; Ren, Z.-Q.; Fan, Y.-W.; Hu, S.-B.; Zhuo, C.-F.; Deng, Z.-Y. Improvement of protein quality and degradation of allergen in soybean meal fermented by Neurospora crassa. LWT 2019, 101, 220–228. [Google Scholar] [CrossRef]
- Refstie, S.; Sahlström, S.; Bråthen, E.; Baeverfjord, G.; Krogedal, P. Lactic acid fermentation eliminates indigestible carbohydrates and antinutritional factors in soybean meal for Atlantic salmon (Salmo salar). Aquaculture 2005, 246, 331–345. [Google Scholar] [CrossRef]
- Wang, P.; Zhou, Q.; Feng, J.; He, J.; Lou, Y.; Zhu, J. Effect of dietary fermented soybean meal on growth, intestinal morphology and microbiota in juvenile large yellow croaker, Larimichthys crocea. Aquacul. Res. 2019, 50, 748–757. [Google Scholar] [CrossRef]
- Yan, L.; Wu, L.; Quan, Y.; Xing, X.; Lin, J.; Wang, H. Effects of replacing fish meal with fermented soybean on growth performance, feed utilization and non-specific immune indexes of Phoxinus lagowskii Dybowski. J. Northwest A F Univ. Nat. Sci. Ed. 2017, 45, 7–13. [Google Scholar]
- Rahimnejad, S.; Zhang, J.-J.; Wang, L.; Sun, Y.; Zhang, C. Evaluation of Bacillus pumillus SE5 fermented soybean meal as a fish meal replacer in spotted seabass (Lateolabrax maculatus) feed. Aquaculture 2021, 531, 735975. [Google Scholar] [CrossRef]
- Li, C.; Zhang, B.; Liu, C.; Zhou, H.; Wang, X.; Mai, K.; He, G. Effects of dietary raw or Enterococcus faecium fermented soybean meal on growth, antioxidant status, intestinal microbiota, morphology, and inflammatory responses in turbot (Scophthalmus maximus L.). Fish Shellfish Immunol. 2020, 100, 261–271. [Google Scholar] [CrossRef]
- Zhang, J.; Qi, T.; Deng, C.; Li, X. Development of phylogenetic relationship of different crucian carp (Carassius auratus) strains in China. Henan Aquacul. 2018, 3, 25–27. [Google Scholar]
- Xu, Q.; Yang, Z.; Zhu, W.; Chen, S.; Xiao, S.; Liu, J.; Wang, H.; Lan, S. Effects of fermented soybean meal instead of fish meal on growth, serum biochemical indexes and intestinal structure of Carassius auratus. Feed Ind. Mag. 2021, 42, 31–37. [Google Scholar]
- Castrillo, G.; Teixeira, P.J.P.L.; Paredes, S.H.; Law, T.F.; de Lorenzo, L.; Feltcher, M.E.; Finkel, O.M.; Breakfield, N.W.; Mieczkowski, P.; Jones, C.D.; et al. Root microbiota drive direct integration of phosphate stress and immunity. Nature 2017, 543, 513–518. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. UPARSE: Highly accurate OUT sequences from microbial applicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naïve Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed]
- DeSantis, T.Z.; Hugenholtz, P.; Larsen, N.; Rojas, M.; Brodie, E.L.; Keller, K.; Huber, T.; Dalevi, D.; Hu, P.; Andersen, G.L. Greengenes, a chimera-checked 16S rRNA gene database and workbench compatible with ARB. Appl. Environ. Microbiol. 2006, 72, 5069–5072. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013; Available online: http://www.R-project.org/ (accessed on 21 October 2021).
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral. Ecol. 2001, 26, 32–46. [Google Scholar] [CrossRef]
- Dixon, P. VEGAN, a package of R functions for community ecology. J. Veg. Sci. 2003, 14, 927–930. [Google Scholar] [CrossRef]
- Chi, C.-H.; Cho, S.-J. Improvement of bioactivity of soybean meal by solid-state fermentation with Bacillus amyloliquefaciens versus Lactobacillus spp. and Saccharomyces cerevisiae. LWT-Food Sci. Technol. 2016, 68, 619–625. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, X.; Yang, G.; Guo, L. Research on the enzyme activity of neutral protease from Bacillus subtilis L1 strain fermenting the soybean meal. Food Ferment. Sci. Technol. 2018, 54, 55–59. [Google Scholar]
- Li, Y.; Shi, F.; Ma, L.; Liu, J.; Meng, Q. Preliminary study on the relationship between activity of POD and SOD in hyphae and pathogenicity of Sclerotinia sclerotiorum on soybean. J. Heilongjiang Bayi Agricul. Univ. 2021, 33, 1–6, 14. [Google Scholar]
- Yin, H.; Huang, J.; Li, X.; Zheng, X. Effect of replacing fish meal with fermented and unfermented soybean meal on growth performance, serum antioxidant capacity and digestive enzymes of juvenile Cyprinus carpio-haematopterus. Feed Indust. 2019, 40, 46–52. [Google Scholar]
- Zhang, P.; Lin, H.; Chen, G. Research progress on the effects of oral immunoglobulin in animal health. China Feed 2019, 22, 20–23. [Google Scholar]
- Ganguly, S.; Prasad, A. Microflora in fish digestive tract plays significant role in digestion and metabolism. Rev. Fish Biol. Fish. 2012, 22, 11–16. [Google Scholar] [CrossRef]
- Ni, J.; Yu, Y.; Zhang, T.; Gao, L. Comparison of intestinal bacterial communities in grass carp, Ctenopharyngodon idellus, from two different habitats. Chin. J. Oceanol. Limnol. 2012, 30, 757–765. [Google Scholar] [CrossRef]
- Ni, J.; Yan, Q.; Yu, Y.; Zhang, T. Factors influencing the grass carp gut microbiome and its effect on metabolism. FEMS Microbiol. Ecol. 2014, 87, 704–714. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.R.; Ran, C.; Ringø, E.; Zhou, Z.G. Progress in fish gastrointestinal microbiota research. Rev. Aquacul. 2018, 10, 626–640. [Google Scholar] [CrossRef]
- Li, J.; Ni, J.; Li, J.; Wang, C.; Li, X.; Wu, S.; Zhang, T.; Yu, Y.; Yan, Q. Comparative study on gastrointestinal microbiota of eight fish species with different feeding habits. J. Appl. Microbiol. 2014, 117, 1750–1760. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.C.R.; Snowberg, L.K.; Caporaso, J.G.; Knight, R.; Bolnick, D.I. Dietary input of microbes and host genetic variation shape among-population differences in stickleback gut microbiota. ISME J. 2015, 9, 2515–2526. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | FSM0 | FSM20 | FSM40 | FSM60 | FSM80 |
|---|---|---|---|---|---|
| Peruvian fish meal | 10 | 8 | 6 | 4 | 2 |
| Soybean meal | 21 | 0 | 0 | 0 | 0 |
| Fermented soybean meal | 0 | 24 | 27 | 29 | 32 |
| Rapeseed meal | 15 | 16.5 | 17.2 | 19 | 20 |
| Rice bran | 7.6 | 7 | 5.5 | 4 | 2.5 |
| Wheat bran | 4 | 3 | 2.5 | 1.76 | 0.4 |
| Bentonite | 1.43 | 0.23 | 0 | 0 | 0.24 |
| Flour | 18 | 18 | 18 | 18 | 18 |
| Cottonseed meal | 14 | 14 | 14 | 14 | 14 |
| DDGS | 5 | 5 | 5 | 5 | 5 |
| Soybean oil | 1 | 1.1 | 1.4 | 1.6 | 1.9 |
| Ca(H2PO4)2 | 1.1 | 1.3 | 1.5 | 1.7 | 2 |
| 98% Lysine Hydrochloride | 0 | 0 | 0.03 | 0.07 | 0.09 |
| Vitamin and Mineral premix a | 1 | 1 | 1 | 1 | 1 |
| Salt | 0.3 | 0.3 | 0.3 | 0.3 | 0.3 |
| Choline chloride (50%) | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 |
| Betaine | 0.15 | 0.15 | 0.15 | 0.15 | 0.15 |
| Antioxidant | 0.02 | 0.02 | 0.02 | 0.02 | 0.02 |
| Fungicide | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 |
| Total | 100 | 100 | 100 | 100 | 100 |
| Proximate composition | |||||
| Crude protein | 32.81 | 32.38 | 32.16 | 32.81 | 31.94 |
| Crude lipid | 5 | 5.7 | 5.7 | 6 | 5 |
| Ash | 11.15 | 10.35 | 10.3 | 10.15 | 10.75 |
| Items | FSM0 | FSM20 | FSM40 | FSM60 | FSM80 |
|---|---|---|---|---|---|
| T-AOC (mM) | 0.54 ± 0.03 b | 0.68 ± 0.07 a | 0.73 ± 0.01 a | 0.69 ± 0.05 a | 0.65 ± 0.13 ab |
| POD (U/mL) | 24.33 ± 4.87 b | 28.15 ± 6.99 ab | 35.39 ± 2.91 a | 27.44 ± 0.31 ab | 26.41 ± 2.78 ab |
| T-SOD (U/mL) | 556.1 ± 38.27 | 563.06 ± 19.47 | 603.44 ± 39.85 | 598.71 ± 45.71 | 560.19 ± 85.54 |
| MDA (nmol/mL) | 5.19 ± 0.85 | 5.18 ± 0.62 | 4.41 ± 1.17 | 4.61 ± 0.33 | 5.00 ± 0.53 |
| CAT (U/mL) | 2.39 ± 0.06 | 2.50 ± 1.08 | 2.67 ± 0.70 | 2.88 ± 0.56 | 2.33 ± 0.66 |
| IgM (mg/mL) | 6.05 ± 1.19 c | 14.67 ± 0.53 ab | 15.62 ± 2.17 a | 14.49 ± 1.58 ab | 13.23 ± 1.76 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, Q.; Yang, Z.; Chen, S.; Zhu, W.; Xiao, S.; Liu, J.; Wang, H.; Lan, S. Effects of Replacing Dietary Fish Meal by Soybean Meal Co-Fermented Using Bacillus subtilis and Enterococcus faecium on Serum Antioxidant Indices and Gut Microbiota of Crucian Carp Carassius auratus. Fishes 2022, 7, 54. https://doi.org/10.3390/fishes7020054
Xu Q, Yang Z, Chen S, Zhu W, Xiao S, Liu J, Wang H, Lan S. Effects of Replacing Dietary Fish Meal by Soybean Meal Co-Fermented Using Bacillus subtilis and Enterococcus faecium on Serum Antioxidant Indices and Gut Microbiota of Crucian Carp Carassius auratus. Fishes. 2022; 7(2):54. https://doi.org/10.3390/fishes7020054
Chicago/Turabian StyleXu, Qian, Zheng Yang, Siyu Chen, Wenjuan Zhu, Siyuan Xiao, Jing Liu, Hongquan Wang, and Shile Lan. 2022. "Effects of Replacing Dietary Fish Meal by Soybean Meal Co-Fermented Using Bacillus subtilis and Enterococcus faecium on Serum Antioxidant Indices and Gut Microbiota of Crucian Carp Carassius auratus" Fishes 7, no. 2: 54. https://doi.org/10.3390/fishes7020054