
Structural Transitions of Papain-like Cysteine Proteases: Implications for Sensor Development

Abstract
:1. Introduction
2. Materials and Methods
2.1. Papain Purification
2.2. Bradford Microplate Assay for Determining Protein Concentration
2.3. Determination of Proteolytic Activity
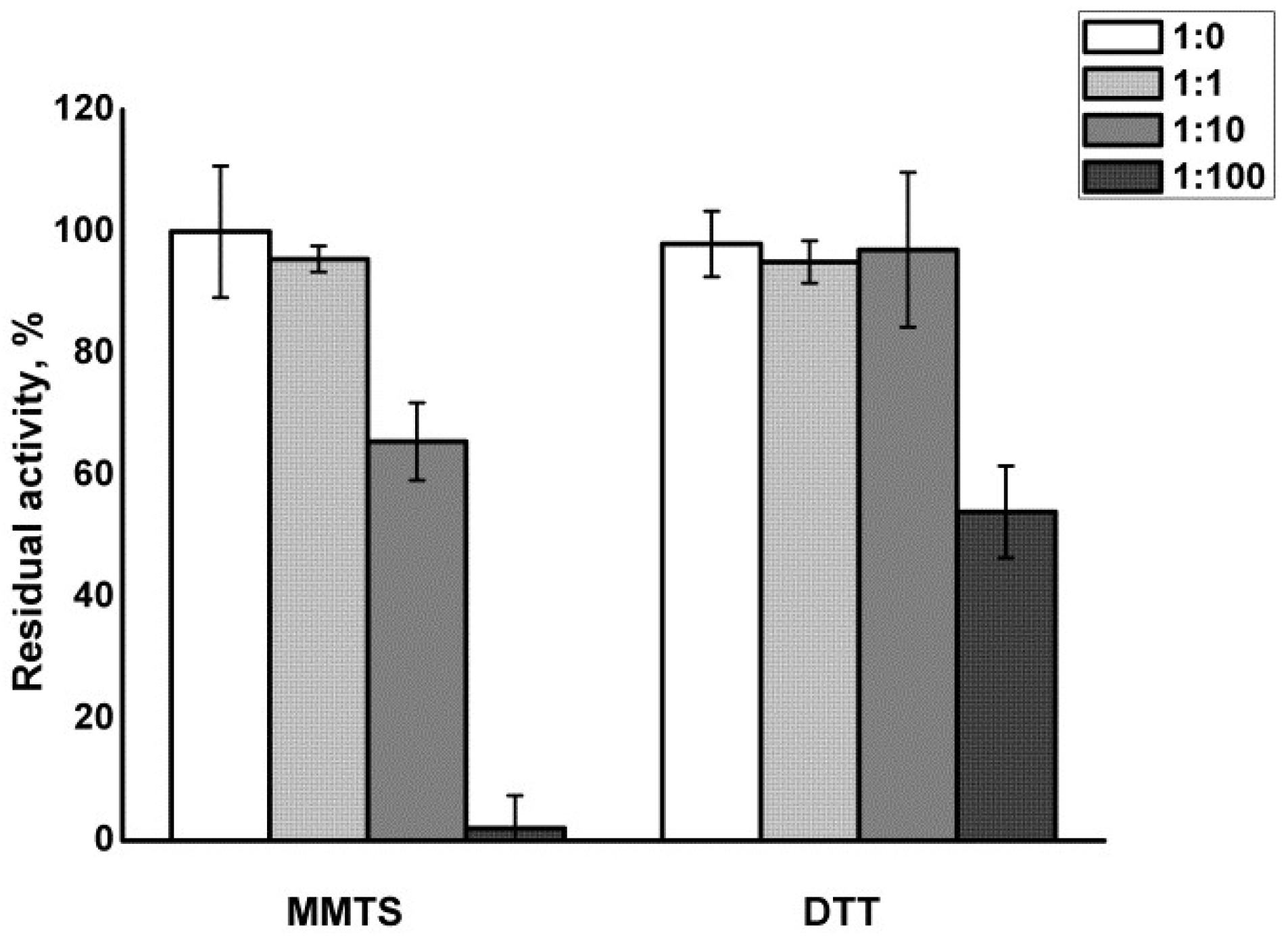
2.4. MMTS Treatment of Papain and Its Activity Recovery
2.5. Structural Characterization of MMTS-Treated Papain
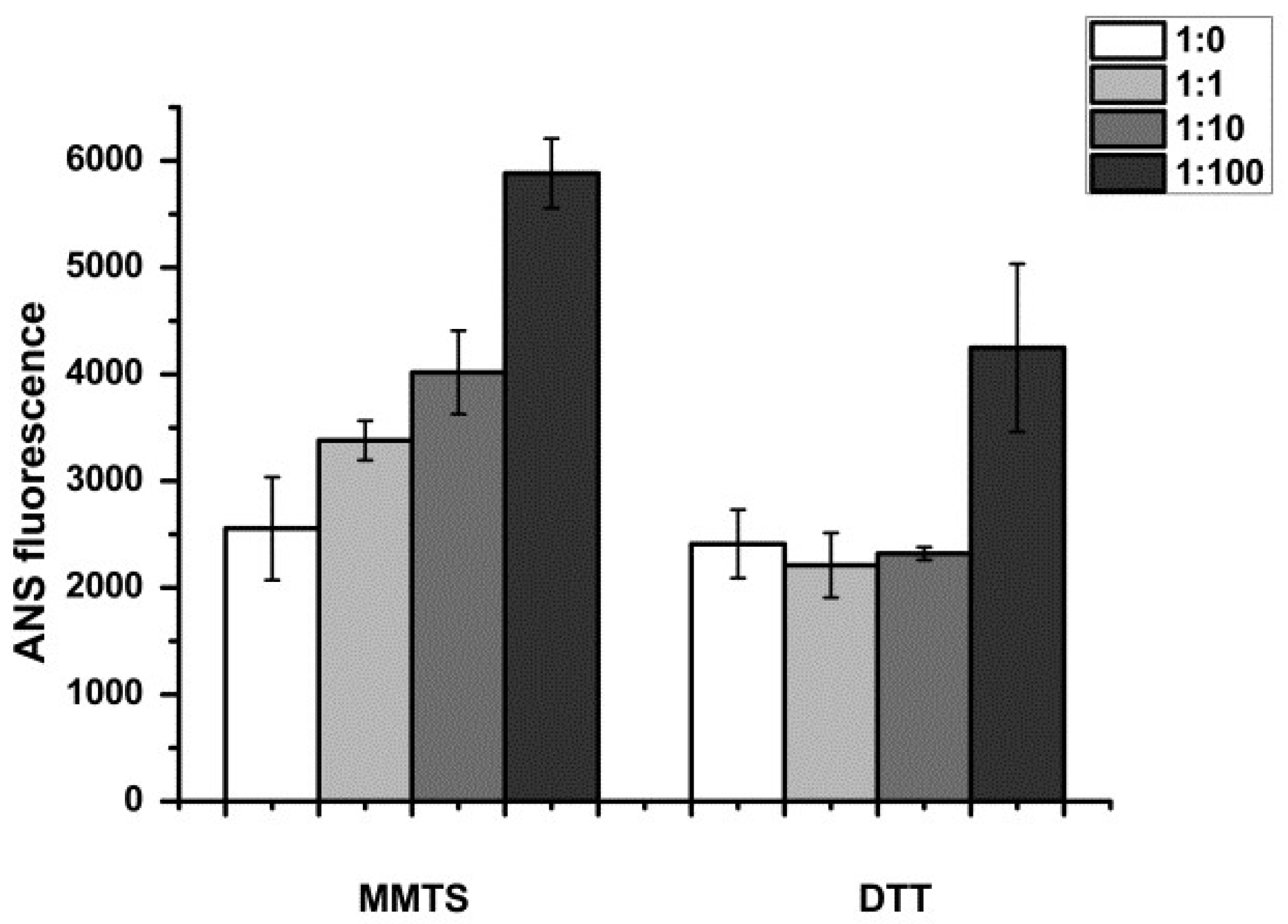
2.5.1. 8-Anilinonaphthalene-1-sulfonic Acid (ANS) Fluorescence
2.5.2. Sodium Dodecylsulfate Polyacrylamide Gel Electrophoresis (SDS-PAGE)
2.5.3. Fourier Transform Infrared Spectroscopy (FTIR) and Secondary Structures Calculation
3. Results
3.1. Papain Treatment and Recovery
3.2. Structural Transitions and Aggregation
3.2.1. Monitoring of Water-Exposed Hydrophobic Regions
3.2.2. Papain Aggregation
3.2.3. Transitions of Secondary Structures
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ANS | 8-Anilinonaphthalene-1-sulfonic acid |
| ATR | attenuated total reflectance |
| BAPNA | Nα-Benzoyl-DL-arginine p-nitroanilide hydrochloride |
| BSA | bovine serum albumin |
| DMSO | dimethyl sulfoxide |
| DTT | dithiothreitol |
| FTIR | fourier transform infrared spectroscopy |
| IR | infrared |
| MERS | Middle East respiratory syndrome |
| MMTS | S-Methyl methanethiosulfonate |
| RT | room temperature |
| SARS | severe acute respiratory syndrome |
| SDS-PAGE | sodium dodecyl sulphate–polyacrylamide gel electrophoresis |
| TCA | trichloroacetic acid |
| Tris | tris(hydroxymethyl)aminomethane |
References
- Buttle, D.J.; Mort, J.S. Cysteine Proteases. In Encyclopedia of Biological Chemistry; Elsevier: Amsterdam, The Netherlands, 2013; pp. 589–592. [Google Scholar]
- Yang, H.R.; Hwang, D.H.; Prakash, R.L.M.; Kim, J.-H.; Hong, I.-H.; Kim, S.; Kim, E.; Kang, C. Exploring the Fibrin(Ogen)Olytic, Anticoagulant, and Antithrombotic Activities of Natural Cysteine Protease (Ficin) with the κ-Carrageenan-Induced Rat Tail Thrombosis Model. Nutrients 2022, 14, 3552. [Google Scholar] [CrossRef] [PubMed]
- Balakireva, A.; Zamyatnin, A. Indispensable Role of Proteases in Plant Innate Immunity. Int. J. Mol. Sci. 2018, 19, 629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perišić Nanut, M.; Pečar Fonović, U.; Jakoš, T.; Kos, J. The Role of Cysteine Peptidases in Hematopoietic Stem Cell Differentiation and Modulation of Immune System Function. Front. Immunol. 2021, 12, 680279. [Google Scholar] [CrossRef]
- Hasanbasic, S.; Jahic, A.; Karahmet, E.; Sejranic, A.; Prnjavorac, B. The Role of Cysteine Protease in Alzheimer Disease. Mater. Socio-Med. 2016, 28, 235. [Google Scholar] [CrossRef] [Green Version]
- Behl, T.; Chadha, S.; Sehgal, A.; Singh, S.; Sharma, N.; Kaur, R.; Bhatia, S.; Al-Harrasi, A.; Chigurupati, S.; Alhowail, A.; et al. Exploring the Role of Cathepsin in Rheumatoid Arthritis. Saudi J. Biol. Sci. 2022, 29, 402–410. [Google Scholar] [CrossRef]
- Liu, C.-L.; Guo, J.; Zhang, X.; Sukhova, G.K.; Libby, P.; Shi, G.-P. Cysteine Protease Cathepsins in Cardiovascular Disease: From Basic Research to Clinical Trials. Nat. Rev. Cardiol. 2018, 15, 351–370. [Google Scholar] [CrossRef]
- Miike, S.; Kita, H. Human Eosinophils Are Activated by Cysteine Proteases and Release Inflammatory Mediators. J. Allergy Clin. Immunol. 2003, 111, 704–713. [Google Scholar] [CrossRef] [PubMed]
- Mishra, M.; Singh, V.; Singh, S. Structural Insights Into Key Plasmodium Proteases as Therapeutic Drug Targets. Front. Microbiol. 2019, 10, 394. [Google Scholar] [CrossRef] [Green Version]
- Chaimon, S.; Limpanont, Y.; Reamtong, O.; Ampawong, S.; Phuphisut, O.; Chusongsang, P.; Ruangsittichai, J.; Boonyuen, U.; Watthanakulpanich, D.; O’Donoghue, A.J.; et al. Molecular Characterization and Functional Analysis of the Schistosoma Mekongi Ca2+-Dependent Cysteine Protease (Calpain). Parasit. Vectors 2019, 12, 383. [Google Scholar] [CrossRef] [Green Version]
- Siqueira-Neto, J.L.; Debnath, A.; McCall, L.-I.; Bernatchez, J.A.; Ndao, M.; Reed, S.L.; Rosenthal, P.J. Cysteine Proteases in Protozoan Parasites. PLoS Negl. Trop. Dis. 2018, 12, e0006512. [Google Scholar] [CrossRef] [Green Version]
- Osipiuk, J.; Azizi, S.-A.; Dvorkin, S.; Endres, M.; Jedrzejczak, R.; Jones, K.A.; Kang, S.; Kathayat, R.S.; Kim, Y.; Lisnyak, V.G.; et al. Structure of Papain-like Protease from SARS-CoV-2 and Its Complexes with Non-Covalent Inhibitors. Nat. Commun. 2021, 12, 743. [Google Scholar] [CrossRef] [PubMed]
- Alaofi, A.L.; Shahid, M.; Raish, M.; Ansari, M.A.; Syed, R.; Kalam, M.A. Identification of Doxorubicin as Repurposing Inhibitory Drug for MERS-CoV PLpro. Molecules 2022, 27, 7553. [Google Scholar] [CrossRef] [PubMed]
- Adem, S.; Jain, S.; Sveiven, M.; Zhou, X.; O’Donoghue, A.J.; Hall, D.A. Giant Magnetoresistive Biosensors for Real-Time Quantitative Detection of Protease Activity. Sci. Rep. 2020, 10, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Gorodkiewicz, E. The Surface Plasmon Resonance Imaging Sensor for Papain Based on Immobilized Cystatin. Protein Pept. Lett. 2007, 14, 443–445. [Google Scholar] [CrossRef] [PubMed]
- Tokarzewicz, A.; Romanowicz, L.; Sankiewicz, A.; Hermanowicz, A.; Sobolewski, K.; Gorodkiewicz, E. A New Analytical Method for Determination of Cathepsin L Based on the Surface Plasmon Resonance Imaging Biosensor. Int. J. Mol. Sci. 2019, 20, 2166. [Google Scholar] [CrossRef] [Green Version]
- Sokolinskaya, E.L.; Putlyaeva, L.V.; Polinovskaya, V.S.; Lukyanov, K.A. Genetically Encoded Fluorescent Sensors for SARS-CoV-2 Papain-like Protease PLpro. Int. J. Mol. Sci. 2022, 23, 7826. [Google Scholar] [CrossRef]
- Yang, B.; Lv, S.; Chen, F.; Liu, C.; Cai, C.; Chen, C.; Chen, X. A Resonance Light Scattering Sensor Based on Bioinspired Molecularly Imprinted Polymers for Selective Detection of Papain at Trace Levels. Anal. Chim. Acta 2016, 912, 125–132. [Google Scholar] [CrossRef]
- Markovic, S.; Milosevic, J.; Djuric, M.; Lolic, A.; Polovic, N. One-Step Purification and Freeze Stability of Papain at Acidic PH Values. Arch. Biol. Sci. 2021, 73, 57–64. [Google Scholar] [CrossRef]
- Guo, Y.; Wang, Z.; Qu, W.; Shao, H.; Jiang, X. Colorimetric Detection of Mercury, Lead and Copper Ions Simultaneously Using Protein-Functionalized Gold Nanoparticles. Biosens. Bioelectron. 2011, 26, 4064–4069. [Google Scholar] [CrossRef]
- Lai, C.; Qin, L.; Zeng, G.; Liu, Y.; Huang, D.; Zhang, C.; Xu, P.; Cheng, M.; Qin, X.; Wang, M. Sensitive and Selective Detection of Mercury Ions Based on Papain and 2,6-Pyridinedicarboxylic Acid Functionalized Gold Nanoparticles. RSC Adv. 2016, 6, 3259–3266. [Google Scholar] [CrossRef]
- Xiang, X.; Shi, J.; Guo, M.; Jin, H.; Huang, F. Colorimetric Sensing of Sinapine Based on Competitive Interaction of Papain-Coated Gold Nanoparticles and Sinapine toward Pb2+. RSC Adv. 2014, 4, 42905–42909. [Google Scholar] [CrossRef]
- Athira, C.R.; Shilpa, S.; Parvathy, C.M.; Nithya, S.; Vinothaa, V.; Subashini, S.P.; Varun Gopal, K.; Parimalam, B.; Namboori, P.K.K. Computational Characterization of CNT-Papain Interactions for Developing a Biosensor. In Proceedings of the Second International Conference on Advances in Communication, Network, and Computing, Bangalore, India, 10–11 March 2011. [Google Scholar]
- Kozlowski, R.; Ragupathi, A.; Dyer, R.B. Characterizing the Surface Coverage of Protein–Gold Nanoparticle Bioconjugates. Bioconjug. Chem. 2018, 29, 2691–2700. [Google Scholar] [CrossRef]
- Sahoo, B.; Sahu, S.K.; Bhattacharya, D.; Dhara, D.; Pramanik, P. A Novel Approach for Efficient Immobilization and Stabilization of Papain on Magnetic Gold Nanocomposites. Colloids Surf. B Biointerfaces 2013, 101, 280–289. [Google Scholar] [CrossRef]
- Brito, A.M.M.; Oliveira, V.; Icimoto, M.Y.; Nantes-Cardoso, I.L. Collagenase Activity of Bromelain Immobilized at Gold Nanoparticle Interfaces for Therapeutic Applications. Pharmaceutics 2021, 13, 1143. [Google Scholar] [CrossRef]
- Karala, A.; Ruddock, L.W. Does S—Methyl Methanethiosulfonate Trap the Thiol–Disulfide State of Proteins? Antioxid. Redox Signal. 2007, 9, 527–531. [Google Scholar] [CrossRef]
- Bradford, M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Vatic, S.; Mirkovic, N.; Milosevic, J.; Jovcic, B.; Polovic, N. Trypsin activity and freeze-thaw stability in the presence of ions and non-ionic surfactants. J. Biosci. Bioeng. 2021, 131, 234–240. [Google Scholar] [CrossRef]
- Milošević, J.; Prodanović, R.; Polović, N. On the Protein Fibrillation Pathway: Oligomer Intermediates Detection Using ATR-FTIR Spectroscopy. Molecules 2021, 26, 970. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Milošević, J.; Petrić, J.; Jovčić, B.; Janković, B.; Polović, N. Exploring the Potential of Infrared Spectroscopy in Qualitative and Quantitative Monitoring of Ovalbumin Amyloid Fibrillation. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2020, 229, 117882. [Google Scholar] [CrossRef]
- Rašković, B.; Popović, M.; Ostojić, S.; Anđelković, B.; Tešević, V.; Polović, N. Fourier Transform Infrared Spectroscopy Provides an Evidence of Papain Denaturation and Aggregation during Cold Storage. Spectrochim. Acta A. Mol. Biomol. Spectrosc. 2015, 150, 238–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vedantham, G.; Sparks, H.G.; Sane, S.U.; Tzannis, S.; Przybycien, T.M. A Holistic Approach for Protein Secondary Structure Estimation from Infrared Spectra in H2O Solutions. Anal. Biochem. 2000, 285, 33–49. [Google Scholar] [CrossRef]
- Goossens, K.; Haelewyn, J.; Meersman, F.; de Ley, M.; Heremans, K. Pressure- and Temperature-Induced Unfolding and Aggregation of Recombinant Human Interferon-Gamma: A Fourier Transform Infrared Spectroscopy Study. Biochem. J. 2003, 370, 529–535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milošević, J.; Janković, B.; Prodanović, R.; Polović, N. Comparative Stability of Ficin and Papain in Acidic Conditions and the Presence of Ethanol. Amino Acids 2019, 51, 829–838. [Google Scholar] [CrossRef] [PubMed]
- Hermanson, G.T. Functional Targets for Bioconjugation. In Bioconjugate Techniques, 3rd ed.; Elsevier: Amsterdam, The Netherlands, 2013; pp. 127–228. [Google Scholar]
- Makarov, V.A.; Tikhomirova, N.K.; Savvateeva, L.; Petushkova, A.I.; Serebryakova, M.; Baksheeva, V.E.; Gorokhovets, N.; Zernii, E.Y.; Zamyatnin, A.A. Novel Applications of Modification of Thiol Enzymes and Redox-Regulated Proteins Using S-Methyl Methanethiosulfonate (MMTS). Biochim. Biophys. Acta Proteins Proteom. 2019, 1867, 140259. [Google Scholar] [CrossRef]
- Rodríguez-Romero, A.; Hernández-Santoyo, A.; del Pozo Yauner, L.; Kornhauser, A.; Fernández-Velasco, D.A. Structure and Inactivation of Triosephosphate Isomerase from Entamoeba Histolytica. J. Mol. Biol. 2002, 322, 669–675. [Google Scholar] [CrossRef]
- Brewer, S.H.; Glomm, W.R.; Johnson, M.C.; Knag, M.K.; Franzen, S. Probing BSA Binding to Citrate-Coated Gold Nanoparticles and Surfaces. Langmuir 2005, 21, 9303–9307. [Google Scholar] [CrossRef]
- Mohan, V.; Das, N.; Das, A.; Mishra, V.; Sen, P. Spectroscopic Insight on Ethanol-Induced Aggregation of Papain. J. Phys. Chem. B 2019, 123, 2280–2290. [Google Scholar] [CrossRef]
- Raskovic, B.; Babic, N.; Korac, J.; Polovic, N. Evidence of β-Sheet Structure Induced Kinetic Stability of Papain upon Thermal and Sodium Dodecyl Sulphate Denaturation. J. Serb. Chem. Soc. 2015, 80, 613–625. [Google Scholar] [CrossRef]
- Milošević, J.; Vrhovac, L.; Đurković, F.; Janković, B.; Malkov, S.; Lah, J.; Polović, N.Đ. Isolation, Identification, and Stability of Ficin 1c Isoform from Fig Latex. New J. Chem. 2020, 44, 15716–15723. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein:MMTS Ratio | β-Sheet | Unordered | α-Helix | Turn | Error |
|---|---|---|---|---|---|
| MMTS 1:0 | 25.5 | 43.0 | 24.9 | 6.6 | 1.5 |
| MMTS 1:1 | 27.9 | 39.5 | 26.6 | 6.0 | 1.4 |
| MMTS 1:10 | 37.8 | 33.9 | 20.9 | 7.4 | 1.0 |
| MMTS 1:100 | 52.0 | 28.2 | 14.6 | 7.2 | 1.8 |
| DTT-MMTS 1:0 | 24.2 | 44.2 | 25.5 | 6.1 | 0.5 |
| DTT-MMTS 1:1 | 25.2 | 41.9 | 26.0 | 6.9 | 1.0 |
| DTT-MMTS 1:10 | 26.8 | 40.2 | 26.9 | 6.1 | 0.9 |
| DTT-MMTS 1:100 | 30.1 | 38.6 | 24.9 | 6.4 | 1.6 |
| X-ray | 25 | 42 | 26 | 7 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marković, S.; Andrejević, N.S.; Milošević, J.; Polović, N.Đ. Structural Transitions of Papain-like Cysteine Proteases: Implications for Sensor Development. Biomimetics 2023, 8, 281. https://doi.org/10.3390/biomimetics8030281
Marković S, Andrejević NS, Milošević J, Polović NĐ. Structural Transitions of Papain-like Cysteine Proteases: Implications for Sensor Development. Biomimetics. 2023; 8(3):281. https://doi.org/10.3390/biomimetics8030281
Chicago/Turabian StyleMarković, Srdjan, Natalija S. Andrejević, Jelica Milošević, and Natalija Đ. Polović. 2023. "Structural Transitions of Papain-like Cysteine Proteases: Implications for Sensor Development" Biomimetics 8, no. 3: 281. https://doi.org/10.3390/biomimetics8030281