
Enhancing Kidney Vasculature in Tissue Engineering—Current Trends and Approaches: A Review

Abstract
:1. Introduction
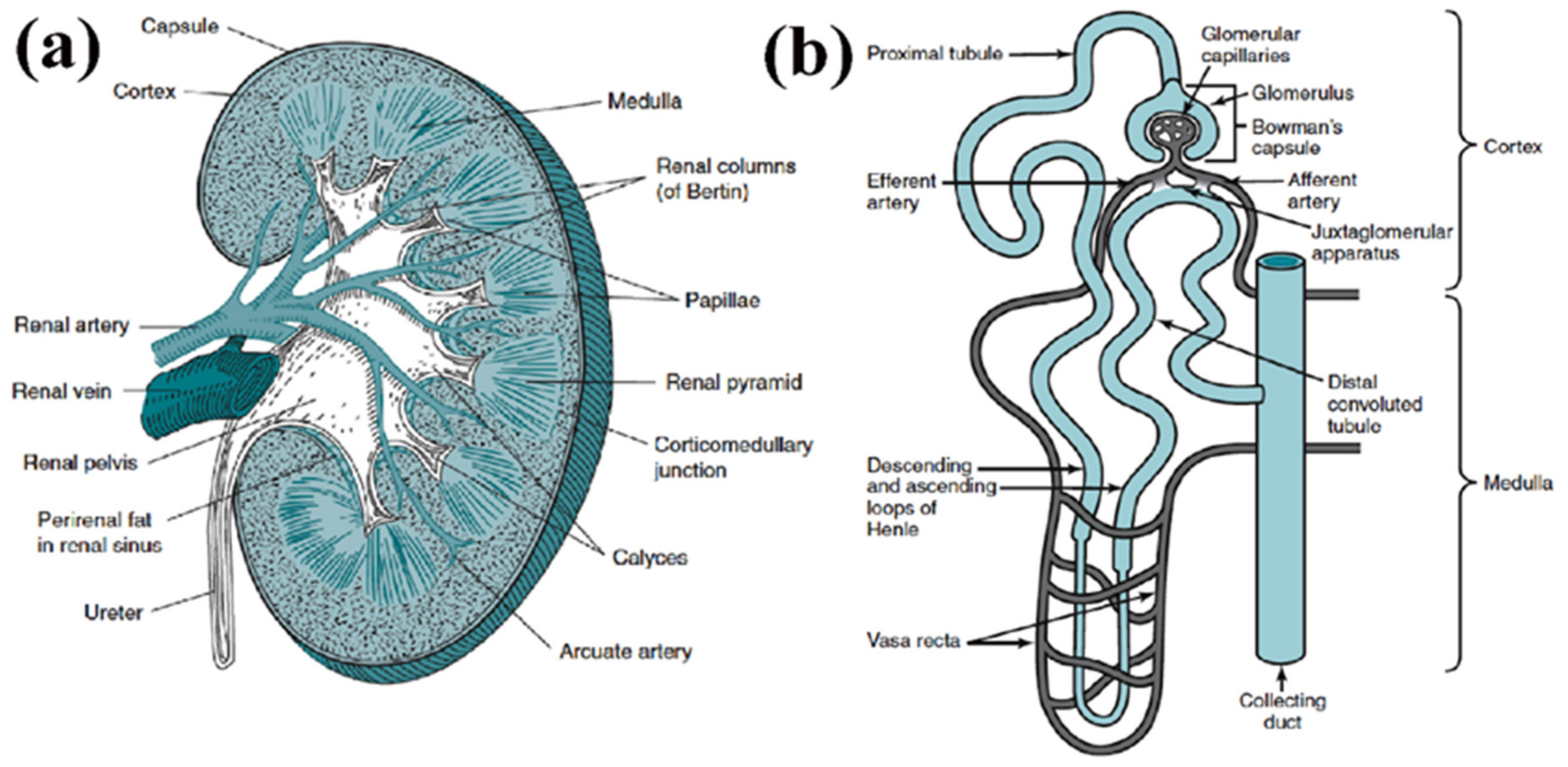
2. Renal Vascular Structure and Cellular Composition
3. Whole-Kidney Tissue Engineering—The Top-Down Approach
3.1. Decellularized ECM
3.2. Vascular Corrosion Casts
4. Kidney Organoids—The Bottom-Up Approach
Microphysiological Systems
5. Conclusions and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Giwa, S.; Lewis, J.K.; Alvarez, L.; Langer, R.; Roth, E.A.; Church, G.M.; Markmann, J.F.; Sachs, D.H.; Chandraker, A.; Wertheim, A.J.; et al. The promise of organ and tissue preservation to transform medicine. Nat. Biotechnol. 2017, 35, 530–542. [Google Scholar] [CrossRef] [PubMed]
- Lo, D.J.; Kaplan, B.; Kirk, A.D. Biomarkers for kidney transplant rejection. Nat. Rev. Nephrol. 2014, 10, 215–225. [Google Scholar] [CrossRef] [PubMed]
- Dzobo, K.; Thomford, N.E.; Senthebane, D.A.; Shipanga, H.; Rowe, A.; Dandara, C.; Pillay, M.; Motaung, K.S.C.M. Advances in Regenerative Medicine and Tissue Engineering: Innovation and Transformation of Medicine. STEM Cells Int. 2018, 2018, 1–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Sherbiny, I.M.; Yacoub, M.H. Hydrogel scaffolds for tissue engineering: Progress and challenges. Glob. Cardiol. Sci. Pract. 2013, 2013, 316–342. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Yang, J.; Wan, Y.; Bei, J. Research progress on biodegradable materials in tissue engineering. Chem. Bull./Huaxue Tongbao 2004, 67, 237–245. [Google Scholar]
- Khatoon, Z.; McTiernan, C.D.; Suuronen, E.J.; Mah, T.-F.; Alarcon, E.I. Bacterial biofilm formation on implantable devices and approaches to its treatment and prevention. Heliyon 2018, 4, e01067. [Google Scholar] [CrossRef] [Green Version]
- Kular, J.K.; Basu, S.; I Sharma, R. The extracellular matrix: Structure, composition, age-related differences, tools for analysis and applications for tissue engineering. J. Tissue Eng. 2014, 5, 041731414557112. [Google Scholar] [CrossRef] [PubMed]
- Rademakers, T.; Horvath, J.M.; Blitterswijk, C.A.; LaPointe, V.L.S. Oxygen and nutrient delivery in tissue engi-neering: Approaches to graft vascularization. J. Tissue Eng. Regen. Med. 2019, 10, 1815–1829. [Google Scholar] [CrossRef]
- Betz, C.; Lenard, A.; Belting, H.-G.; Affolter, M. Cell behaviors and dynamics during angiogenesis. Development 2016, 143, 2249–2260. [Google Scholar] [CrossRef] [Green Version]
- Marsano, A.; Medeiros da Cunha, C.M.; Ghanaati, S.; Gueven, S.; Centola, M.; Tsaryk, R.; Barbeck, M.; Stuedle, C.; Barbero, A.; Helmrich, U.; et al. Spontaneous in vivo Chondro-genesis of bone marrow-derived Mesenchymal progenitor cells by blocking vascular endothelial growth factor signaling. Stem Cells Transl. Med. 2016, 5, 1730–1738. [Google Scholar] [CrossRef] [Green Version]
- Kumar, V.V.; Brewster, L.P.; Caves, J.M.; Chaikof, E.L. Tissue engineering of blood vessels: Functional require-ments, progress and future challenges. Cariovasc. Eng. Technol. 2011, 2, 137–148. [Google Scholar] [CrossRef] [Green Version]
- Fu, J.; Wang, D.-A. In Situ Organ-Specific Vascularization in Tissue Engineering. Trends Biotechnol. 2018, 36, 834–849. [Google Scholar] [CrossRef]
- Auger, F.A.; Gibot, L.; Lacroix, D. The Pivotal Role of Vascularization in Tissue Engineering. Annu. Rev. Biomed. Eng. 2013, 15, 177–200. [Google Scholar] [CrossRef]
- A Kaballo, M.; Canney, M.; O’Kelly, P.; Williams, Y.; O’Seaghdha, C.M.; Conlon, P.J. A comparative analysis of survival of patients on dialysis and after kidney transplantation. Clin. Kidney J. 2018, 11, 389–393. [Google Scholar] [CrossRef]
- Gupta, A.K.; Sarkar, P.; Wertheim, J.A.; Pan, X.; Carroll, T.J.; Oxburgh, L. Asynchronous mixing of kidney progenitor cells potentiates nephrogenesis in organoids. Commun. Biol. 2020, 3, 1–11. [Google Scholar] [CrossRef]
- Batchelder, C.A.; Martinez, M.L.; Tarantal, A.F. Natural Scaffolds for Renal Differentiation of Human Embryonic Stem Cells for Kidney Tissue Engineering. PLoS ONE 2015, 10, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Takasato, M.; Wymeersch, F.J. Challenges to future regenerative applications using kidney organoids. Curr. Opin. Biomed. Eng. 2020, 13, 144–151. [Google Scholar] [CrossRef]
- Wragg, N.; Burke, L.; Wilson, S.L. A critical review of current progress in 3D kidney biomanufacturing: Advances, challenges, and recommendations. Ren. Replace. Ther. 2019, 5, 18. [Google Scholar] [CrossRef] [Green Version]
- Bülow, R.D.; Boor, P. Extracellular Matrix in Kidney Fibrosis: More Than Just a Scaffold. J. Histochem. Cytochem. 2019, 67, 643–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagao, R.J.; Xu, J.; Luo, P.; Xue, J.; Wang, Y.; Kotha, S.; Zeng, W.; Fu, X.; Himmelfarb, J.; Zheng, Y. Decellularized Human Kidney Cortex Hydrogels Enhance Kidney Microvascular Endothelial Cell Maturation and Quiescence. Tissue Eng. Part A 2016, 22, 1140–1150. [Google Scholar] [CrossRef] [Green Version]
- Schmidt-Ott, K.M. How to grow a kidney: Patient-specific kidney organoids come of age. Nephrol. Dial. Transplant. 2016, 32, 17–23. [Google Scholar] [CrossRef]
- Nishinakamura, R. Human kidney organoids: Progress and remaining challenges. Nat. Rev. Nephrol. 2019, 15, 613–624. [Google Scholar] [CrossRef]
- Mohamed, T.; Sequeira-Lopez, M.L.S. Development of the renal vasculature. Semin. Cell. Dev. Biol. 2018, 91, 132–146. [Google Scholar] [CrossRef]
- Shaw, I.; Rider, S.; Mullins, J.; Hughes, J.; Péault, B. Pericytes in the renal vasculature: Roles in health and disease. Nature Rev. Nephrol. 2018, 14, 521–534. [Google Scholar] [CrossRef]
- Moorthy, V.; Blichfeldt, T.C. Anatomy and Physiology of the Kidney. In Pathophysiology of Kidney Disease and Hypertension; Elsevier: Amsterdam, The Netherlands, 2009; Chapter 1; pp. 1–15. [Google Scholar]
- Berg, C.W.V.D.; Ritsma, L.; Avramut, M.C.; Wiersma, L.E.; Berg, B.M.V.D.; Leuning, D.G.; Lievers, E.; Koning, M.; Vanslambrouck, J.M.; Koster, A.J.; et al. Renal Subcapsular Transplantation of PSC-Derived Kidney Organoids Induces Neo-vasculogenesis and Significant Glomerular and Tubular Maturation In Vivo. Stem Cell Rep. 2018, 10, 751–765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aird, W.C. Endothelial cell heterogeneity. Cold Spring Harb. Perspect. Med. 2012, 2, a006429. [Google Scholar] [CrossRef] [PubMed]
- Uwamori, H.; Ono, Y.; Yamashita, T.; Arai, K.; Sudo, R. Comparison of organ-specific endothelial cells in terms of microvascular formation and endothelial barrier functions. Microvasc. Res. 2019, 122, 60–70. [Google Scholar] [CrossRef] [PubMed]
- Wimmer, R.A.; Leopoldi, A.; Aichinger, M.; Wick, N.; Hantusch, B.; Novatchkova, M.; Taubenschmid, J.; Hämmerle, M.; Esk, C.; Bagley, J.A.; et al. Human blood vessel organoids as a model of diabetic vasculopathy. Nature 2019, 565, 505–510. [Google Scholar] [CrossRef] [PubMed]
- Shafiee, A.; Patel, J.; Lee, J.S.; Hutmacher, D.W.; Fisk, N.M.; Khosrotehrani, K. Mesenchymal stem/stromal cells enhance engraftment, vasculogenic and pro-angiogenic activities of endothelial colony forming cells in immunocompetent hosts. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef]
- Moore, E.M.; Suresh, V.; Ying, G.; West, J.L. M0 and M2 Macrophages Enhance Vascularization of Tissue Engineering Scaffolds. Regen. Eng. Transl. Med. 2018, 4, 51–61. [Google Scholar] [CrossRef]
- Chong, M.; Ng, W.K.; Chan, J.K.Y. Concise Review: Endothelial Progenitor Cells in Regenerative Medicine: Applications and Challenges. Stem Cells Transl. Med. 2016, 5, 530–538. [Google Scholar] [CrossRef] [Green Version]
- Rich, M.H.; Lee, M.K.; Baek, K.; Jeong, J.H.; Kim, D.H.; Millet, L.J.; Bashir, R.; Kong, H. Material-mediated proangiogenic factor release pattern modulates quality of regenerated blood vessels. J. Control. Release 2014, 196, 363–369. [Google Scholar] [CrossRef]
- Akar, B.; Jiang, B.; Somo, S.I.; Appel, A.A.; Larson, J.C.; Tichauer, K.M.; Brey, E.M. Biomaterials with persistent growth factor gradients in vivo accelerate vascularized tissue formation. Biomaterials 2015, 72, 61–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Figliuzzi, M.; Bonandrini, B.; Remuzzi, A. Decellularized Kidney Matrix as Functional Material for whole Organ Tissue Engineering. J. Appl. Biomater. Funct. Mater. 2017, 15, e326–e333. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.-Q.; Wang, L.-R.; Pan, L.-L.; Wang, H.; Zhu, G.-Q.; Liu, W.-Y.; Wang, J.-T.; Braddock, M.; Zheng, M.-H. Kidney bioengineering in regenerative medicine: An emerging therapy for kidney disease. Cytotherapy 2016, 18, 186–197. [Google Scholar] [CrossRef]
- Salvatori, M.; Peloso, A.; Katari, R.; Orlando, G. Regeneration and Bioengineering of the Kidney: Current Status and Future Challenges. Curr. Urol. Rep. 2014, 15, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Peloso, A.; Petrosyan, A.; Da Sacco, S.; Booth, C.; Zambon, J.P.; O’Brien, T.; Aardema, C.; Robertson, J.; E De Filippo, R.; Soker, S.; et al. Renal Extracellular Matrix Scaffolds from Discarded Kidneys Maintain Glomerular Morphometry and Vascular Resilience and Retains Critical Growth Factors. Transplantation 2015, 99, 1807–1816. [Google Scholar] [CrossRef] [PubMed]
- Song, J.J.; Guyette, J.P.; Gilpin, S.E.; Gonzalez, G.; Vacanti, J.P.; Ott, H.C. Regeneration and experimental ortho-topic transplantation of a bioengineered kidney. Nat. Med. 2013, 19, 646–651. [Google Scholar] [CrossRef]
- Hussein, K.H.; Saleh, T.; Ahmed, E.; Kwak, H.H.; Park, K.M.; Yang, S.R.; Kang, B.J.; Choi, K.Y.; Kang, K.S.; Woo, H.M. Biocompatibility and hemocompatibility of efficiently decellularized whole porcine kidney for tissue en-gineering. J. Biomed. Mater. Res. A 2018, 7, 2034–2047. [Google Scholar] [CrossRef]
- Zambon, J.P.; Ko, I.K.; Abolbashari, M.; Huling, J.; Clouse, C.; Kim, T.H.; Smith, C.; Atala, A.; Yoo, J.J. Compara-tive analysis of two porcine kidney decellularization methods for maintenance of functional vascular architec-tures. Acta Biomater. 2018, 75, 226–234. [Google Scholar] [CrossRef]
- Ross, E.A.; Abrahamson, D.R.; St John, P.; Clapp, W.L.; Williams, M.J.; Terada, N.; Hamazaki, T.; Ellison, G.W.; Batich, C.D. Mouse stem cells seeded into decellularized rat kidney scaffolds endothelialize and remodel basement membranes. Organogenesis 2012, 8, 49–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciampi, O.; Bonandrini, B.; Derosas, M.; Conti, S.; Rizzo, P.; Benedetti, V.; Figliuzzi, M.; Remuzzi, A.; Benigni, A.; Remuzzi, G.; et al. Engineering the vasculature of decellularized rat kidney scaffolds using human induced pluripotent stem cell-derived endothelial cells. Sci. Rep. 2019, 9, 8001. [Google Scholar] [CrossRef]
- Leuning, D.G.; Witjas, F.M.R.; Maanaoui, M.; De Graaf, A.M.A.; Lievers, E.; Geuens, T.; Avramut, C.M.; Wiersma, L.E.; Berg, C.W.V.D.; Sol, W.M.P.J.; et al. Vascular bioengineering of scaffolds derived from human discarded transplant kidneys using human pluripotent stem cell–derived endothelium. Arab. Archaeol. Epigr. 2018, 19, 1328–1343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y.; Liu, L.; Zhang, J.; Wei, Z.; Mei, J. Glutaraldehyde Cross-linking Modification of Decellularized Rat Kidney Scaffolds. Adv. Struct. Saf. Stud. 2017, 1577, 111–119. [Google Scholar] [CrossRef]
- Wang, M.; Bao, L.; Qiu, X.; Yang, X.; Liu, S.; Su, Y.; Wang, L.; Liu, B.; He, Q.; Liu, S.; et al. Immobilization of hep-arin on decellularized kidney scaffold to construct microenvironment for antithrombosis and inducing reendo-thelialization. Sci. China Life Sci. 2018, 61, 1168–1177. [Google Scholar] [CrossRef] [PubMed]
- Tarantal, A.F.; Batchelder, C.A. Scaffolds for kidney tissue engineering. In Handbook of Tissue Engineering Scaffolds; Woodhead Publishing: Sawston, UK, 2019; Volume 2, pp. 477–492. [Google Scholar] [CrossRef]
- Lih, E.; Park, W.; Park, K.W.; Chun, S.Y.; Kim, H.; Joung, Y.K.; Kwon, T.G.; Hubbell, J.A.; Han, D.K. A Bioinspired Scaffold with Anti-Inflammatory Magnesium Hydroxide and Decellularized Extracellular Matrix for Renal Tissue Regeneration. ACS Cent. Sci. 2019, 5, 458–467. [Google Scholar] [CrossRef]
- Su, J.; Satchell, S.C.; Shah, R.N.; Wertheim, J.A. Kidney decellularized extracellular matrix hydrogels: Rheological characterization and human glomerular endothelial cell response to encapsulation. J. Biomed. Mater. Res. A 2018, 9, 2448. [Google Scholar] [CrossRef]
- Hiraki, H.L.; Nagao, R.J.; Himmelfarb, J.; Zheng, Y. Fabricating a Kidney Cortex Extracellular Matrix-Derived Hydrogel. J. Vis. Exp. 2018, e58314. [Google Scholar] [CrossRef]
- Yu, Y.; Cui, H.; Wen, G.; Xu, J.; Wang, C.; Chai, Y.; Zheng, B.; Zhang, J.; Mei, J.; Chen, C. Hypoxia-inducible Factor-1α directs renal regeneration induced by decellularized scaffolds. Biomaterials 2018, 165, 48–55. [Google Scholar] [CrossRef]
- Huling, J.; Ko, I.K.; Atala, A.; Yoo, J.J. Fabrication of biomimetic vascular scaffolds for 3D tissue constructs using vascular corrosion casts. Acta Biomater. 2016, 32, 190–197. [Google Scholar] [CrossRef]
- Huling, J.; Min, S.-I.; Kim, D.S.; Ko, I.K.; Atala, A.; Yoo, J.J. Kidney regeneration with biomimetic vascular scaffolds based on vascular corrosion casts. Acta Biomater. 2019, 95, 328–336. [Google Scholar] [CrossRef] [PubMed]
- Little, M.H.; Hale, L.J.; Howden, S.E.; Kumar, S.V. Generating Kidney from Stem Cells. Annu. Rev. Physiol. 2019, 81, 335–357. [Google Scholar] [CrossRef]
- Tajiri, S.; Yamanaka, S.; Fujimoto, T.; Matsumoto, K.; Taguchi, A.; Nishinakamura, R.; Okano, H.J.; Yokoo, T. Regenerative potential of induced pluripotent stem cells derived from patients undergoing haemodialysis in kidney regeneration. Sci. Rep. 2018, 8, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Combes, A.N.; Zappia, L.; Er, P.X.; Oshlack, A.; Little, M.H. Single-cell analysis reveals congruence between kidney organoids and human fetal kidney. Genome Med. 2019, 11, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chuah, J.K.C.; Zink, D. Stem cell-derived kidney cells and organoids: Recent breakthroughs and emerging applications. Biotechnol. Adv. 2017, 35, 150–167. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.; Nishinakamura, R. How to rebuild the kidney: Recent advances in kidney organoids. J. Biochem. 2019, 166, 7–12. [Google Scholar] [CrossRef]
- Morizane, R.; Lam, A.Q.; Freedman, B.S.; Kishi, S.; Valerius, M.T.; Bonventre, J.V. Nephron organoids derived from human pluripotent stem cells model kidney development and injury. Nat. Biotechnol. 2015, 33, 1193–1200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharmin, S.; Taguchi, A.; Kaku, Y.; Yoshimura, Y.; Ohmori, T.; Sakuma, T.; Mukoyama, M.; Yamamoto, T.; Ku-rihara, H.; Nishinakamura, R. Human Induced Pluripotent Stem Cell-Derived Podocytes Mature into Vascular-ized Glomeruli upon Experimental Transplantation. J. Am. Soc. Nephrol. 2016, 27, 1778–1791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morizane, R.; Bonventre, J.V. Generation of nephron progenitor cells and kidney organoids from human plu-ripotent stem cells. Nat. Protoc. 2017, 1, 195–207. [Google Scholar] [CrossRef] [PubMed]
- Low, J.H.; Li, P.; Chew, E.G.Y.; Zhou, B.; Suzuki, K.; Zhang, T.; Lian, M.M.; Liu, M.; Aizawa, E.; Esteban, C.R.; et al. Generation of Human PSC-Derived Kidney Organoids with Patterned Nephron Segments and a De Novo Vascular Network. Cell Stem Cell 2019, 25, 373–387.e9. [Google Scholar] [CrossRef]
- Takasato, M.; Er, P.X.; Becroft, M.; Vanslambrouck, J.M.; Stanley, E.G.; Elefanty, A.G.; Little, M.H. Directing hu-man embryonic stem cell differentiation towards a renal lineage generates a self-organizing kidney. Nat. Cell Biol. 2014, 16, 118–126. [Google Scholar] [CrossRef]
- Takasato, M.; Er, P.X.; Chiu, H.S.; Maier, B.; Baillie, G.J.; Ferguson, C.; Parton, R.G.; Wolvetang, E.J.; Roost, M.S.; de Sousa Lopes, S.M.C.; et al. Kidney organoids from human iPS cells contain multiple lineages and mod-el human nephrogenesis. Nature 2015, 526, 564–568. [Google Scholar] [CrossRef]
- Howden, S.E.; Little, M.H. Generating Kidney Organoids from Human Pluripotent Stem Cells Using Defined Conditions. Methods Mol. Biol. 2020, 2155, 183–192. [Google Scholar] [CrossRef] [PubMed]
- Przepiorski, A.; Sander, V.; Tran, T.; Hollywood, J.A.; Sorrenson, B.; Shih, J.-H.; Wolvetang, E.J.; McMahon, A.P.; Holm, T.M.; Davidson, A.J. A Simple Bioreactor-Based Method to Generate Kidney Organoids from Pluripotent Stem Cells. Stem Cell Rep. 2018, 11, 470–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Czerniecki, S.M.; Cruz, N.M.; Harder, J.L.; Menon, R.; Annis, J.; Otto, E.A.; Gulieva, R.E.; Islas, L.V.; Kim, Y.K.; Tran, L.M.; et al. High-Throughput Screening Enhances Kidney Organoid Differentiation from Human Plu-ripotent Stem Cells and Enables Automated Multidimensional Phenotyping. Cell Stem Cell 2018, 22, 929–940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.V.; Er, P.X.; Lawlor, K.; Motazedian, A.; Scurr, M.; Ghobrial, I.; Combes, A.N.; Zappia, L.; Oshlack, A.; Stanley, E.G.; et al. Kidney micro-organoids in suspension culture as a scalable source of human pluripotent stem cell-derived kidney cells. Development 2019, 146, 172361. [Google Scholar] [CrossRef] [Green Version]
- Taguchi, A.; Nishinakamura, R. Higher-Order Kidney Organogenesis from Pluripotent Stem Cells. Cell Stem Cell 2017, 21, 730–746.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, H.; Uchimura, K.; Donnelly, E.L.; Kirita, Y.; Morris, S.A.; Humphreys, B.D. Comparative Analysis and Re-finement of Human PSC-Derived Kidney Organoid Differentiation with Single-Cell Transcriptomics. Cell Stem Cell 2018, 23, 869–881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geuens, T.; Van Blitterswijk, C.A.; Lapointe, V.L.S. Overcoming kidney organoid challenges for regenerative medicine. NPJ Regen. Med. 2020, 5, 1–6. [Google Scholar] [CrossRef]
- Basu, J.; Bertram, T.A.; Ludlow, J.W. Regenerating Kidney Structure and Function: An Industry Perspective. In Kidney Transplantation, Bioengineering, and Regeneration; Academic Press: Cambridge, MA, USA, 2017; Chapter 87; pp. 1177–1187. [Google Scholar]
- Rosines, E.; Sampogna, R.V.; Johkura, K.; Vaughn, D.A.; Choi, Y.; Sakurai, H.; Shah, M.M.; Nigam, S.K. Staged in vitro Reconstitution and Implantation of Engineered Rat Kidney Tissue. Proc. Natl. Acad. Sci. USA 2007, 104, 20938–20943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taguchi, A.; Kaku, Y.; Ohmori, T.; Sharmin, S.; Ogawa, M.; Sasaki, H.; Nishinakamura, R. Redefining the In Vivo Origin of Metanephric Nephron Progenitors Enables Generation of Complex Kidney Structures from Pluripo-tent Stem Cells. Cell Stem Cell 2014, 14, 53–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bantounas, I.; Ranjzad, P.; Tengku, F.; Silajdžić, E.; Forster, D.; Asselin, M.-C.; Lewis, P.; Lennon, R.; Plagge, A.; Wang, Q.; et al. Generation of Functioning Nephrons by Implanting Human Pluripotent Stem Cell-Derived Kidney Progenitors. Stem Cell Rep. 2018, 10, 766–779. [Google Scholar] [CrossRef] [Green Version]
- Murakami, Y.; Naganuma, H.; Tanigawa, S.; Fujimori, T.; Eto, M.; Nishinakamura, R. Reconstitution of the embryonic kidney identifies a donor cell contribution to the renal vasculature upon transplantation. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Francipane, M.G.; Han, B.; Oxburgh, L.; Sims-Lucas, S.; Li, Z.; Lagasse, E. Kidney-in-a-lymph node: A novel or-ganogenesis assay to model human renal development and test nephron progenitor cell fates. J. Tissue Eng. Regen. Med. 2019, 9, 1724–1731. [Google Scholar] [CrossRef]
- Komori, J.; Boone, L.; DeWard, A.; Hoppo, T.; Lagasse, E. The mouse lymph node as an ectopic transplantation site for multiple tissues. Nat. Biotechnol. 2012, 30, 976–983. [Google Scholar] [CrossRef] [Green Version]
- Cyster, J.G. Chemokines and Cell Migration in Secondary Lymphoid Organs. Science 1999, 286, 2098–2102. [Google Scholar] [CrossRef]
- Von Andrian, U.H.; Mempel, T.R. Homing and cellular traffic in lymph nodes. Nat. Rev. Immunol. 2003, 3, 867–878. [Google Scholar] [CrossRef]
- Garreta, E.; Prado, P.; Tarantino, C.; Oria, R.; Fanlo, L.; Martí, E.; Zalvidea, D.; Trepat, X.; Roca-Cusachs, P.; Ga-valdà-Navarro, A.; et al. Fine tuning the extracellular environment accelerates the derivation of kidney organoids from human pluripo-tent stem cells. Nat. Mater. 2019, 18, 397–405. [Google Scholar] [CrossRef] [PubMed]
- Phipson, B.; Er, P.X.; Combes, A.N.; Forbes, T.A.; Howden, S.E.; Zappia, L.; Yen, H.-J.; Lawlor, K.; Hale, L.J.; Sun, J.; et al. Evaluation of variability in human kidney organoids. Nat. Methods 2019, 16, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Laschke, M.W.; Vollmar, B.; Menger, M.D. Inosculation: Connecting the Life-Sustaining Pipelines. Tissue Eng. Part B Rev. 2009, 15, 455–465. [Google Scholar] [CrossRef]
- Nashimoto, Y.; Hayashi, T.; Kunita, I.; Nakamasu, A.; Torisawa, Y.-S.; Nakayama, M.; Takigawa-Imamura, H.; Kotera, H.; Nishiyama, K.; Miura, T.; et al. Integrating perfusable vascular networks with a three-dimensional tissue in a microfluidic device. Integr. Biol. 2017, 9, 506–518. [Google Scholar] [CrossRef] [PubMed]
- Mu, X.; Zheng, W.; Xiao, L.; Zhang, W.; Jiang, X. Engineering a 3D vascular network in hydrogel for mimicking a nephron. Lab. Chip. 2013, 13, 1612–1618. [Google Scholar] [CrossRef] [PubMed]
- Homan, K.A.; Gupta, N.; Kroll, K.T.; Kolesky, D.B.; Skylar-Scott, M.; Miyoshi, T.; Mau, D.; Valerius, M.T.; Ferrante, T.; Bonventre, J.V.; et al. Flow-enhanced vascularization and maturation of kidney organoids in vitro. Nat. Methods 2019, 16, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Wikswo, J.P. The relevance and potential roles of microphysiological systems in biology and medicine. Exp. Biol. Med. 2014, 239, 1061–1072. [Google Scholar] [CrossRef] [PubMed]
- Glass, N.R.; Takasako, M.; Er, P.X.; Titmarsh, D.M.; Hidalgo, A.; Wolvetang, E.J.; Little, M.H.; Cooper-White, J.J. Multivariate patterning of human pluripotent cells under perfusion reveals critical roles of induced paracrine factors in kidney organoid development. Sci. Adv. 2020, 6, eaaw2746. [Google Scholar] [CrossRef] [Green Version]
- Jansen, K.; Castilho, M.; Aarts, S.; Kaminski, M.M.; Lienkamp, S.S.; Pichler, R.; Malda, J.; Vermonden, T.; Jansen, J.; Masereeuw, R. Fabrication of Kidney Proximal Tubule Grafts Using Biofunctionalized Electrospun Polymer Scaffolds. Macromol. Biosci. 2019, 19, e1800412. [Google Scholar] [CrossRef]
- Petrosyan, A.; Cravedi, P.; Villani, V.; Angeletti, A.; Manrique, J.; Renieri, A.; De Filippo, R.E.; Perin, L.; Da Sacco, S. A glomerulus-on-a-chip to recapitulate the human glomerular filtration barrier. Nat. Commun. 2019, 10, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Rayner, S.; Phong, K.; Xue, J.; Lih, D.; Shankland, S.J.; Kelly, E.J.; Himmelfarb, J.; Zheng, Y. Reconstructing the Human Renal Vascular–Tubular Unit In Vitro. Adv. Healthc. Mater. 2018, 7, e1801120. [Google Scholar] [CrossRef]
- Ligresti, G.; Nagao, R.J.; Xue, J.; Choi, Y.J.; Xu, J.; Ren, S.; Aburatani, T.; Anderson, S.K.; Macdonald, J.W.; Bammler, T.K.; et al. A Novel Three–Dimensional Human Peritubular Microvascular System. J. Am. Soc. Nephrol. 2015, 27, 2370–2381. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Top-Down Approaches | Bottom-Up Approaches |
|---|---|
dECM
| Organoids
|
Vascular Corrosion Casts
| Microphysiological Systems
|
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lebedenko, C.G.; Banerjee, I.A. Enhancing Kidney Vasculature in Tissue Engineering—Current Trends and Approaches: A Review. Biomimetics 2021, 6, 40. https://doi.org/10.3390/biomimetics6020040
Lebedenko CG, Banerjee IA. Enhancing Kidney Vasculature in Tissue Engineering—Current Trends and Approaches: A Review. Biomimetics. 2021; 6(2):40. https://doi.org/10.3390/biomimetics6020040
Chicago/Turabian StyleLebedenko, Charlotta G., and Ipsita A. Banerjee. 2021. "Enhancing Kidney Vasculature in Tissue Engineering—Current Trends and Approaches: A Review" Biomimetics 6, no. 2: 40. https://doi.org/10.3390/biomimetics6020040