
The Increase in Kaffir Lime Leaf Production Due to Gibberellin Is Diminished by Pruning

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Planting Materials
2.2. Research Procedure
2.3. Data Mining and Analysis
3. Results
3.1. Contra Effect between Gibberellin and Pruning on the Plant Height and Shoot Number
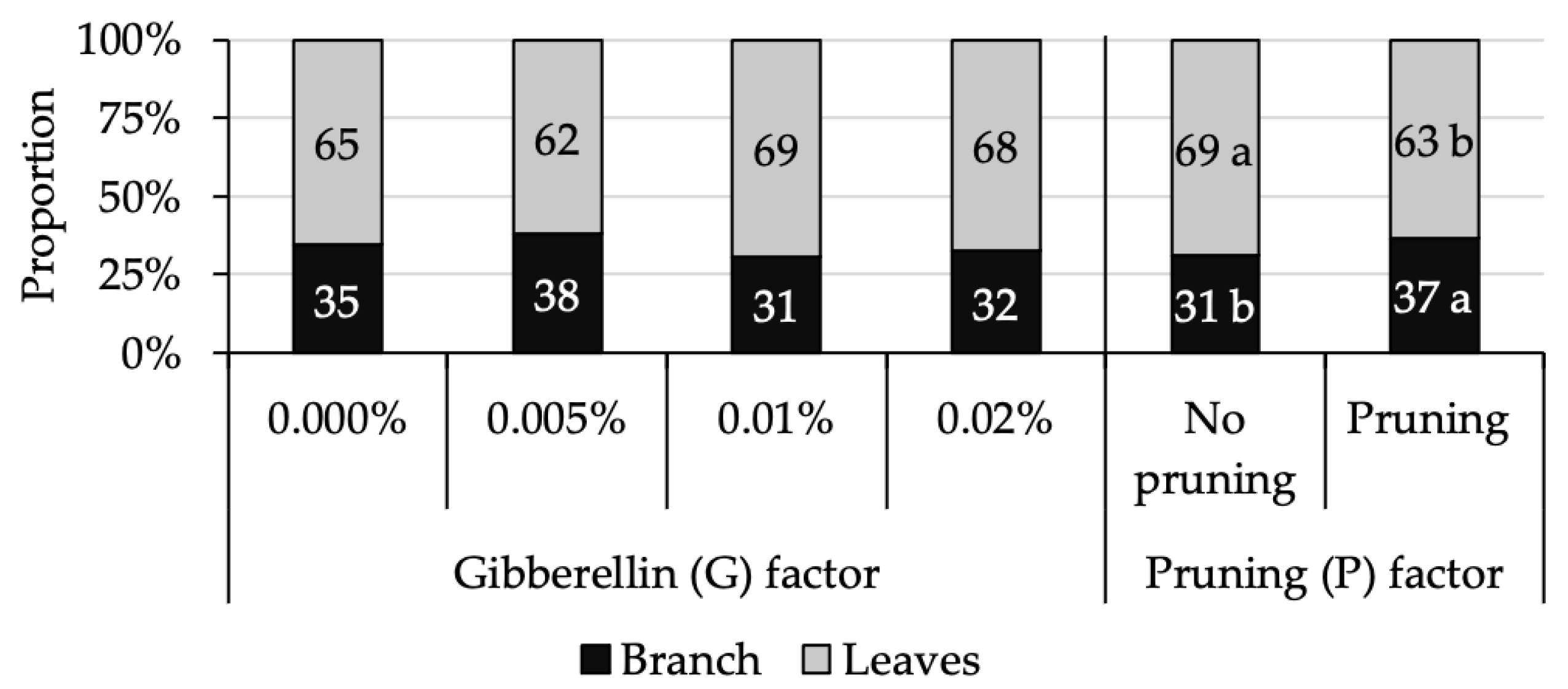
3.2. The Effect of Pruning Is More Dominant Than Gibberellin on Flushing Performance
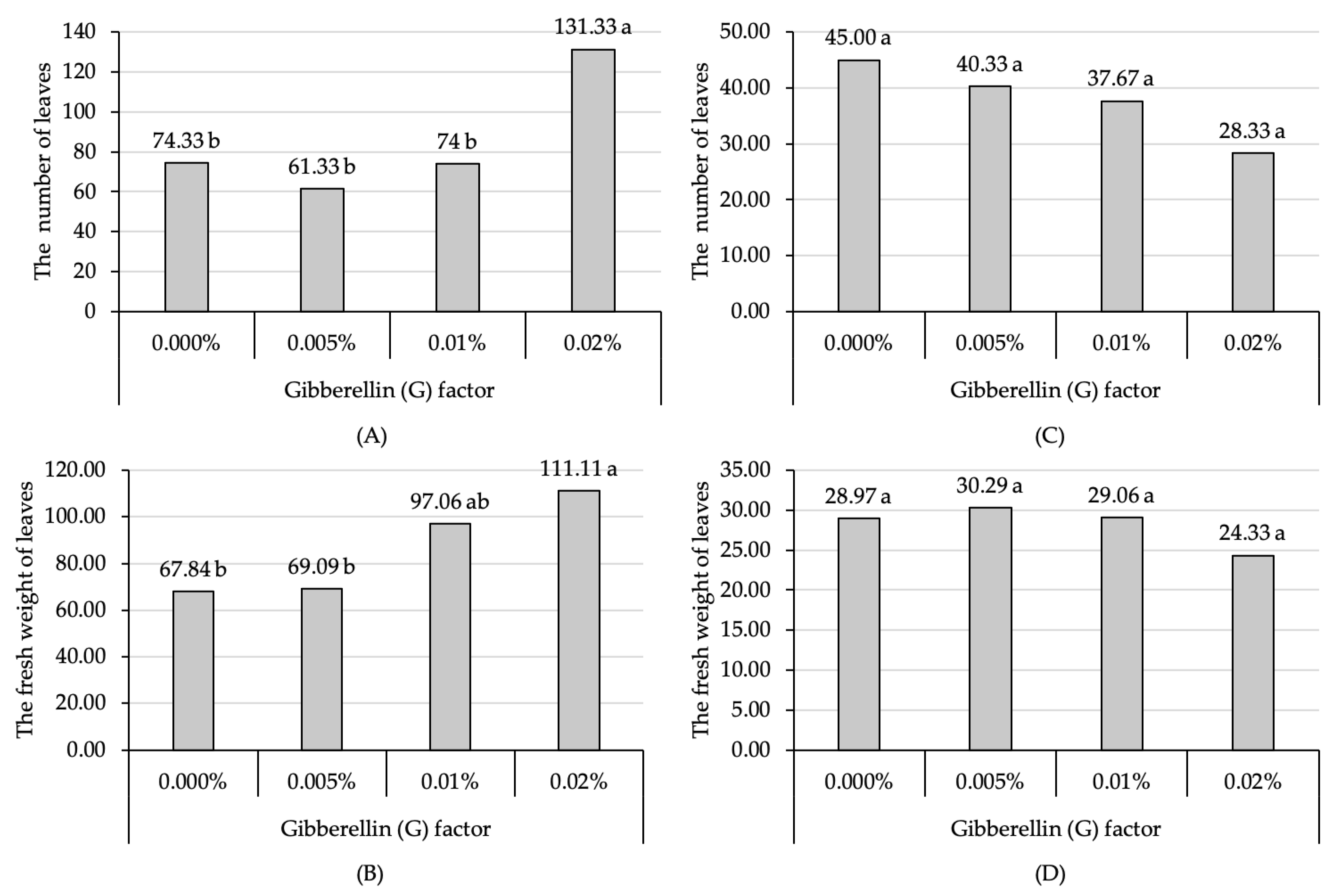
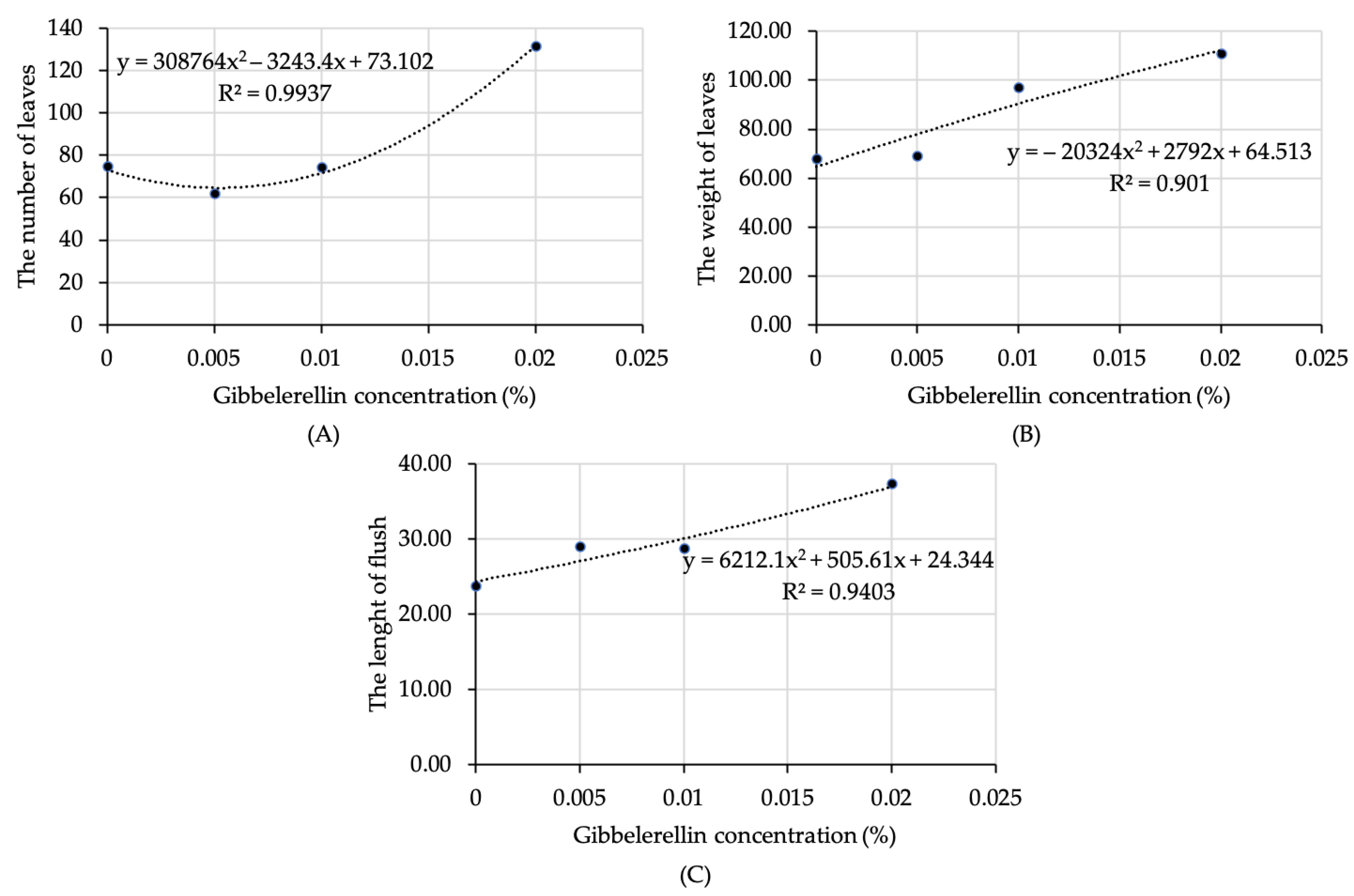
3.3. The Increase in Leaf Production Due to Gibberellin Is Diminished by Pruning
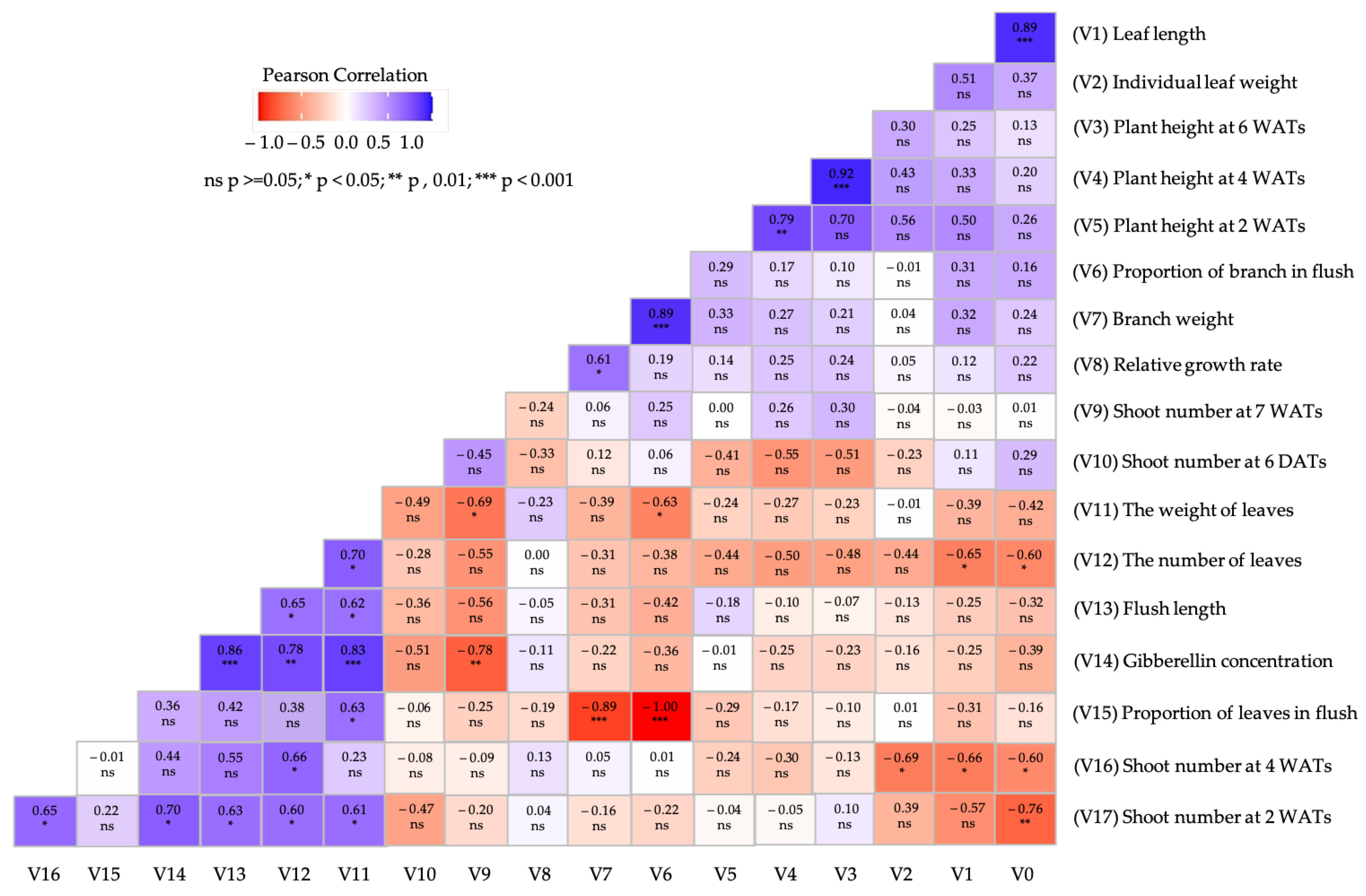
3.4. Multivariate Analysis Confirmed the Positive Effect of Gibberellin on Leaf Production of Unpruned Plants
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Budiarto, R.; Poerwanto, R.; Santosa, E.; Efendi, D.; Agusta, A. Production, post-harvest and marketing of kaffir lime (Citrus hystrix DC) in Tulungagung, Indonesia. J. Trop. Crop Sci. 2019, 6, 138–143. [Google Scholar] [CrossRef]
- Budiarto, R.; Poerwanto, R.; Santosa, E.; Efendi, D.; Agusta, A. Sensory evaluation of the quality of kaffir lime (Citrus hystrix DC.) leaves exposed to different postharvest treatments. J. Trop. Crop Sci. 2021, 8, 71–79. [Google Scholar] [CrossRef]
- Wongpornchai, S. Kaffir lime leaf. In Handbook of Herbs and Spices, 2nd ed.; Peter, K.V., Ed.; Woodhead Publishing Limited: Cambridge, UK, 2012. [Google Scholar]
- Araujo, E.F.D.; Queiroz, L.P.D.; Machado, M.A. What is Citrus? taxonomic implications from a study of cp-DNA evolution in the tribe Citreae (Rutaceae subfamily Aurantioideae). Org. Divers. Evol. 2003, 3, 55–62. [Google Scholar] [CrossRef]
- Efendi, D.; Budiarto, R. Benefit and challenges of using tropical fruits as ornamental trees for green city. Acta Hortic. 2022, 1334, 369–377. [Google Scholar] [CrossRef]
- Budiarto, R.; Poerwanto, R.; Santosa, E.; Efendi, D.; Agusta, A. Short communication: Allometric model to estimate bifoliate leaf area and weight of kaffir lime (Citrus hystrix). Biodiversitas 2021, 22, 2815–2820. [Google Scholar] [CrossRef]
- Budiarto, R.; Poerwanto, R.; Santosa, E.; Efendi, D. Morphological evaluation and determination keys of 21 citrus genotypes at seedling stage. Biodiversitas 2021, 22, 1570–1579. [Google Scholar] [CrossRef]
- Ansori, A.N.M.; Supriyadi, A.P.; Kartjito, M.V.; Rizqi, F.; Adrianto, H. Biolarvacidal effectivities of polar and non-polar extract fraction from kaffir lime (Citrus hystrix) leaf against 3rd instar larvae of Aedes aegypti. J. Biol. Eng. Res. Rev. 2015, 2, 14–17. [Google Scholar]
- Kooltheat, N.; Kamuthachad, L.; Anthapanya, M.; Samakchan, N.; Sranujit, R.P.; Potup, P.; Ferrante, A.; Usuwanthim, K. Kaffir lime leaves extract inhibits biofilm formation by Streptococcus mutans. Nutrition 2016, 32, 486–490. [Google Scholar] [CrossRef] [PubMed]
- Budiarto, R.; Sholikin, M.M. Kaffir lime essential oil variation in the last fifty years: A meta-analysis of plant origins, plant parts and extraction methods. Horticulturae 2022, 8, 1132. [Google Scholar] [CrossRef]
- Othman, S.N.A.M.; Hassan, M.A.; Nahar, L.; Basar, N.; Jamil, S.; Sarker, S.D. Essential oils from the Malaysian Citrus (Rutaceae) medicinal plants. Medicines 2016, 3, 13. [Google Scholar] [CrossRef] [PubMed]
- Wany, A.; Jha, S.; Nigam, V.K.; Pandey, D.M. Chemical analysis and therapeutic uses of citronella oil from Cymbopogon winterianus: A short review. Int. J. Adv. Res. 2013, 1, 504–521. [Google Scholar]
- Agouillal, F.; Taher, Z.M.; Moghrani, H.; Nasrallah, N.; Enshasy, H.E. A review of genetic taxonomy, biomolecules chemistry and bioactivities of Citrus hystrix DC. Biosci. Biotechnol. Res. Asia 2017, 14, 285–305. [Google Scholar] [CrossRef]
- Rammanee, K.; Hongpattarakere, T. Effects of tropical citrus essential oils on growth, aflatoxin production, and ultrastucture alterations of Aspergillus flavus and Aspergillus parasiticus. Food Bioproc. Technol. 2011, 4, 1050–1059. [Google Scholar] [CrossRef]
- OuYang, Q.; Liu, Y.; Oketch, O.R.; Zhang, M.; Shao, X.; Tao, N. Citronellal exerts its antifungal activity by targeting ergosterol biosynthesis in Penicillium digitatum. J. Fungi 2021, 7, 432. [Google Scholar] [CrossRef]
- Vimol, S.; Chanwit, T.; Veena, N.; Nuntavan, B.; Kulkanya, C.; Siwimol, P.; Sirirat, C.; Somporn, S. Antibacterial activity of essential oils from Citrus hystrix (makrut lime) against respiratory tract pathogens. Sci. Asia 2012, 38, 212–217. [Google Scholar]
- Lota, M.L.; Serra, D.R.D.; Tomi, F.; Jacquemond, C.; Casanova, J. Volatile components of peel and leaf oils of lemon and lime species. J. Agric. Food Chem. 2002, 50, 796–805. [Google Scholar] [CrossRef]
- Nerio, L.S.; Olivero-Verbel, J.; Stashenko, E. Repellent activity of essential oils: A review. Biores. Technol. 2010, 101, 372–378. [Google Scholar] [CrossRef]
- Dickens, J.C.; Bohbot, J.D. Mini review: Mode of action of mosquito repellents. Pestic. Biochem. Phys. 2013, 106, 149–155. [Google Scholar] [CrossRef]
- Sharma, R.; Rao, R.; Kumar, S.; Mahant, S.; Khatkar, S. Therapeutic potential of citronella essential oil: A review. Curr. Drug Discov. Technol. 2019, 16, 330–339. [Google Scholar] [CrossRef]
- Yahya, A.; Rubiyanto, D.; Fatimah, I. Heterogeneous catalytic conversion of citronellal into isopulegol and methol: Literature review. Sci. Technol. Indones. 2021, 6, 166–180. [Google Scholar] [CrossRef]
- Lenardao, E.J.; Botteselle, G.V.; Azambuja, F.D.; Perin, G.; Jacob, R.G. Citronellal as key compound in organic synthesis. Tetrahedron 2007, 63, 6671–6712. [Google Scholar] [CrossRef]
- Jacob, R.G.; Perin, G.; Loi, L.N.; Pinno, C.S.; Lenardao, E.J. Green synthesis of (−)-isopulegol from (+)-citronellal: Application to essential oil of citronella. Tetrahedron Lett. 2003, 44, 3605–3608. [Google Scholar] [CrossRef]
- Foley, J.A.; Ramankutty, N.; Brauman, K.A.; Cassidy, E.S.; Gerber, J.S.; Johnston, M.; Mueller, N.D.; O’Connell, C.; Ray, D.K.; West, P.C.; et al. Solutions for a cultivated planet. Nature 2011, 478, 337–342. [Google Scholar] [CrossRef]
- Rademacher, W. Plant growth regulators: Backgrounds and uses in plant production. J. Plant Growth Regul. 2015, 34, 845–872. [Google Scholar] [CrossRef]
- Chadwick, D.; Wei, J.; Yan’an, T.; Guanghui, Y.; Qirong, S.; Qing, C. Improving manure nutrient management towards sustainable agricultural intensification in China. Agric. Ecosyst. Environ. 2015, 209, 34–46. [Google Scholar] [CrossRef]
- Abberton, M.; Batley, J.; Bentley, A.; Bryant, J.; Cai, H.; Cockram, J.; de Oliveira, A.C.; Cseke, L.J.; Dempewolf, H.; Pace, C.D.; et al. Global agricultural intensification during climate change: A role for genomics. Plant Biotechnol. J. 2016, 14, 1095–1098. [Google Scholar] [CrossRef]
- Garmendia, A.; Beltran, R.; Zornoza, C.; Garcia-Breijo, F.J.; Reig, J.; Merle, H. Gibberellic acid in Citrus spp. flowering and fruiting: A systematic review. PLoS ONE 2019, 14, e0223147. [Google Scholar] [CrossRef]
- Takahashi, N.; Kitamura, H.; Kawarada, A.; Seta, Y.; Takai, M.; Tamura, S.; Sumiki, Y. Biochemical studies on “Bakanae” fungus. Bull. Agricult. Chem. Soc. Jpn. 1955, 19, 267–281. [Google Scholar]
- Tang, L.; Lovatt, C.J. Effects of low temperature and gibberellic acid on floral gene expression and floral determinacy in ‘Washington’ navel orange (Citrus sinensis L. Osbeck). Sci. Hortic. 2019, 243, 92–100. [Google Scholar] [CrossRef]
- Chao, C.C.T.; Khuong, T.; Zheng, Y.; Lovatt, C.J. Response of evergreen perennial tree crops to gibberellic acid is crop load-dependent. I: GA3 increases the yield of commercially valuable “Nules” Clementine Mandarin fruit only in the off-crop year of an alternate-bearing orchard. Sci. Hortic. 2011, 130, 743–752. [Google Scholar] [CrossRef]
- Duarte, A.M.M.; García-Luis, A.; Molina, R.V.; Monerri, C.; Navarro, V.; Nebauer, S.G.; Sánchez-Perales, M.; Guardiola, J.L. Long-term effect of winter gibberellic acid sprays and auxin applications on crop value of “Clausellina” satsuma. J. Am. Soc. Hortic. Sci. 2006, 131, 586–592. [Google Scholar] [CrossRef]
- Maia, E.; de Siqueira, D.L.; Cecon, P.R. Production, florescence and fructification of Pokan mandarin tree submitted to gibberellic acid application. Cienc. Rural. 2010, 40, 507–512. [Google Scholar] [CrossRef]
- Muñoz-Fambuena, N.; Mesejo, C.; González-Mas, M.C.; Iglesias, D.J.; Primo-Millo, E.; Agustí, M. Gibberellic acid reduces flowering intensity in sweet orange [Citrus sinensis (L.) osbeck] by repressing cift gene expression. J. Plant Growth Regul. 2012, 31, 529–536. [Google Scholar] [CrossRef]
- Gambetta, G.; Mesejo, C.; Martínez-Fuentes, A.; Reig, C.; Gravina, A.; Agustí, M. Gibberellic acid and norflurazon affecting the time-course of flavedo pigment and abscisic acid content in “Valencia” sweet orange. Sci. Hortic. 2014, 180, 94–101. [Google Scholar] [CrossRef]
- Zhang, J.; Cao, Y.; Tang, J.; He, X.; Li, M.; Li, C.; Ren, X.; Ding, Y. Physiology and application of gibberellins in postharvest horticultural crops. Horticulturae 2023, 9, 625. [Google Scholar] [CrossRef]
- Davies, F.S.; Zalman, G. Gibberellic acid, fruit freezing, and post-freeze quality of “Hamlin” oranges. HortTechnology 2006, 16, 301–305. [Google Scholar] [CrossRef]
- Khalid, S.; Malik, A.U.; Khan, A.S.; Jamil, A. Influence of exogenous applications of plant growth regulators on fruit quality of young “Kinnow” mandarin (Citrus nobilis × C. deliciosa) trees. Int. J. Agric. Biol. 2012, 14, 229–234. [Google Scholar]
- Akter, N.; Islam, M.R.; Karim, M.A.; Hossain, T. Alleviation of drought stress in maize by exogenous application of gibberellic acid and cytokinin. J. Crop Sci. Biotechnol. 2014, 17, 41–48. [Google Scholar] [CrossRef]
- Rady, M.M.; Boriek, S.H.K.; Abd El-Mageed, T.A.; Seif El-Yazal, M.A.; Ali, E.F.; Hassan, F.A.S.; Abdelkhalik, A. Exogenous gibberellic acid or dilute bee honey boosts drought stress tolerance in Vicia faba by rebalancing osmoprotectants. Plants 2021, 10, 748. [Google Scholar] [CrossRef]
- Sprangers, K.; Thys, S.; Dusschoten, D.V.; Beemster, G.T.S. Giberrellin enhances the anisotopy of cell expansion in the growth zone of the maize leaf. Front. Plant Sci. 2020, 11, 1163. [Google Scholar] [CrossRef]
- Li, J.; Sima, W.; Ouyang, B.; Wang, T.; Ziaf, K.; Luo, Z.; Liu, L.; Li, H.; Chen, M.; Huang, Y.; et al. Tomato SIDREB gene restricts leaf expansion and internode elongation by downregulating key genes for gibberellin biosynthesis. J. Exp. Bot. 2012, 63, 6407–6420. [Google Scholar] [CrossRef]
- Budiarto, R.; Poerwanto, R.; Santosa, E.; Efendi, D.; Agusta, A. Agronomical and physiological characters of kaffir lime (Citrus hystrix DC) seedling under artificial shading and pruning. Emir. J. Food Agric. 2019, 31, 222–230. [Google Scholar] [CrossRef]
- Budiarto, R.; Poerwanto, R.; Santosa, E.; Efendi, D. Shoot manipulations improve flushing and flowering of mandarin citrus in Indonesia. J. Appl. Hortic. 2018, 20, 112–118. [Google Scholar] [CrossRef]
- Budiarto, R.; Poerwanto, R.; Santosa, E.; Efendi, D.; Agusta, A. Agro-physiological traits of kaffir lime in response to pruning and nitrogen fertilizer under mild shading. Plants 2023, 12, 1155. [Google Scholar] [CrossRef] [PubMed]
- Garrido, A.; Conde, A.; Serôdio, J.; De Vos, R.C.H.; Cunha, A. Fruit Photosynthesis: More to Know about Where, How and Why. Plants 2023, 12, 2393. [Google Scholar] [CrossRef] [PubMed]
- Simkin, A.J.; Faralli, M.; Ramamoorthy, S.; Lawson, T. Photosynthesis in non-foliar tissues: Implications for yield. Plant J. 2020, 101, 1001–1015. [Google Scholar] [CrossRef]
- Aschan, G.; Pfanz, H. Non-foliar photosynthesis—A strategy of additional carbon acquisition. Flora 2003, 198, 81–97. [Google Scholar] [CrossRef]
- Brazel, A.J.; Ó’Maoileídigh, D.S. Photosynthetic activity of reproductive organs. J. Exp. Bot. 2019, 70, 1737–1753. [Google Scholar] [CrossRef]
- Smith, M.R.; Rao, I.M.; Merchant, A. Source-sink relationships in crop plants and their influence on yield development and nutritional quality. Front. Plant Sci. 2018, 9, 1889. [Google Scholar] [CrossRef]
- Yu, S.M.; Lo, S.F.; Ho, T.H.D. Source–sink communication: Regulated by hormone, nutrient, and stress cross-signaling. Trends Plant Sci. 2015, 18, 844–857. [Google Scholar] [CrossRef]
- White, A.C.; Rogers, A.; Rees, M.; Osborne, C.P. How can we make plants grow faster? A source–sink perspective on growth rate. J. Exp. Bot. 2016, 67, 31–45. [Google Scholar] [CrossRef]
- Yuan, R.; Alferez, F.; Kostenyuk, I.; Singh, S.; Syvertsen, J.P.; Burns, J.K. Partial defoliation can decrease average leaf size but has little effect on orange tree growth, fruit yield and juice quality. HortScience 2005, 40, 2011–2015. [Google Scholar] [CrossRef]
- Eissenstat, D.M.; Duncan, L.W. Root growth and carbohydrate responses in bearing citrus trees following partial canopy removal. Tree Physiol. 1992, 10, 245–257. [Google Scholar] [CrossRef] [PubMed]
- Ramos, Y.C.; Stuchi, E.S.; Girardi, E.A.; de Leão, H.C.; Gesteira, A.D.S.; Passos, O.S.; Filho, W.D.S. Dwarfing rootstocks for ‘valencia’ sweet orange. Acta Hortic. 2015, 1065, 351–354. [Google Scholar] [CrossRef]
- Pedroso, F.K.J.V.; Prudente, D.A.; Bueno, A.C.R.; Machado, E.C.; Ribeiro, R.V. Drought tolerance in citrus trees is enhanced by rootstock-dependent changes in root growth and carbohydrate availability. Environ. Exp. Bot. 2014, 101, 26–35. [Google Scholar] [CrossRef]
- Silva, S.F.; Miranda, M.T.; Costa, V.E.; Machado, E.C.; Ribeiro, R.V. Sink strength of citrus rootstocks under water deficit. Tree Physiol. 2021, 41, 1372–1383. [Google Scholar] [CrossRef]
- Miranda, M.T.; Da Silva, S.F.; Silveira, N.M.; Pereira, L.; Machado, E.C.; Ribeiro, R.V. Root osmotic adjustment and stomatal control of leaf gas exchange are dependent on citrus rootstocks under water deficit. J. Plant Growth Regul. 2021, 40, 11–19. [Google Scholar] [CrossRef]
- Allario, T.; Brumos, J.; Colmenero-Flores, J.M.; Iglesias, D.J.; Pina, J.A.; Navarro, L.; Talon, M.; Ollitrault, P.; Morillon, R. Tetraploid Rangpur lime rootstock increases drought tolerance via enhanced constitutive root abscisic acid production. Plant Cell. Environ. 2013, 36, 856–868. [Google Scholar] [CrossRef]
- Sahin-Cevik, M.; Cevik, B.; Coskan, A. Identification and expression analysis of salinity-induced genes in Rangpur lime (Citrus limonia). Hortic. Plant J. 2020, 6, 267–276. [Google Scholar] [CrossRef]
- Qureshi, M.A.; Jaskani, M.J.; Khan, A.S.; Ahmad, R. Influence of endogenous plant hormones on physiological and growth attributes of kinnow mandarin grafted on nine rootstocks. J. Plant Growth Regul. 2022, 41, 1254–1264. [Google Scholar] [CrossRef]
- Gao, S.; Chu, C. Gibberellin metabolism and signaling: Targets for improving agronomic performance of crops. Plant Cell Physiol. 2020, 61, 1902–1911. [Google Scholar] [CrossRef] [PubMed]
- Tallón, C.I.; Porras, I.; Pérez-Tornero, O. Efficient propagation and rooting of three citrus rootstocks using different plant growth regulators. In Vitro Cell. Dev. Biol.-Plant 2012, 48, 488–499. [Google Scholar] [CrossRef]
- Roro, A.G.; Dukker, S.A.F.; Melby, T.I.; Solhaug, K.A.; Torre, S.; Olsen, J.E. UV-B-induced inhibition of stem elongation and leaf expansion in pea depends on modulation of gibberellin metabolism and intact gibberellin signalling. J. Plant Growth Regul. 2017, 36, 680–690. [Google Scholar] [CrossRef]
- Sun, T.; Gubler, F. Molecular mechanism of gibberellin signaling in plants. Annu. Rev. Plant Biol. 2004, 55, 197–223. [Google Scholar] [CrossRef] [PubMed]
- Davière, J.; Achard, P. Gibberellin signaling in plants. Development 2013, 140, 1147–1151. [Google Scholar] [CrossRef]
- Ubeda-Tomás, S.; Federici, F.; Casimiro, I.; Beemster, G.T.S.; Bhalerao, R.; Swarup, R.; Doerner, P.; Haseloff, J.; Bennett, M.J. Gibberellin signaling in the endodermis controls Arabidopsis root meristem size. Curr. Biol. 2009, 19, 1194–1199. [Google Scholar] [CrossRef]
- Falcioni, R.; Moriwaki, T.; Bonato, C.M.; Souza, L.A.D.; Nanni, M.R.; Antunes, W.C. Distinct growth light and gibberellin regimes alter leaf anatomy and reveal their influence on leaf optical properties. Environ. Exp. Bot. 2017, 140, 86–95. [Google Scholar] [CrossRef]
- Wen, Y.; Su, S.; Ma, L.; Wang, X. Effects of gibberellic acid on photosynthesis and endogenous hormones of Camellia oleifera Abel. in 1st and 6th leaves. J. For. Res. 2018, 23, 309–317. [Google Scholar] [CrossRef]
- Zang, Y.; Chun, I.; Zhang, L.; Hong, S.; Zheng, W.; Xu, K. Effect of gibberellic acid application on plant growth attributes, return bloom, and fruit quality of rabbiteye blueberry. Sci. Hortic. 2016, 200, 13–18. [Google Scholar] [CrossRef]
- Rezvani, M.; Zaefarian, F.; Amini, V. Effects of chemical treatments and environmental factors on seed dormancy and germination of shepherd’s purse (Capsella bursa-pastoris (L.) Medic.). Acta Bot. Bras. 2014, 28, 495–501. [Google Scholar] [CrossRef]
- Sponsel, V.M.; Hedden, P. Gibberellin biosynthesis and inactivation. In Plant Hormones: Biosynthesis, Signal Transduction, Action! Davies, P.J., Ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2004; pp. 63–94. [Google Scholar]
- Gupta, R.; Chakrabarty, S.K. Gibberellic acid in plant: Still a mystery unresolved. Plant Signal. Behav. 2013, 8, e25504. [Google Scholar] [CrossRef] [PubMed]
- Colebrook, E.H.; Thomas, S.G.; Phillips, A.L.; Hedden, P.; Davies, S.A.; Dow, J.A.T.; Lukowiak, K. The role of gibberellin signaling in plant responses to abiotic stress. J. Exp. Biol. 2014, 217, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Ramon, U.; Weiss, D.; Illouz-Eliaz, N. Underground gibberellin activity: Differential gibberellin response in tomato shoots and roots. New Phytol. 2021, 229, 1196–1200. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Plant Height (cm) | Plant Shoot Number (Shoots) | ||||
|---|---|---|---|---|---|---|
| 2 WATs | 4 WATs | 6 WATs | 2 WATs | 4 WATs | 6 WATs | |
| Gibberellin (G) factor | ||||||
| 0.000% | 50.67 b | 65 b | 66.67 b | 3.5 a | 5.17 ab | 8.67 a |
| 0.005% | 61.5 a | 73.5 a | 76 a | 4.33 a | 5 ab | 4.17 b |
| 0.01% | 55.83 b | 64.33 b | 64.17 b | 4 a | 4 b | 7.5 a |
| 0.02% | 53.08 b | 64.67 b | 66.17 b | 6.17 a | 6.17 a | 3.33 b |
| Pruning (P) factor | ||||||
| No pruning | 75.42 a | 84 a | 85.17 a | 4.58 a | 6.17 a | 4 b |
| Pruning | 35.13 b | 49 b | 51.33 b | 4.42 a | 4 b | 7.83 a |
| Treatment | Flush Number (Flush) | Flush Length (cm) | Leaf Length (cm) | Leaf Area (cm2) | Individual Leaf Weight (g) |
|---|---|---|---|---|---|
| Gibberellin (G) factor | |||||
| 0.000% | 6.33 a | 18.58 b | 10.05 b | 26.34 a | 0.79 a |
| 0.005% | 4.50 a | 22.67 b | 10.55 ab | 29.69 a | 0.94 a |
| 0.01% | 4.83 a | 19.75 b | 11.38 a | 31.41 a | 1.05 a |
| 0.02% | 6.00 a | 31.50 a | 10.70 ab | 28.43 a | 0.86 a |
| Pruning (P) factor | |||||
| No pruning | 6.67 a | 29.58 a | 11.66 a | 33.27 a | 1.06 a |
| Pruning | 4.17 b | 16.67 b | 9.68 b | 24.66 b | 0.76 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Budiarto, R.; Mubarok, S.; Nanda, M.A.; Nabiyyu, M.; Jaya, M.H.I.S. The Increase in Kaffir Lime Leaf Production Due to Gibberellin Is Diminished by Pruning. Horticulturae 2023, 9, 1018. https://doi.org/10.3390/horticulturae9091018
Budiarto R, Mubarok S, Nanda MA, Nabiyyu M, Jaya MHIS. The Increase in Kaffir Lime Leaf Production Due to Gibberellin Is Diminished by Pruning. Horticulturae. 2023; 9(9):1018. https://doi.org/10.3390/horticulturae9091018
Chicago/Turabian StyleBudiarto, Rahmat, Syariful Mubarok, Muhammad Achirul Nanda, Maulidan Nabiyyu, and Moh Haris Imron S Jaya. 2023. "The Increase in Kaffir Lime Leaf Production Due to Gibberellin Is Diminished by Pruning" Horticulturae 9, no. 9: 1018. https://doi.org/10.3390/horticulturae9091018