
Phytochemical Properties of Silk Floss Tree Stem Bark Extract and Its Potential as an Eco-Friendly Biocontrol Agent against Potato Phytopathogenic Microorganisms
Abstract
:1. Introduction
2. Materials and Methods
2.1. Source of Plant Bacterial Phytopathogens
2.2. Plant Fungal Pathogens
2.3. Preparation of Silk Floss Extract
2.4. Antibacterial Activity of SFSB Extract
2.5. Silk Floss Stem Bark Extract (SFSB) Antifungal Activity
2.6. SFSB Extract Antioxidant Activity
2.7. HPLC Analysis
2.8. Gas Chromatography Mass Spectroscopy (GCMS) Analysis
2.9. Statistical Analyses
3. Results and Discussion
3.1. SFSB Extract Bacterial Inhibitory Effect In Vitro
3.2. Fungal Pathogens
3.2.1. Isolation and Identification
3.2.2. Molecular Identification and Phylogenetic Analysis
3.3. Effect of SFSB Extract on the Fungal Pathogens
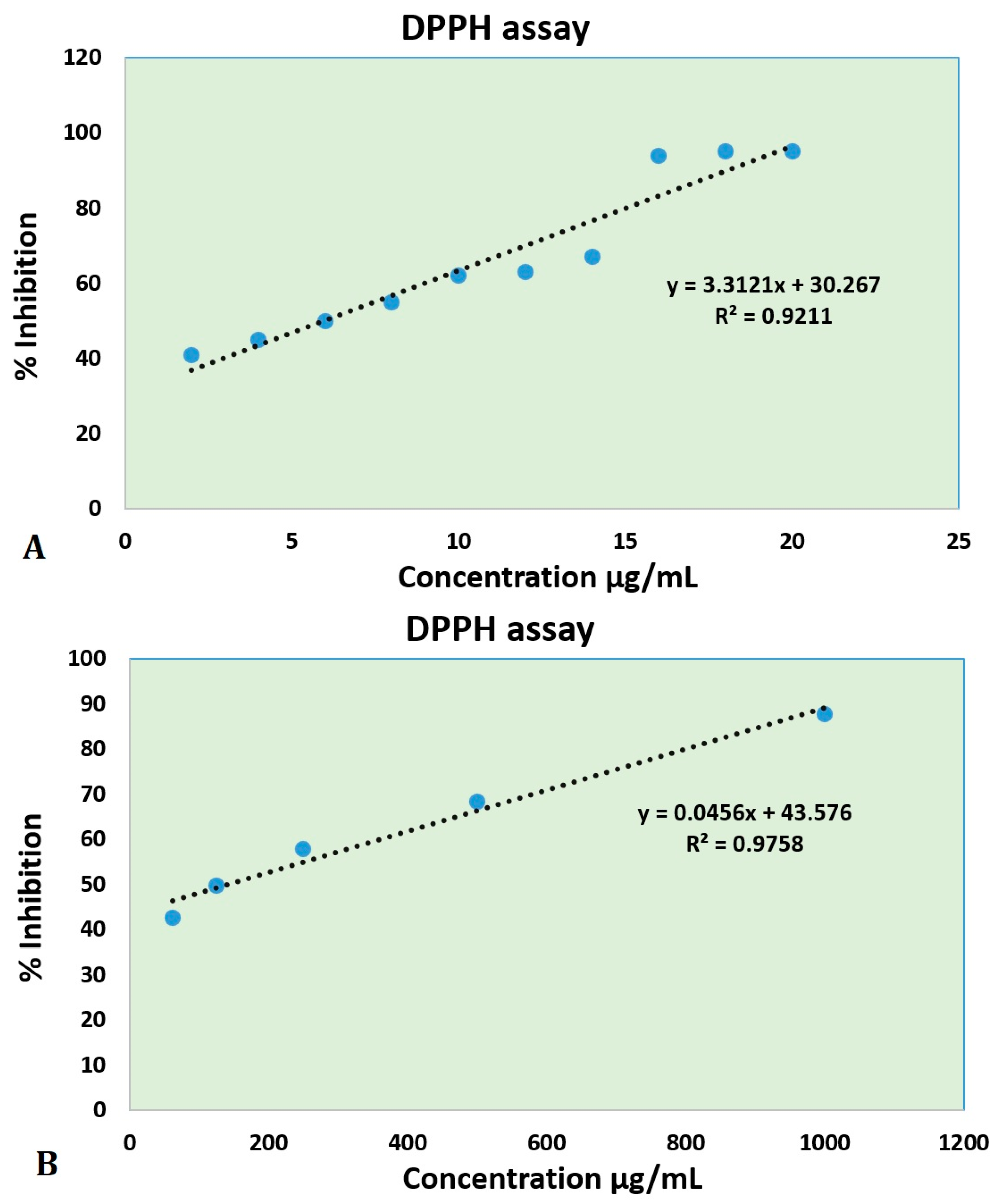
3.4. Antioxidant Activity
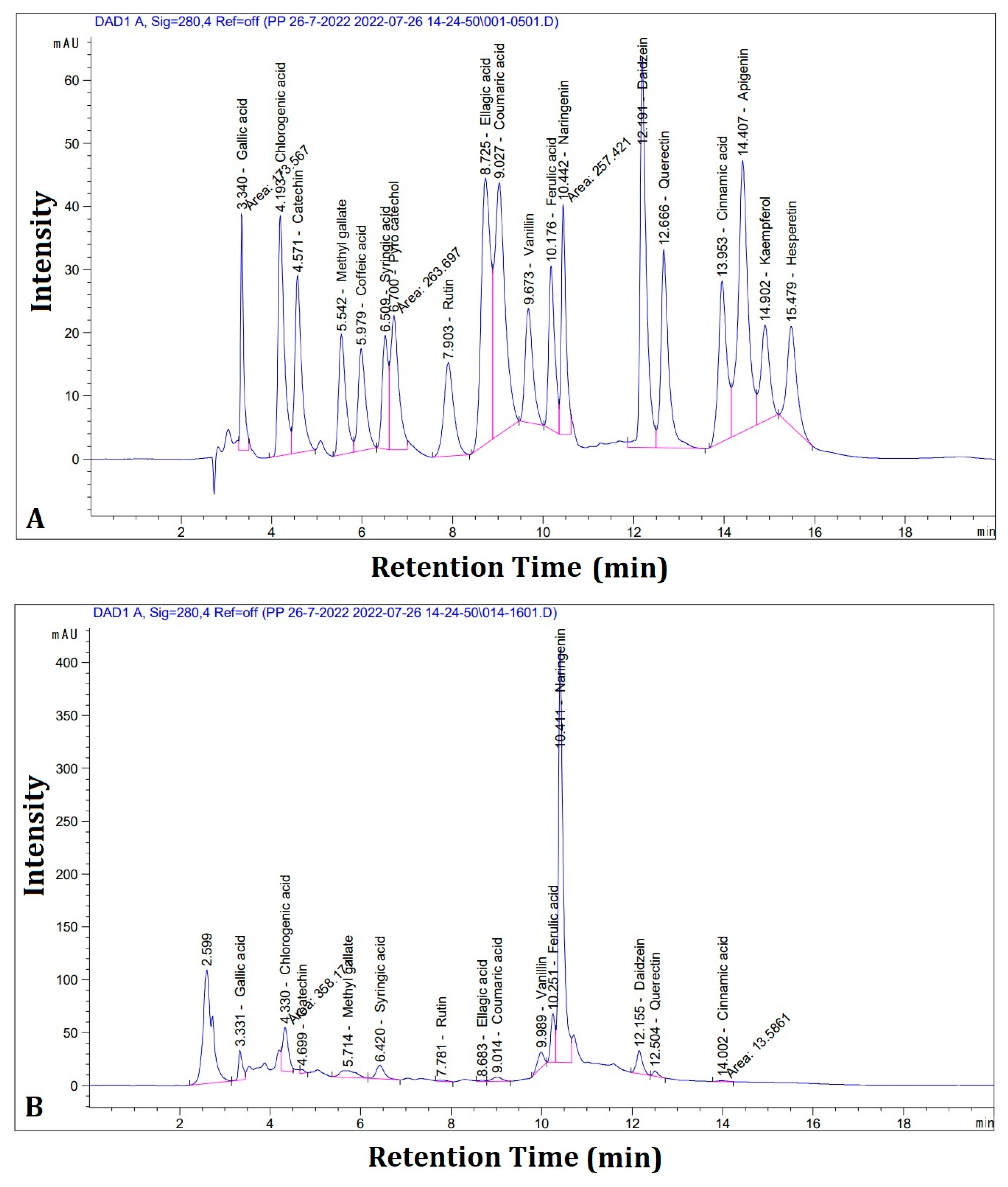
3.5. Silk Floss (SFSB) Extract Polyphenolic Content
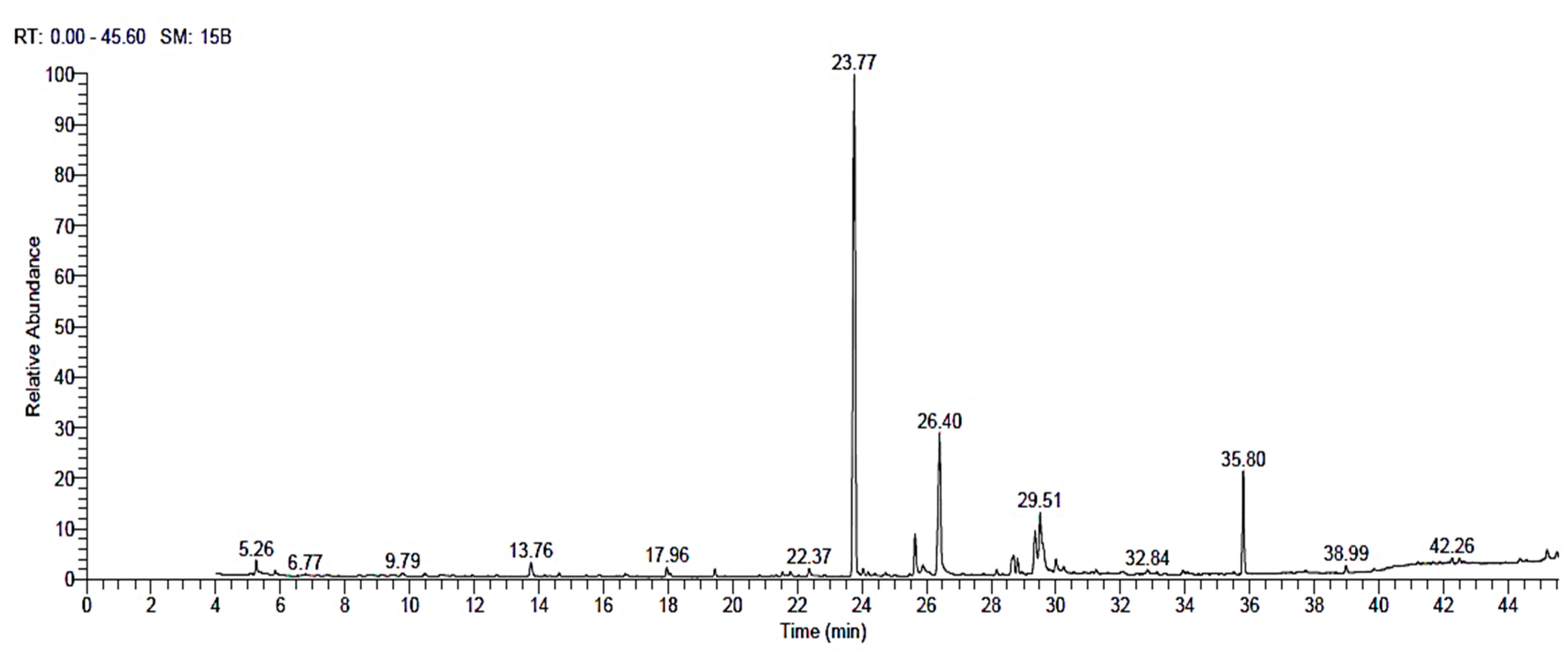
3.6. Silk Floss (SFSB) Extract GC-MS Analysis
4. Conclusions
Supplementary Materials
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Talib, W.H.; Mahasneh, A.M. Antimicrobial, cytotoxicity and phytochemical screening of Jordanian plants used in traditional medicine. Molecules 2010, 15, 1811–1824. [Google Scholar]
- Parveen, S.; Wani, A.H.; Bhat, M.Y.; Malik, A.R.; Koka, J.A.; Ashraf, N. Antimycotic potential of some phytoextracts on some pathogenic fungi. J. Biopestic. 2017, 10, 60–65. [Google Scholar]
- Abdelkhalek, A.; Salem, M.Z.M.M.; Hafez, E.; Behiry, S.I.; Qari, S.H. The Phytochemical, Antifungal, and First Report of the Antiviral Properties of Egyptian Haplophyllum tuberculatum Extract. Biology 2020, 9, 248. [Google Scholar] [CrossRef]
- Alzoreky, N.S.; Nakahara, K. Antibacterial activity of extracts from some edible plants commonly consumed in Asia. Int. J. Food Microbiol. 2003, 80, 223–230. [Google Scholar] [PubMed]
- Castro, S.B.R.; Leal, C.A.G.; Freire, F.R.; Carvalho, D.A.; Oliveira, D.F.; Figueiredo, H.C.P. Antibacterial activity of plant extracts from Brazil against fish pathogenic bacteria. Braz. J. Microbiol. 2008, 39, 756–760. [Google Scholar]
- Gonelimali, F.D.; Lin, J.; Miao, W.; Xuan, J.; Charles, F.; Chen, M.; Hatab, S.R. Antimicrobial properties and mechanism of action of some plant extracts against food pathogens and spoilage microorganisms. Front. Microbiol. 2018, 9, 1639. [Google Scholar]
- Joly, A.B. Botany: An Introduction to Plant Taxonomy; National Publishing Company: São Paulo, Brazil, 1991; Volume 10, p. 462. [Google Scholar]
- Adjanohoun, E.J.; Ahyi, A.M.R.; Ake, A. Médecine Traditionnelle et Pharmacopée: Contribution aux Études Ethnobotaniques et Floristiques en République Populaire du Congo; Agence de Coopération Culturelle et Technique: Paris, France, 1988. [Google Scholar]
- Refaat, J.; Desoky, S.Y.; Ramadan, M.A.; Kamel, M.S. Bombacaceae: A phytochemical review. Pharm. Biol. 2013, 51, 100–130. [Google Scholar]
- Samia, S.H.; Afaf, E.A.G. Pharmacognostical and antibacterial studies of chorisia speciosa st. Hil. flower [Bomhacaceae]. Mansoura J. Pharm. Sci. 2003, 19, 40–59. [Google Scholar]
- Ashmawy, A.M.; Azab, S.S.; Eldahshan, O.A. Effects of Chorisia crispiflora ethyl acetate extract on P21 and NF-κB in breast cancer cells. J. Am. Sci. 2012, 8, 965–972. [Google Scholar]
- Refaat, J.; Yehia Desoukey, S.; Ramadan, M.A.; Kamel, M.S.; Han, J.; Isoda, H. Comparative polyphenol contents, free radical scavenging properties and effects on adipogenesis of Chorisia chodatii and Chorisia speciosa. J. Med. Herbs 2015, 5, 193–207. [Google Scholar]
- El-Alfy, T.S.; El-Sawi, S.A.; Sleem, A.; Moawad, D.M. Investigation of flavonoidal content and biological activities of Chorisia insignis Hbk. leaves. Aust. J. Basic Appl. Sci. 2010, 4, 1334–1348. [Google Scholar]
- Behiry, S.I.; Soliman, S.A.; Al-Mansori, A.-N.A.; Al-Askar, A.A.; Arishi, A.A.; Elsharkawy, M.M.; Abdelkhalek, A.; Heflish, A.A. Chorisia speciosa Extract Induces Systemic Resistance against Tomato Root Rot Disease Caused by Rhizoctonia solani. Agronomy 2022, 12, 2309. [Google Scholar]
- Nasr, E.M.; Assaf, M.H.; Darwish, F.M.; Ramadan, M.A. Phytochemical and biological study of Chorisia speciosa A. St. Hil. cultivated in Egypt. J. Pharmacogn. Pharmacol. 2018, 7, 649–656. [Google Scholar]
- Statistics, F.A.O. Food and Agriculture Organization of United Nations. Agriculture Data (FAO); FAO: Rome, Italy, 2011. [Google Scholar]
- Rahman, M.M.; Khan, A.A.; Akanda, A.M.; Mian, I.H.; Alam, M.Z. Chemical Control of Bacterial Soft Rot of Onion Caused by Burkholderia cepacia. J. Plant Pathol. 2013, 29, 1–4. [Google Scholar]
- Gardan, L.; Gouy, C.; Christen, R.; Samson, R. Elevation of three subspecies of Pectobacterium carotovorum to species level: Pectobacterium atrosepticum sp. nov., Pectobacterium betavasculorum sp. nov. and Pectobacterium wasabiae sp. nov. Int. J. Syst. Evol. Microbiol. 2003, 53, 381–391. [Google Scholar]
- Behiry, S.I.; Ashmawy, N.A.; Abdelkhalek, A.A.; Younes, H.A.; Khaled, A.E.; Hafez, E.E. Compatible- and incompatible-type interactions related to defense genes in potato elucidation by Pectobacterium carotovorum. J. Plant Dis. Prot. 2018, 125, 197–204. [Google Scholar] [CrossRef]
- Abo-Zaid, G.; Abdelkhalek, A.; Matar, S.; Darwish, M.; Abdel-Gayed, M. Application of Bio-Friendly Formulations of Chitinase-Producing Streptomyces cellulosae Actino 48 for Controlling Peanut Soil-Borne Diseases Caused by Sclerotium rolfsii. J. Fungi 2021, 7, 167. [Google Scholar]
- Abd El-Rahim, W.M.; Moawad, H.; Khalafallah, M. Enhancing the growth of promising fungal strains for rapid dye removal. Fresenius Environ. Bull. 2003, 12, 764–770. [Google Scholar]
- Abd El-Rahim, W.M.; Mostafa, E.M.; Moawad, H. High cell density cultivation of six fungal strains efficient in azo dye bioremediation. Biotechnol. Rep. 2016, 12, 1–5. [Google Scholar]
- Toth, I.K.; Van Der Wolf, J.M.; Saddler, G.; Lojkowska, E.; Hélias, V.; Pirhonen, M.; Tsror, L.; Elphinstone, J.G. Dickeya species: An emerging problem for potato production in Europe. Plant Pathol. 2011, 60, 385–399. [Google Scholar]
- Czajkowski, R.; Pérombelon, M.C.M.; Jafra, S.; Lojkowska, E.; Potrykus, M.; Van Der Wolf, J.M.; Sledz, W. Detection, identification and differentiation of Pectobacterium and Dickeya species causing potato blackleg and tuber soft rot: A review. Ann. Appl. Biol. 2015, 166, 18–38. [Google Scholar]
- Ji, P.; Momol, M.T.; Olson, S.M.; Pradhanang, P.M.; Jones, J.B. Evaluation of thymol as biofumigant for control of bacterial wilt of tomato under field conditions. Plant Dis. 2005, 89, 497–500. [Google Scholar]
- Williamson, L.; Nakaho, K.; Hudelson, B.; Allen, C. Ralstonia solanacearum race 3, biovar 2 strains isolated from geranium are pathogenic on potato. Plant Dis. 2002, 86, 987–991. [Google Scholar]
- Schaad, N.W.; Chun, W. Laboratory Guide for Identification of Plant Pathogenic Bacteria, 3rd ed.; American Phytopathological Society (APS Press): St. Paul, MN, USA, 2001. [Google Scholar]
- Farrar, J.; Nunez, J.; Davis, R. Losses due to lenticel rot are an increasing concern for Kern County potato growers. Calif. Agric. 2009, 63, 127–130. [Google Scholar]
- Bokshi, A.I.; Morris, S.C.; Deverall, B.J. Effects of benzothiadiazole and acetylsalicylic acid on β-1, 3-glucanase activity and disease resistance in potato. Plant Pathol. 2003, 52, 22–27. [Google Scholar]
- Olivier, C.; Halseth, D.E.; Mizubuti, E.S.G.; Loria, R. Postharvest application of organic and inorganic salts for suppression of silver scurf on potato tubers. Plant Dis. 1998, 82, 213–217. [Google Scholar]
- Krebs, H.; Jaggir, W. Effect of plant extracts against soft rot of potatoes: Erwiniacarotovora Flora and Fauna n Industrial Crops. Agrarforschung 1999, 6, 17–20. [Google Scholar]
- Simeon, A.U.; Abubakar, A. Evaluation of some plant extracts for the control of bacterial soft rot of tubers. Am. J. Exp. Agric. 2014, 4, 1869–1876. [Google Scholar]
- Iwu, M.W.; Duncan, A.R.; Okunji, C.O. New Antimicrobials of Plant Origin; Perspectives on New Crops and New Uses; ASHS Press: Alexandria, VA, USA, 1999; Volume 457, p. 462. [Google Scholar]
- Purushotham, S.P.; Anupama, N. In vitro antimicrobial screening of medicinal plants against clinical and phytopathogenic bacteria and fungi. Int. J. Pharm. Sci. Res. 2018, 9, 3005–3014. [Google Scholar]
- da Silva, C.M.A.; da Silva Costa, B.M.; da Silva, A.G.; de Souza, E.B.; dos Santos Correia, M.T.; de Menezes, L.V.L. Antimicrobial activity of several Brazilian medicinal plants against phytopathogenic bacteria. Afr. J. Microbiol. Res. 2016, 10, 578–583. [Google Scholar]
- Taha, A.S.; Elgat, W.A.A.; Fares, Y.G.D.; Dessoky, E.S.; Behiry, S.I.; Salem, M.Z.M. Using Plant Extractives as Eco-friendly Pulp Additives: Mechanical and Antifungal Properties of Paper Sheets Made from Linen Fibers. BioResources 2021, 16, 2589. [Google Scholar]
- Horváth, G.; Szabó, L.G.; Lemberkovics, É.; Botz, L.; Kocsis, B. Characterization and TLC-bioautographic detection of essential oils from some Thymus taxa. Determination of the activity of the oils and their components against plant pathogenic bacteria. JPC J. Planar Chromatogr. TLC 2004, 17, 300–304. [Google Scholar]
- Ashmawy, N.A.; Salem, M.Z.M.; El Shanhorey, N.; Al-Huqail, A.; Ali, H.M.; Behiry, S.I. Eco-friendly wood-biofungicidal and antibacterial activities of various Coccoloba uvifera L. leaf extracts: HPLC analysis of phenolic and flavonoid compounds. BioResources 2020, 15, 4165–4187. [Google Scholar]
- Okla, M.K.; Alamri, S.A.; Salem, M.Z.M.; Ali, H.M.; Behiry, S.I.; Nasser, R.A.; Alaraidh, I.A.; Al-Ghtani, S.M.; Soufan, W. Yield, phytochemical constituents, and antibacterial activity of essential oils from the leaves/twigs, branches, branch wood, and branch bark of Sour Orange (Citrus aurantium L.). Processes 2019, 7, 363. [Google Scholar]
- Behiry, S.I.; El-Hefny, M.; Salem, M.Z.M. Toxicity effects of Eriocephalus africanus L. leaf essential oil against some molecularly identified phytopathogenic bacterial strains. Nat. Prod. Res. 2020, 34, 3394–3398. [Google Scholar]
- Behiry, S.I.; Soliman, S.A.; Al-Askar, A.A.; Alotibi, F.O.; Basile, A.; Abdelkhalek, A.; Elsharkawy, M.M.; Salem, M.Z.M.; Hafez, E.E.; Heflish, A.A. Plantago lagopus extract as a green fungicide induces systemic resistance against Rhizoctonia root rot disease in tomato plants. Front. Plant Sci. 2022, 13, 966929. [Google Scholar] [CrossRef]
- Al-Askar, A.A.; Rashad, Y.M. Efficacy of some plant extracts against Rhizoctonia solani on pea. J. Plant Prot. Res. 2010, 50, 239–243. [Google Scholar] [CrossRef]
- Summerell, B.A.; Laurence, M.H.; Liew, E.C.Y.; Leslie, J.F. Biogeography and phylogeography of Fusarium: A review. Fungal Divers. 2010, 44, 3–13. [Google Scholar]
- Enya, J.; Togawa, M.; Takeuchi, T.; Yoshida, S.; Tsushima, S.; Arie, T.; Sakai, T. Biological and phylogenetic characterization of Fusarium oxysporum complex, which causes yellows on Brassica spp., and proposal of F. oxysporum f. sp. rapae, a novel forma specialis pathogenic on B. rapa in Japan. Phytopathology 2008, 98, 475–483. [Google Scholar]
- Garcia, B.; Grajales, A.; Cardenas, M.E.; Sierra, R.; Cepero, d.G.; Bernal, A.; Jimenez, P.; Restrepo, S. First report of Fusarium oxysporum causing potato dry rot in Solanum tuberosum in Colombia. New Dis. Reports 2011, 24, 14. [Google Scholar]
- Salem, M.Z.M.; Behiry, S.I.; EL-Hefny, M. Inhibition of Fusarium culmorum, Penicillium chrysogenum and Rhizoctonia solani by n-hexane extracts of three plant species as a wood-treated oil fungicide. J. Appl. Microbiol. 2019, 126, 1683–1699. [Google Scholar]
- Youssef, N.H.; Qari, S.H.; Matar, S.; Hamad, N.A.; Dessoky, E.S.; Elshaer, M.M.; Sobhy, S.; Abdelkhalek, A.; Zakaria, H.M.; Heflish, A.A. Licorice, Doum, and Banana Peel Extracts Inhibit Aspergillus flavus Growth and Suppress Metabolic Pathway of Aflatoxin B1 Production. Agronomy 2021, 11, 1587. [Google Scholar]
- Abdelkhalek, A.; Al-Askar, A.A.; Alsubaie, M.M.; Behiry, S.I. First Report of Protective Activity of Paronychia argentea Extract against Tobacco Mosaic Virus Infection. Plants 2021, 10, 2435. [Google Scholar] [PubMed]
- Behiry, S.I.; Mohamed, A.A.; Younes, H.A.; Salem, M.Z.M.; Salem, A.Z.M. Antigenic and pathogenicity activities of Ralstonia solanacearum race 3 biovar 2 molecularly identified and detected by indirect ELISA using polyclonal antibodies generated in rabbits. Microb. Pathog. 2018, 115, 216–221. [Google Scholar] [PubMed]
- Ashmawy, N.A.; Behiry, S.I.; Al-Huqail, A.A.; Ali, H.M.; Salem, M.Z.M. Bioactivity of Selected Phenolic Acids and Hexane Extracts from Bougainvilla spectabilis and Citharexylum spinosum on the Growth of Pectobacterium carotovorum and Dickeya solani Bacteria: An Opportunity to Save the Environment. Processes 2020, 8, 482. [Google Scholar]
- Salem, M.Z.M.; Behiry, S.I.; Salem, A.Z.M. Effectiveness of root-bark extract from Salvadora persica against the growth of certain molecularly identified pathogenic bacteria. Microb. Pathog. 2018, 117, 320–326. [Google Scholar]
- Aktaruzzaman, M.; Xu, S.-J.; Kim, J.-Y.; Woo, J.-H.; Hahm, Y.-I.; Kim, B.-S. First report of potato stem-end rot caused by Fusarium oxysporum in Korea. Mycobiology 2014, 42, 206–209. [Google Scholar]
- Dubey, S.C.; Tripathi, A.; Upadhyay, B.K.; Deka, U.K. Diversity of Rhizoctonia solani associated with pulse crops in different agro-ecological regions of India. World J. Microbiol. Biotechnol. 2014, 30, 1699–1715. [Google Scholar]
- Booth, C. Fusarium oxysporum CMI Descriptions of Pathogenic Fungi and Bacteria; Common Wealth Agricultural Bureaux: London, UK, 1970. [Google Scholar]
- Misawa, T.; Kurose, D.; Mori, M.; Toda, T. Characterization of Japanese Rhizoctonia solani AG-2-1 isolates using rDNA-ITS sequences, culture morphology, and growth temperature. J. Gen. Plant Pathol. 2018, 84, 387–394. [Google Scholar]
- Möller, E.M.; Bahnweg, G.; Sandermann, H.; Geiger, H.H. A simple and efficient protocol for isolation of high molecular weight DNA from filamentous fungi, fruit bodies, and infected plant tissues. Nucleic Acids Res. 1992, 20, 6115. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. Guid. Methods Appl. 1990, 18, 315–322. [Google Scholar]
- Nishizawa, T.; Komatsuzaki, M.; Sato, Y.; Kaneko, N.; Ohta, H. Molecular characterization of fungal communities in non-tilled, cover-cropped upland rice field soils. Microbes Environ. 2010, 25, 204–210. [Google Scholar]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- NCCLS. Approved Standards M2-A6 Performance Standards for Antimicrobial Disk Susceptibility Tests; National Committee for Clinical Laboratory Standards: Pittsburgh, PA, USA, 1997; ISBN 1562385860. [Google Scholar]
- Kumar, A.; Shukla, R.; Singh, P.; Prasad, C.S.; Dubey, N.K. Assessment of Thymus vulgaris L. essential oil as a safe botanical preservative against post harvest fungal infestation of food commodities. Innov. Food Sci. Emerg. Technol. 2008, 9, 575–580. [Google Scholar]
- Dissanayake, M. Inhibitory effect of selected medicinal plant extracts on phytopathogenic fungus Fusarium oxysporum (Nectriaceae) Schlecht. Emend. Snyder and Hansen. Annu. Res. Rev. Biol. 2014, 4, 133–142. [Google Scholar]
- Shimada, K.; Fujikawa, K.; Yahara, K.; Nakamura, T. Antioxidative properties of xanthan on the autoxidation of soybean oil in cyclodextrin emulsion. J. Agric. Food Chem. 1992, 40, 945–948. [Google Scholar]
- Abdelkhalek, A.; Király, L.; Al-Mansori, A.N.A.; Younes, H.A.; Zeid, A.; Elsharkawy, M.M.; Behiry, S.I. Defense Responses and Metabolic Changes Involving Phenylpropanoid Pathway and PR Genes in Squash (Cucurbita pepo L.) following Cucumber mosaic virus Infection. Plants 2022, 11, 1908. [Google Scholar] [CrossRef]
- Kausar, F.; Intisar, A.; Din, M.I.; Aamir, A.; Hussain, T.; Aziz, P.; Mutahir, Z.; Fareed, S.; Samreen, B.; Sadaqat, K. Volatile Composition and Antibacterial Activity of Leaves of Chorisia speciosa. J. Mex. Chem. Soc. 2020, 64, 339–348. [Google Scholar]
- El Sawi, S.A.M.; Hanafy, D.M.M.M.; El Alfy, T.S.M.A. Composition of the non-polar extracts and antimicrobial activity of Chorisia insignis HBK. leaves. Asian Pacific J. Trop. Dis. 2014, 4, 473–479. [Google Scholar]
- Brogden, R.N.; Speight, T.M.; Avery, G.S. Amoxycillin: A review of its antibacterial and pharmacokinetic properties and therapeutic use. Drugs 1975, 9, 88–140. [Google Scholar]
- Waxman, D.J.; Strominger, J.L. Penicillin-binding proteins and the mechanism of action of beta-lactam antibiotics. Annu. Rev. Biochem. 1983, 52, 825–869. [Google Scholar]
- Karaman, R. From conventional prodrugs to prodrugs designed by molecular orbital methods. In Frontiers in Computational Chemistry; Elsevier: Amsterdam, The Netherlands, 2015; pp. 187–249. [Google Scholar]
- Sneh, B.; Jabaji-Hare, S.; Neate, S.M.; Dijst, G. Rhizoctonia species: Taxonomy, Molecular Biology, Ecology, Pathology and Disease Control; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2013; ISBN 9401729018. [Google Scholar]
- Leslie, J.F.; Summerell, B.A. The Fusarium Laboratory Manual; John Wiley & Sons: Hoboken, NJ, USA, 2007; ISBN 0813819199. [Google Scholar]
- Singh, R.P.; Singh, P.K.; Rutkoski, J.; Hodson, D.P.; He, X.; Jørgensen, L.N.; Hovmøller, M.S.; Huerta-Espino, J. Disease impact on wheat yield potential and prospects of genetic control. Annu. Rev. Phytopathol. 2016, 54, 303–322. [Google Scholar]
- Hof, H. Critical annotations to the use of azole antifungals for plant protection. Antimicrob. Agents Chemother. 2001, 45, 2987–2990. [Google Scholar]
- Al-Huqail, A.A.; Behiry, S.I.; Salem, M.Z.M.; Ali, H.M.; Siddiqui, M.H.; Salem, A.Z.M. Antifungal, antibacterial, and antioxidant activities of Acacia saligna (Labill.) HL Wendl. flower extract: HPLC analysis of phenolic and flavonoid compounds. Molecules 2019, 24, 700. [Google Scholar]
- Jat, J.G.; Agalave, H.R. Fungitoxic properties of some leaf extracts against oilseed-borne fungi. Sci. Res. Rep. 2013, 3, 210–215. [Google Scholar]
- Ambikapathy, V.; Gomathi, S.; Panneerselvam, A. Effect of antifungal activity of some medicinal plants against Pythium debaryanum (Hesse). Asian J. Plant Sci. Res. 2011, 1, 131–134. [Google Scholar]
- Behbahani, B.A.; Shahidi, F.; Yazdi, F.T.; Mohebbi, M. Antifungal effect of aqueous and ethanolic mangrove plant extract on pathogenic fungus “in vitro”. Int. J. Agron. Plant Prod. 2013, 4, 1652–1658. [Google Scholar]
- Ashraf, Z.; Muhammad, A.; Imran, M.; Tareq, A.H. In vitro antibacterial and antifungal activity of methanol, chloroform and aqueous extracts of Origanum vulgare and their comparative analysis. Int. J. Org. Chem. 2011, 1, 257–261. [Google Scholar]
- Moorthy, K.K.; Subramaniam, P.; Senguttuvan, J. In vitro antifungal activity of various extracts of leaf and stem parts of Solena amplexicaulis (Lam.) Gandhi. Int. J. Pharm. Pharm. Sci. 2013, 5, 745–747. [Google Scholar]
- Heim, K.E.; Tagliaferro, A.R.; Bobilya, D.J. Flavonoid antioxidants: Chemistry, metabolism and structure-activity relationships. J. Nutr. Biochem. 2002, 13, 572–584. [Google Scholar]
- Procházková, D.; Boušová, I.; Wilhelmová, N. Antioxidant and prooxidant properties of flavonoids. Fitoterapia 2011, 82, 513–523. [Google Scholar]
- Mani, J.S.; Johnson, J.B.; Steel, J.C.; Broszczak, D.A.; Neilsen, P.M.; Walsh, K.B.; Naiker, M. Natural product-derived phytochemicals as potential agents against coronaviruses: A review. Virus Res. 2020, 284, 197989. [Google Scholar]
- Shaygannia, E.; Bahmani, M.; Zamanzad, B.; Rafieian-Kopaei, M. A review study on Punica granatum L. J. Evid. Based. Complementary Altern. Med. 2016, 21, 221–227. [Google Scholar]
- Duda-Madej, A.; Stecko, J.; Sobieraj, J.; Szymańska, N.; Kozłowska, J. Naringenin and Its Derivatives—Health-Promoting Phytobiotic against Resistant Bacteria and Fungi in Humans. Antibiotics 2022, 11, 1628. [Google Scholar]
- Duda-Madej, A.; Kozłowska, J.; Krzyżek, P.; Anioł, M.; Seniuk, A.; Jermakow, K.; Dworniczek, E. Antimicrobial O-alkyl derivatives of naringenin and their oximes against multidrug-resistant bacteria. Molecules 2020, 25, 3642. [Google Scholar]
- Murti, Y. Biological Evaluation of Synthesized Naringenin Derivatives as Antimicrobial Agents. Anti-Infective Agents 2021, 19, 192–199. [Google Scholar]
- Vandeputte, O.M.; Kiendrebeogo, M.; Rasamiravaka, T.; Stevigny, C.; Duez, P.; Rajaonson, S.; Diallo, B.; Mol, A.; Baucher, M.; El Jaziri, M. The flavanone naringenin reduces the production of quorum sensing-controlled virulence factors in Pseudomonas aeruginosa PAO1. Microbiology 2011, 157, 2120–2132. [Google Scholar]
- Górniak, I.; Bartoszewski, R.; Króliczewski, J. Comprehensive review of antimicrobial activities of plant flavonoids. Phytochem. Rev. 2019, 18, 241–272. [Google Scholar]
- Soberón, J.R.; Sgariglia, M.A.; Torrez, J.A.C.; Aguilar, F.A.; Pero, E.J.I.; Sampietro, D.A.; de Luco, J.F.; Labadie, G.R. Antifungal activity and toxicity studies of flavanones isolated from Tessaria dodoneifolia aerial parts. Heliyon 2020, 6, e05174. [Google Scholar]
- Salazar-Aranda, R.; Granados-Guzmán, G.; Pérez-Meseguer, J.; González, G.M.; Waksman de Torres, N. Activity of polyphenolic compounds against Candida glabrata. Molecules 2015, 20, 17903–17912. [Google Scholar]
- Vikram, A.; Jayaprakasha, G.K.; Jesudhasan, P.R.; Pillai, S.D.; Patil, B.S. Suppression of bacterial cell–cell signalling, biofilm formation and type III secretion system by citrus flavonoids. J. Appl. Microbiol. 2010, 109, 515–527. [Google Scholar] [PubMed]
- Lou, Z.; Wang, H.; Zhu, S.; Ma, C.; Wang, Z. Antibacterial activity and mechanism of action of chlorogenic acid. J. Food Sci. 2011, 76, M398–M403. [Google Scholar] [PubMed]
- Sun, Y.; Yu, Y.-M.; Suo, H.-B.; Zhu, Y.-L.; Huang, H.; Hu, Y. Research on extracting technology of chlorogenic acid from honeysuckle. In Proceedings of the Advances in Applied Biotechnology: Proceedings of the 3rd International Conference on Applied Biotechnology (ICAB2016), Tianjin, China, 25–27 November 2016; Springer: Berlin/Heidelberg, Germany, 2018; pp. 811–822. [Google Scholar]
- Huang, Y.; Chen, H.; Zhou, X.; Wu, X.; Hu, E.; Jiang, Z. Inhibition effects of chlorogenic acid on benign prostatic hyperplasia in mice. Eur. J. Pharmacol. 2017, 809, 191–195. [Google Scholar] [PubMed]
- Li, G.; Wang, X.; Xu, Y.; Zhang, B.; Xia, X. Antimicrobial effect and mode of action of chlorogenic acid on Staphylococcus aureus. Eur. Food Res. Technol. 2014, 238, 589–596. [Google Scholar]
- Zheng, Y.; Liu, J.; Cao, M.L.; Deng, J.M.; Kou, J. Extrication process of chlorogenic acid in Crofton weed and antibacterial mechanism of chlorogenic acid on Escherichia coli. J. Environ. Biol. 2016, 37, 1049. [Google Scholar] [PubMed]
- Su, M.; Liu, F.; Luo, Z.; Wu, H.; Zhang, X.; Wang, D.; Zhu, Y.; Sun, Z.; Xu, W.; Miao, Y. The antibacterial activity and mechanism of chlorogenic acid against foodborne pathogen Pseudomonas aeruginosa. Foodborne Pathog. Dis. 2019, 16, 823–830. [Google Scholar]
- Qian, L.; Xiao, H.; Zhao, G.; He, B. Synthesis of modified guanidine-based polymers and their antimicrobial activities revealed by AFM and CLSM. ACS Appl. Mater. Interfaces 2011, 3, 1895–1901. [Google Scholar]
- Borges, A.; Ferreira, C.; Saavedra, M.J.; Simões, M. Antibacterial activity and mode of action of ferulic and gallic acids against pathogenic bacteria. Microb. Drug Resist. 2013, 19, 256–265. [Google Scholar]
- Behiry, S.I.; Philip, B.; Salem, M.Z.M.; Amer, M.A.; El-Samra, I.A.; Abdelkhalek, A.; Heflish, A. Urtica dioica and Dodonaea viscosa leaf extracts as eco-friendly bioagents against Alternaria alternata isolate TAA-05 from tomato plant. Sci. Rep. 2022, 12, 16468. [Google Scholar]
- El Sawi, S.; Moawad, D.; El Alfy, S. Activity of Chorisia insignis HBK. against larynx carcinoma and chemical investigation of its polar extracts. J. Appl. Sci. Res. 2012, 8, 5564–5571. [Google Scholar]
- Youssef, N.H.; Qari, S.H.; Behiry, S.I.; Dessoky, E.S.; El-Hallous, E.I.; Elshaer, M.M.; Kordy, A.; Maresca, V.; Abdelkhalek, A.; Heflish, A.A. Antimycotoxigenic Activity of Beetroot Extracts against Altenaria alternata Mycotoxins on Potato Crop. Appl. Sci. 2021, 11, 4239. [Google Scholar]
- Mohamed, A.A.; Behiry, S.I.; Ali, H.M.; EL-Hefny, M.; Salem, M.Z.M.; Ashmawy, N.A. Phytochemical compounds of branches from P. halepensis oily liquid extract and S. terebinthifolius essential oil and their potential antifungal activity. Processes 2020, 8, 330. [Google Scholar]
- Nazzaro, F.; Fratianni, F.; De Martino, L.; Coppola, R.; De Feo, V. Effect of essential oils on pathogenic bacteria. Pharmaceuticals 2013, 6, 1451–1474. [Google Scholar]
- Desbois, A.P.; Smith, V.J. Antibacterial free fatty acids: Activities, mechanisms of action and biotechnological potential. Appl. Microbiol. Biotechnol. 2010, 85, 1629–1642. [Google Scholar]
- Oke, F.; Aslim, B.; Ozturk, S.; Altundag, S. Essential oil composition, antimicrobial and antioxidant activities of Satureja cuneifolia Ten. Food Chem. 2009, 112, 874–879. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Value | Reference |
|---|---|---|
| HPLC Instrument | Agilent 1260 series (Agilent Technologies, GmbH, Boblingen, Germany) | [64] |
| Column | (Eclipse C18) dimensions: 4.6 mm (diameter) × 250 mm (length) | |
| Particle size: 5 μm | ||
| Mobile phase | H2O (A) and 0.05% CF3COOH in CH3CN (B) | |
| Flow rate | 0.9 mL/min | |
| Gradient program | In this stepwise program, the mobile phase composition changes abruptly at specific intervals to facilitate the separation of the sample components. From 0 to 5 min, the mobile phase contains 80% solvent A and 20% solvent B. From 5 to 8 min, the mobile phase changes to 60% solvent A and 40% solvent B. From 8 to 12 min, the mobile phase remains at 60% solvent A and 40% solvent B. From 12 to 16 min, the mobile phase returns to the initial condition of 82% solvent A and 18% solvent B. From 16 to 20 min, the mobile phase remains at 82% solvent A and 18% solvent B. | |
| Detector | Multi-wavelength type monitored at 280 nm | |
| Injection volume | 5 μL | |
| Column temperature | 40 °C | |
| Analyzed compounds (standards) | 17 common phenolic and flavonoid components: apigenin, caffeic acid, catechin, chlorogenic acid, cinnamic acid, coumaric acid, daidzein, ellagic acid, ferulic acid, gallic acid, methyl gallate, naringenin, pyro catechol, quercetin, rutin, syringic acid, vanillin |
| Parameter | Value |
|---|---|
| Equipment | Agilent 7000D |
| Column | 5% Diphenyl/95% Dimethylpolysiloxan column, packed with HP-5MS capillary column (30 m in length × 250 μm in diameter × 0.25 μm in thickness) |
| Carrier Gas | Pure helium gas (99.99%) |
| Flow Rate | 1 mL/min |
| Ionization Energy | 70 eV |
| Scan Time | 0.2 s |
| Fragment Range | 40 to 600 m/z |
| Injection Quantity | 1 μL (split ratio 10:1) |
| Injector Temperature | 250 °C (constant) |
| Column Oven Temperature | 50 °C for 3 min, raised at 10 °C per min up to 280 °C, and final temperature was increased to 300 °C for 10 min |
| Identification of phytochemicals | Based on the comparison of their retention time (min), peak area, peak height, and mass spectral patterns with those spectral databases of authentic compounds stored in the National Institute of Standards and Technology (NIST) library |
| Treatments | Inhibition Zone (mm) | ||||
|---|---|---|---|---|---|
| Conc. (µg/mL) | Ralstonia solanacearum | Dickeya solani | Pectobacterium atrosepticum | Pectobacterium carotovorum | |
| Silk floss bark extract | 100 | 7.33 ± 0.24 d | 7.67 ± 0.47 c | 7.33 ± 0.24 d | 7.33 ± 0.24 b |
| 200 | 7.67 ± 0.94 cd | 7.67 ± 0.94 c | 7.33 ± 0.94 d | 7.67 ± 0.62 b | |
| 300 | 8.00 ± 0.82 cd | 8.67 ± 0.47 bc | 7.33 ± 0.24 d | 8.33 ± 1.25 b | |
| 400 | 8.67 ± 0.47 cd | 8.67 ± 1.18 bc | 7.33 ± 0.94 d | 8.33 ± 0.94 b | |
| 600 | 8.67 ± 1.70 cd | 8.67 ± 0.47 bc | 7.33 ± 0.47 d | 8.33 ± 0.47 b | |
| 800 | 9.33 ± 0.24 bcd | 9.00 ± 0.82 bc | 7.67 ± 0.47 cd | 8.67 ± 0.94 b | |
| 1000 | 10.33 ± 0.24 bc | 9.33 ± 0.47 b | 9.33 ± 0.47 bc | 9.00 ± 0.00 b | |
| 2000 | 12.00 ± 0.41 ab | 9.33 ± 0.47 b | 9.33 ± 0.94 bc | 9.00 ± 0.00 b | |
| 3000 | 14.33 ± 0.94 a | 9.67 ± 1.25 b | 9.67 ± 1.25 b | 9.67 ± 1.89 ab | |
| * Pc | 11.67 ± 0.85 ab | 13.00 ± 0.00 a | 14.33 ± 0.94 a | 12.67 ± 0.47 a | |
| ** Nc | 00.00 ± 0.00 e | 00.00 ± 0.00 d | 00.00 ± 0.00 e | 00.00 ± 0.00 c | |
| Treatments Conc. (µg/mL) | Inhibition Percentage (%) | |
|---|---|---|
| Rhizoctonia solani | Fusariumoxysporum | |
| 25 | 28.10 ± 4.71 d | 82.38 ± 1.78 c |
| 50 | 39.05 ± 2.94 c | 83.33 ± 0.67 bc |
| 100 | 41.90 ± 0.79 c | 86.19 ± 0.41 ab |
| 200 | 70.48 ± 1.35 b | 86.19 ± 0.67 ab |
| 300 | 83.33 ± 2.43 a | 86.67 ± 0.59 a |
| * Pc | 63.33 ± 0.76 b | 83.81 ± 0.44 abc |
| ** Nc | 00.00 ± 0.00 e | 00.00 ± 0.00 d |
| Compounds | Area | Concentration (µg/g) |
|---|---|---|
| Gallic acid | 173.90 | 812.34 |
| Chlorogenic acid | 358.17 | 2727.49 |
| Catechin | 23.36 | 324.42 |
| Methyl gallate | 166.08 | 651.73 |
| Caffeic acid | * ND | ND |
| Syringic acid | 166.16 | 946.26 |
| Pyro catechol | ND | ND |
| Rutin | 15.57 | 97.16 |
| Ellagic acid | 13.11 | 139.18 |
| Coumaric acid | 62.34 | 104.47 |
| Vanillin | 145.02 | 473.22 |
| Ferulic acid | 285.48 | 1276.18 |
| Naringenin | 2968.31 | 18,698.83 |
| Daidzein | 195.35 | 628.91 |
| Quercetin | 37.72 | 220.42 |
| Cinnamic acid | 13.59 | 22.46 |
| Apigenin | ND | ND |
| Kaempferol | ND | ND |
| Hesperetin | ND | ND |
| RT | Compound | Class | Relative Abundance % |
|---|---|---|---|
| 5.25 | Cyclohexene, 1-methyl-5-(1-methylethenyl)-, (R)- | Terpenoid | 1.29 |
| 13.75 | Benzoic acid 2-(methylamino)-methyl ester | Benzoic acid derivative | 1.32 |
| 17.96 | 1,2-benzenedicarboxylic acid diethyl ester | Phthalate ester | 0.73 |
| 23.77 | Isopropyl myristate | Fatty acid ester | 55.61 |
| 25.64 | Palmitic acid methyl ester | Fatty acid ester | 3.51 |
| 26.41 | n-Hexadecanoic acid | Fatty acid | 14.05 |
| 28.64 | 12,15-Octadecadienoic acid methyl ester | Fatty acid ester | 1.09 |
| 28.70 | Linoleic acid methyl ester | Fatty acid ester | 1.42 |
| 28.82 | 11-Octadecenoic acid methyl ester | Fatty acid ester | 1.23 |
| 29.36 | 9,12-Octadecadienoic acid (Z,Z)- | Fatty acid | 3.95 |
| 29.52 | Oleic acid | Fatty acid | 4.65 |
| 29.60 | cis-Vaccenic acid | Fatty acid | 1.55 |
| 30.00 | Octadecanoic acid | Fatty acid | 1.17 |
| 35.80 | Bis(2-ethylhexyl) phthalate | Phthalate ester | 8.45 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al-Askar, A.A. Phytochemical Properties of Silk Floss Tree Stem Bark Extract and Its Potential as an Eco-Friendly Biocontrol Agent against Potato Phytopathogenic Microorganisms. Horticulturae 2023, 9, 912. https://doi.org/10.3390/horticulturae9080912
Al-Askar AA. Phytochemical Properties of Silk Floss Tree Stem Bark Extract and Its Potential as an Eco-Friendly Biocontrol Agent against Potato Phytopathogenic Microorganisms. Horticulturae. 2023; 9(8):912. https://doi.org/10.3390/horticulturae9080912
Chicago/Turabian StyleAl-Askar, Abdulaziz A. 2023. "Phytochemical Properties of Silk Floss Tree Stem Bark Extract and Its Potential as an Eco-Friendly Biocontrol Agent against Potato Phytopathogenic Microorganisms" Horticulturae 9, no. 8: 912. https://doi.org/10.3390/horticulturae9080912