
Effects of Extreme Root Restriction on the Nutritional and Flavor Quality, and Sucrose Metabolism of Tomato (Solanum lycopersicum L.)

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Culture Conditions
2.2. Measurement of Plant Growth and Fruit Production
2.3. Analysis of Metabolites in Tomato Fruits
2.4. Analysis of Enzyme Activity
2.5. Quantitative Real-Time PCR Analysis
2.6. Statistical Analysis
3. Results
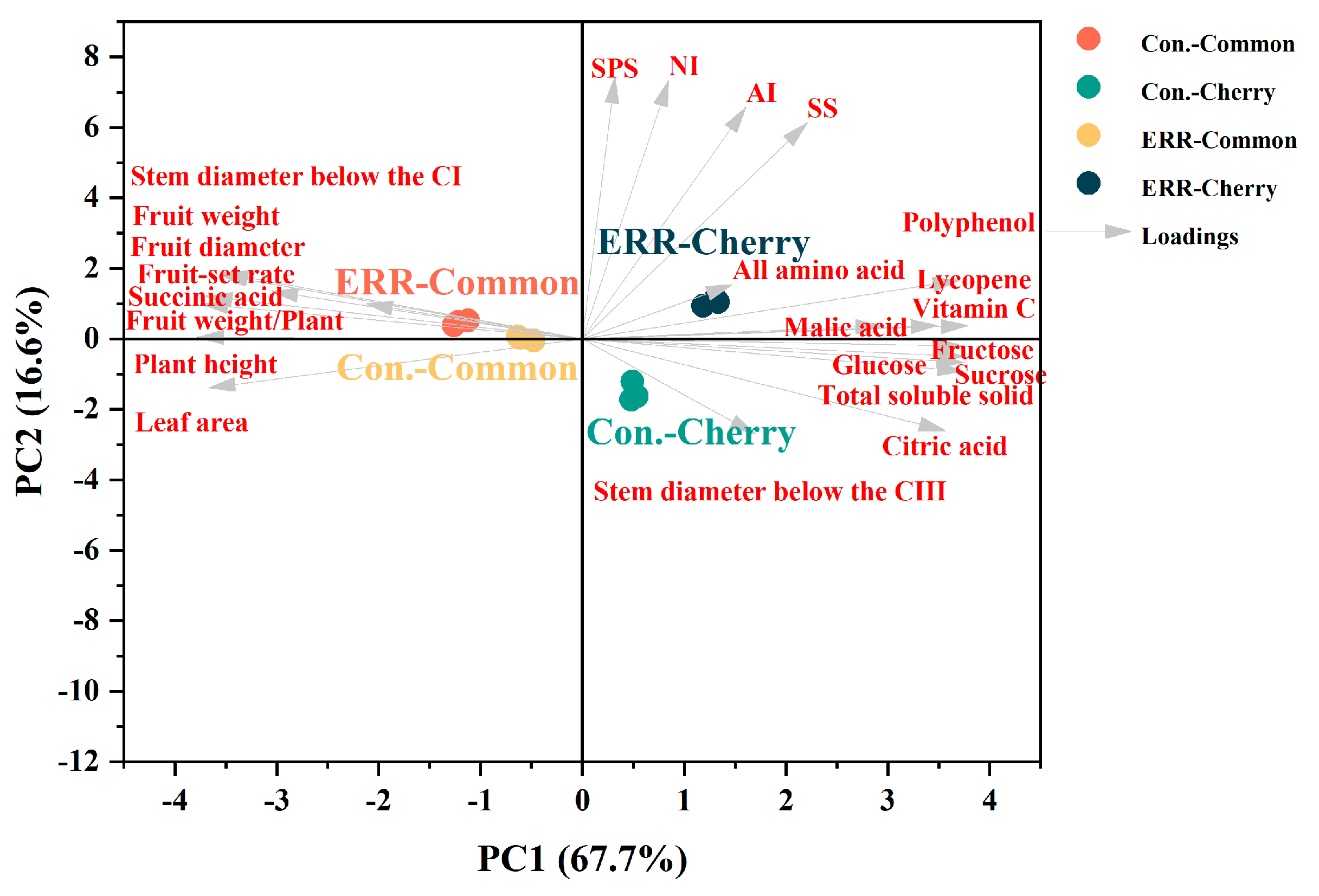
3.1. Plant Growth and Fruit Production
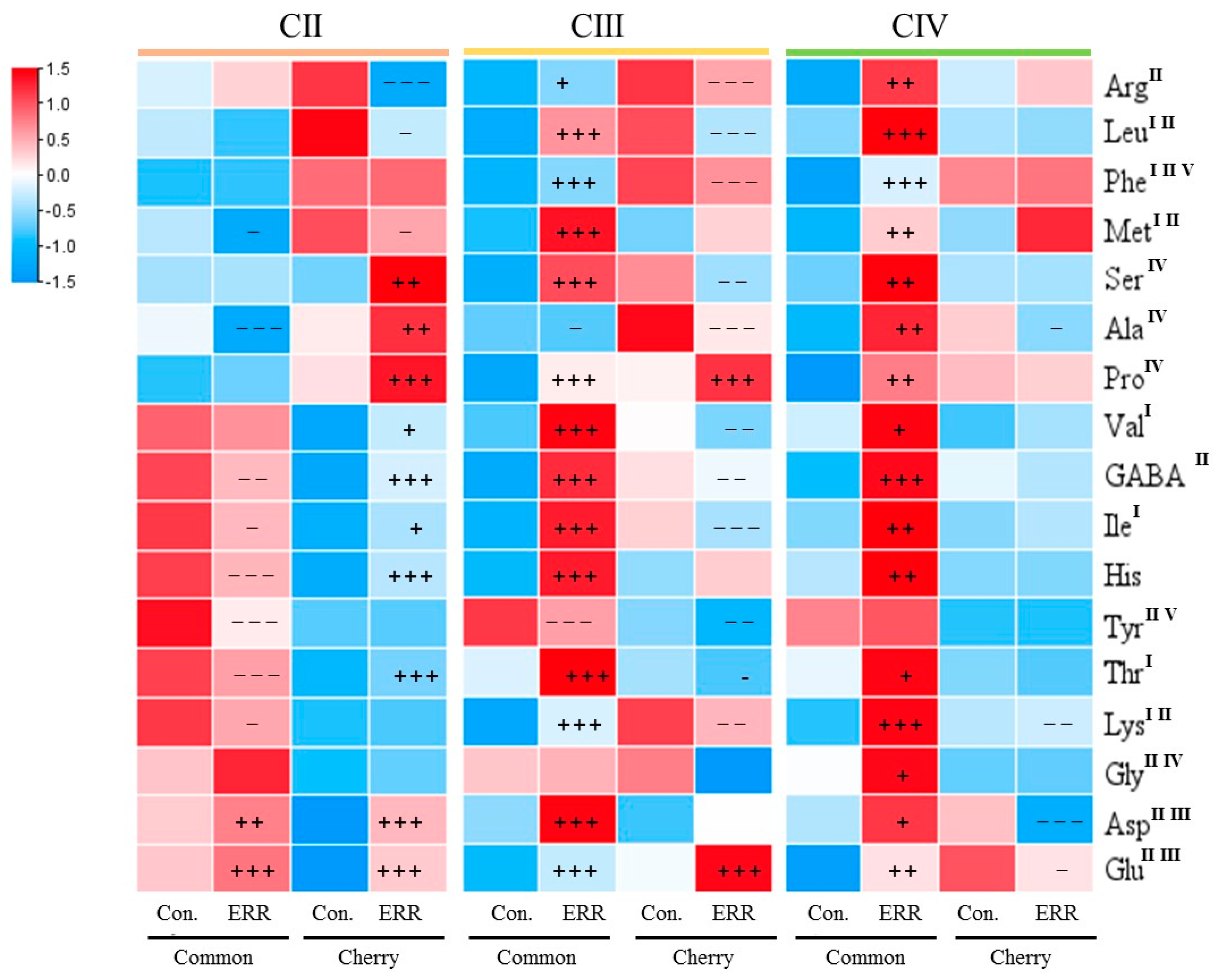
3.2. Nutritional and Flavor Quality of Tomatoes
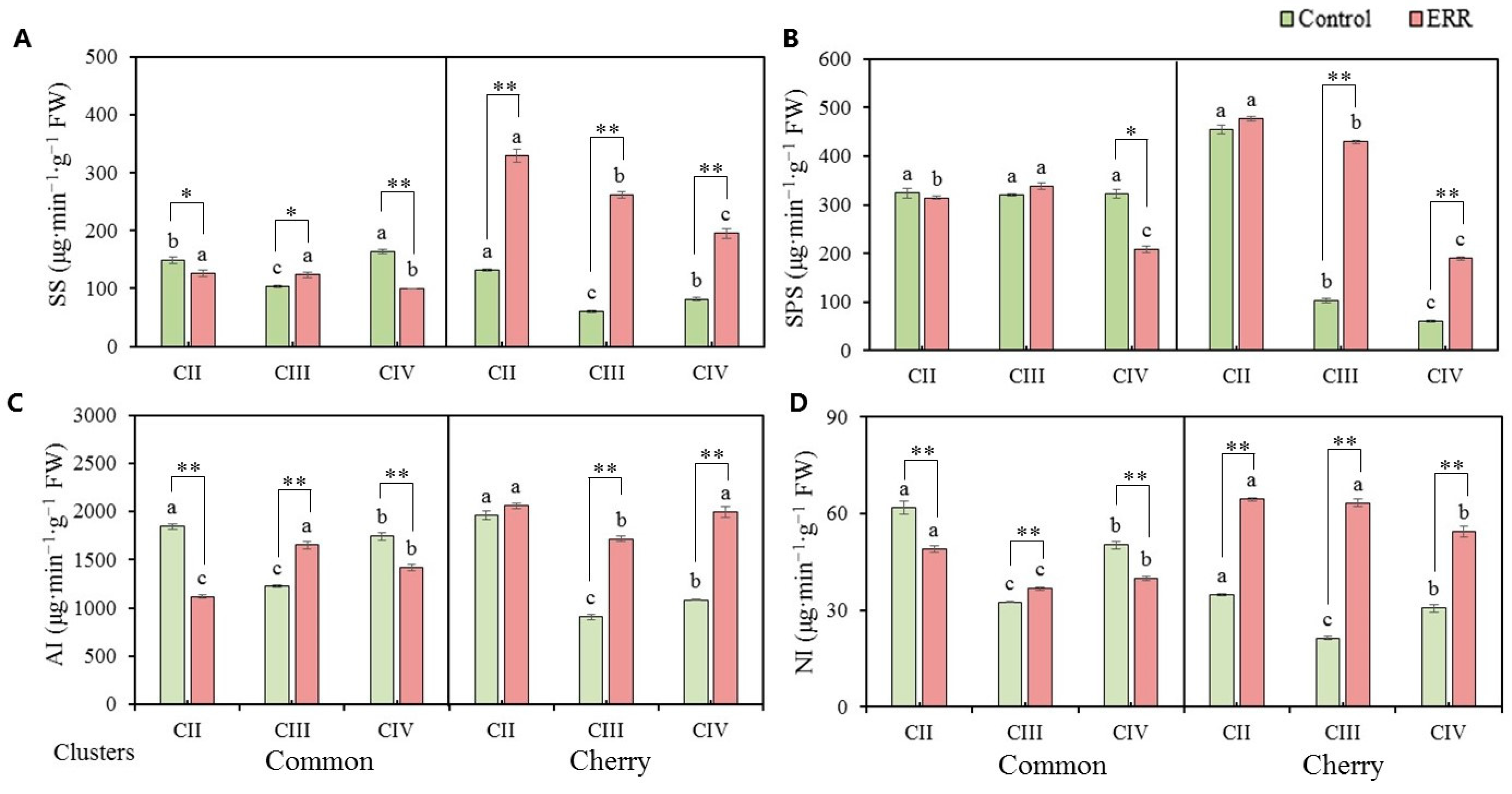
3.3. Activity of Sucrose Metabolism Associated Enzymes
3.4. Genes Expression Level
4. Discussion
4.1. ERR Suppressed Plant Growth and Fruit Size, but Improved Fruit Uniformity
4.2. ERR Improved Nutritional and Flavor Quality of Tomatoes
4.3. ERR Enhanced Enzyme Activity Involved Sucrose Metabolism in Tomato Fruits
4.4. ERR Induced the Expression of Sucrose Metabolism Genes in Tomato Fruits
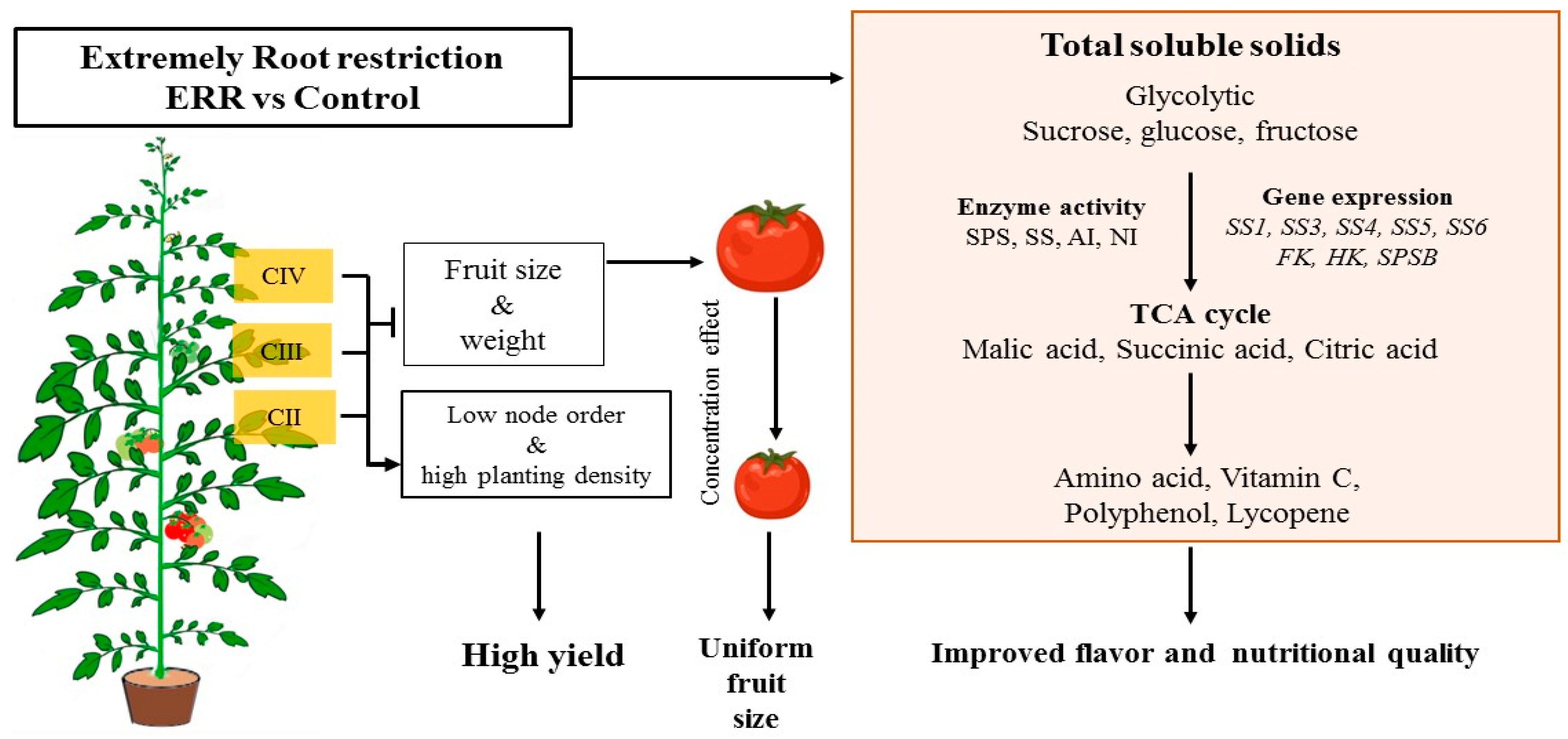
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- FAOSTAT. 2019. Available online: http://www.fao.org/faostat/en/#home (accessed on 3 July 2023).
- Li, T.L.; Xu, Y.; Zhang, J. Current situation and trend of industry development of facilities vegetable, western melon and edible fungus. China Veg. 2019, 11, 6–9. [Google Scholar]
- Zhu, G.; Wang, S.; Huang, Z.; Zhang, S.; Liao, Q.; Zhang, C.; Lin, T.; Qin, M.; Peng, M.; Yang, C.; et al. Rewiring of the fruit metabolome in tomato breeding. Cell 2018, 172, 249–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ripoll, J.; Urban, L.; Brunel, B.; Bertin, N. Water deficit effects on tomato quality depend on fruit developmental stage and genotype. J. Plant Physiol. 2016, 190, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Kyriacou, M.C.; Rouphael, Y. Towards a new definition of quality for fresh fruits and vegetables. Sci. Hortic. 2018, 234, 463–469. [Google Scholar] [CrossRef]
- Ayarna, A.W.; Tsukagoshi, S.; Nkansah, G.O. Effect of root restriction on the performance of three-truss-cultivated tomato in the low node pinching order at high-density cultivation system. Horticulturae 2021, 7, 60. [Google Scholar] [CrossRef]
- Rouphael, Y.; Kyriacou, M.C.; Petropoulos, S.; Pascale, S.; Colla, G. Improving vegetable quality in controlled environments. Sci. Hortic. 2018, 234, 275–289. [Google Scholar] [CrossRef]
- Zahid, M.S.; Hussain, M.; Song, Y.; Li, J.J.; Guo, D.H.; Li, X.Y.; Song, S.R.; Wang, L.; Xu, W.P.; Wang, S.P. Root-zone restriction regulates soil factors and bacterial community assembly of grapevine. Int. J. Mol. Sci. 2022, 23, 15628. [Google Scholar] [CrossRef]
- Yu, X.M.; Wang, B.; Zhang, C.X.; Xu, W.P.; He, J.J.; Zhu, L.; Wang, S.P. Effect of root restriction on nitrogen levels and glutamine synthetase activity in ‘Kyoho’ grapevines. Sci. Hortic. 2012, 137, 156–163. [Google Scholar] [CrossRef]
- Yu, X.M.; Li, J.F.; Zhu, L.N.; Wang, B.; Wang, L.; Bai, Y.; Zhang, C.X.; Xu, W.P.; Wang, S.P. Effects of root restriction on nitrogen and gene expression levels in nitrogen metabolism in ‘jumeigui’ grapevines (Vitis vinifera L. × Vitis labrusca L.). J. Integr. Agric. 2015, 14, 67–79. [Google Scholar] [CrossRef]
- Zakaria, N.I.; Ismail, M.R.; Awang, Y.; Megat Wahab, P.E.; Berahim, Z. Effect of root restriction on the growth, photosynthesis rate, and source and sink relationship of chilli (Capsicum annuum L.) grown in soilless culture. BioMed. Res. Int. 2020, 2020, 2706937. [Google Scholar] [CrossRef] [Green Version]
- Shi, K.; Fu, L.J.; Dong, D.K.; Zhou, Y.H.; Yu, J.Q. Decreased energy synthesis is partially compensated by a switch to sucrose synthase pathway of sucrose degradation in restricted root of tomato plants. Plant Physiol. Biochem. 2008, 46, 1040–1044. [Google Scholar] [CrossRef]
- Li, X.J.; Han, Z.; Li, C.; Xie, Z.S.; Wang, S.P.; Li, B. Effects of root restriction on soluble sugar contents and ultrastructure of phloem tissues in ‘kyoho’ grape berry. Plant Physiol. 2016, 52, 1546–1554. [Google Scholar]
- White, M.D.; Tustin, D.S.; Foote, K.F.; Campbell, J.M. Growth of young sweet cherry trees in response to root restriction using root control bags. Acta Hortic. 2001, 557, 391–398. [Google Scholar] [CrossRef]
- Tamai, D. The practical cultivation and technologist training in tomato low node order pinching and high density planting cultivation. Shisetsu Engei 2014, 165, 62–65. (In Japanese) [Google Scholar]
- Zhang, Y.T.; Kiriiwa, Y.; Nukaya, A. Influence of nutrient concentration and composition on the growth, uptake patterns of nutrient elements and fruit coloring disorder for tomatoes grown in extremely low-volume substrate. Hortic. J. 2015, 84, 37–45. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.T.; Suzuki, K.; Liu, H.C.; Nukaya, A.; Kiriiwa, Y. Fruit yellow-shoulder disorder as related to mineral element uptake of tomatoes grown in high temperature. Sci. Hortic. 2018, 242, 25–29. [Google Scholar] [CrossRef]
- Liu, A.L.; Liu, D.X.; Yin, D.S.; Lian, H.; Zhang, Y.T.; Chen, R.Y. Effects of minimal drainage and nutrient concentration regulation on tomato fruit quality. China Veg. 2021, 10, 66–78, (In Chinese with English Abstract). [Google Scholar]
- Wang, H.C.; Huang, H.B.; Huang, X.M.; Hu, Z.Q. Sugar and acid compositions in the arils of litchi chinensis sonn.: Cultivar differences and evidence for the absence of succinic acid. J. Hortic. Sci. Biotechnol. 2006, 81, 57–62. [Google Scholar] [CrossRef]
- He, R.; Wei, J.J.; Zhang, J.Y.; Tan, X.; Li, Y.M.; Gao, M.F.; Liu, H.C. Supplemental blue light frequencies improve ripening and nutritional qualities of tomato fruits. Front. Plant Sci. 2022, 13, 888976. [Google Scholar] [CrossRef]
- Wang, W.; Liu, D.X.; Qin, M.; Xie, Z.B.; Chen, R.Y.; Zhang, Y.T. Effects of supplemental lighting on potassium transport and fruit coloring of tomatoes grown in hydroponics. Int. J. Mol. Sci. 2021, 22, 2687. [Google Scholar] [CrossRef]
- Zhang, Y.T.; Liu, A.L.; Hao, Y.W.; Su, W.; Sun, G.W.; Song, S.W.; Liu, H.C.; Chen, R.Y. Nitric oxide is essential for melatonin to enhance nitrate tolerance of cucumber seedlings. Molecules 2022, 27, 5806. [Google Scholar] [CrossRef]
- Xie, Z.S.; Li, B.; Forney, C.F.; Xu, W.P.; Wang, S.P. Changes in sugar content and relative enzyme activity in grape berry in response to root restriction. Sci. Hortic. 2009, 123, 39–45. [Google Scholar] [CrossRef]
- Saito, T.; Fukuda, N.; Iikubo, T.; Inai, S.; Fujii, T.; Konishi, C.; Eaura, H. Effects of root-volume restriction and salinity on the fruit yield and quality of processing tomato. J. Jap. Soc. Hortic. Sci. 2008, 77, 165–172. [Google Scholar] [CrossRef] [Green Version]
- Goto, T.; Matsuno, T.; Yoshida, Y.; Kageyama, Y. Photosynthetic, evapotranspiratory and leaf morphological properties of chrysanthemum grown under root restriction as affected by fertigation frequency. Engei Gakkai Zasshi 2002, 71, 277–283. [Google Scholar] [CrossRef]
- Leng, F.; Lin, Q.; Wu, D.; Wang, S.P.; Wang, D.L.; Sun, C.D. Comparative transcriptomic analysis of grape berry in response to root restriction during developmental stages. Molecules 2016, 21, 1431. [Google Scholar] [CrossRef] [Green Version]
- Zhu, L.C.; Li, B.Y.; Wu, L.M.; Li, H.X.; Wang, Z.Y.; Wei, X.Y.; Ma, B.Q.; Zhang, Y.F.; Ma, F.W.; Ruan, Y.L.; et al. MdERDL6-mediated glucose efflux to the cytosol promotes sugar accumulation in the vacuole through up-regulating TSTs in apple and tomato. Proc. Natl. Acad. Sci. USA 2021, 118, e2022788118. [Google Scholar] [CrossRef]
- Saito, T.; Matsukura, C.; Ban, Y.; Shoji, K.; Sugiyama, M.; Fukuda, N.; Nishimura, S. Salinity stress affects assimilate metabolism at the gene-expression level during fruit development and improves fruit quality in tomato. J. Jap. Soc. Hortic. Sci. 2008, 77, 61–68. [Google Scholar] [CrossRef] [Green Version]
- Singh, H.; Kumar, P.; Chaudhari, S.; Edelstein, M. Tomato grafting: A global perspective. Hortic. Sci. 2017, 52, 1328–1336. [Google Scholar] [CrossRef] [Green Version]
- Kumar, P.; Rouphael, Y.; Cardarelli, M.; Colla, G. Effect of nickel and grafting combination on yield, fruit quality, antioxidative enzyme activities, lipid peroxidation, and mineral composition of tomato. J. Plant Nutr. Soil Sci. 2015, 178, 848–860. [Google Scholar] [CrossRef]
- Cantero-Navarro, E.; Romero-Aranda, R.; Fernández-Muñoz, R.; Martínez-Andújar, C.; Pérez-Alfocea, F.; Albacete, A. Improving agronomic water use efficiency in tomato by rootstock-mediated hormonal regulation of leaf biomass. Plant Sci. 2016, 251, 90–100. [Google Scholar] [CrossRef] [PubMed]
- Albacete, A.; Martínez-Andújar, C.; Ghanem, M.E.; Acosta, M.; Sánchez-Bravo, J.; Asins, M.J.; Cuartero, J.; Lutts, S.; Dodd, I.C.; Pérez-Alfocea, F. Rootstock-mediated changes in xylem ionic and hormonal status are correlated with delayed leaf senescence, and increased leaf area and crop productivity in salinized tomato. Plant Cell Environ. 2009, 32, 928–938. [Google Scholar] [CrossRef]
- Kanayama, Y. Sugar metabolism and fruit development in the tomato. Hortic. J. 2017, 86, 417–425. [Google Scholar] [CrossRef] [Green Version]
- Nguyen-Quoc, B.; Foyer, C.H. A Role for ‘futile cycles’ involving invertase and sucrose synthase in sucrose metabolism of tomato fruit. J. Expt. Bot. 2001, 52, 881–889. [Google Scholar] [CrossRef] [Green Version]
- Figás, M.R.; Prohens, J.; Raigón, M.D.; Fita, A.; García-Martínez, M.D.; Casanova, C.; Borras, D.; Plazas, M.; Andújar, I.; Soler, S. Characterization of composition traits related to organoleptic and functional quality for the differentiation, selection and enhancement of local varieties of tomato from different cultivar groups. Food Chem. 2015, 187, 517–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, S.W.; Li, T.L.; Jiang, J. Tomato key sucrose metabolizing enzyme activities and gene expression under NaCl and PEG iso-osmotic stresses. Agric. Sci. China 2009, 8, 1046–1052. [Google Scholar] [CrossRef]
- Miron, D.; Schaffer, A.A. Sucrose phosphate synthase sucrose synthase, and invertase activities in developing fruit of lycopersicon esculentum mill. and the sucrose accumulating lycopersicon hirsutum humb. and Bonpl. Plant Physiol. 1991, 95, 623–627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, B.; He, J.J.; Duan, C.Q.; Yu, X.M.; Zhu, L.N.; Xie, Z.S.; Zhang, C.X.; Xu, W.P.; Wang, S.P. Root restriction affects anthocyanin accumulation and composition in berry skin of “kyoho” grape (Vitis vinifera L. × Vitis labrusca L.) during ripening. Sci. Hortic. 2012, 137, 20–28. [Google Scholar] [CrossRef]
- Li, D.M.; Liu, B.Y.; Wang, Z.P.; Li, X.Y.; Sun, S.J.; Ma, C.; Wang, L.; Wang, S.P. Sugar accumulation may be regulated by a transcriptional cascade of ABA-VvGRIP55-VvMYB15-VvSWEET15 in grape berries under root restriction. Plant Sci. 2022, 322, 111288. [Google Scholar] [CrossRef]
- Carrari, F.; Fernie, A.R. Metabolic regulation underlying tomato fruit development. J. Exp. Bot. 2006, 57, 1883–1897. [Google Scholar] [CrossRef]
- Daloso, D.M.; Williams, T.C.R.; Antunes, W.C.; Antunes, W.C.; Pinheiro, D.P.; Muller, C.; Loureiro, M.E.; Fernie, A.R. Guard cell-specific upregulation of sucrose synthase 3 reveals that the role of sucrose in stomatal function is primarily energetic. New Phytol. 2016, 209, 1470–1483. [Google Scholar] [CrossRef] [Green Version]
- Su, J.Y.; Yao, Y.; Liu, Y.H.; Han, Q.Y.; Zhang, W.L. Function, structure and catalytic mechanism of sucrose phosphate synthase, a review. Chin. J. Biotech. 2021, 37, 1858–1868. [Google Scholar]
- Yang, J.C.; Zhang, J.H.; Wang, Z.Q.; Zhu, Q.S. Activities of starch hydrolytic enzymes and sucrose-phosphate synthase in the stems of rice subjected to water stress during grain filling. J. Exp. Bot. 2001, 52, 2169–2179. [Google Scholar] [CrossRef] [PubMed]
- Claeyssen, E.; Rivoal, J. Isozymes of plant hexokinase: Occurrence, properties and functions. Phytochemistry 2007, 68, 709–731. [Google Scholar] [CrossRef] [PubMed]
- Eveland, A.L.; Jackson, D.P. Sugars, signalling, and plant development. J. Exp. Bot. 2012, 63, 3367–3377. [Google Scholar] [CrossRef] [Green Version]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, D.; Chen, J.; Hao, Y.; Yang, X.; Chen, R.; Zhang, Y. Effects of Extreme Root Restriction on the Nutritional and Flavor Quality, and Sucrose Metabolism of Tomato (Solanum lycopersicum L.). Horticulturae 2023, 9, 813. https://doi.org/10.3390/horticulturae9070813
Liu D, Chen J, Hao Y, Yang X, Chen R, Zhang Y. Effects of Extreme Root Restriction on the Nutritional and Flavor Quality, and Sucrose Metabolism of Tomato (Solanum lycopersicum L.). Horticulturae. 2023; 9(7):813. https://doi.org/10.3390/horticulturae9070813
Chicago/Turabian StyleLiu, Danxia, Jinxiang Chen, Yanwei Hao, Xiaolong Yang, Riyuan Chen, and Yiting Zhang. 2023. "Effects of Extreme Root Restriction on the Nutritional and Flavor Quality, and Sucrose Metabolism of Tomato (Solanum lycopersicum L.)" Horticulturae 9, no. 7: 813. https://doi.org/10.3390/horticulturae9070813