

Efficient Cold Tolerance Evaluation of Four Species of Liliaceae Plants through Cell Death Measurement and Lethal Temperature Prediction

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Plants
2.2. Cold Treatment
2.3. Visual Inspection of Cold-Treated Plants
2.4. Electrolyte Leakage and Prediction of Lethal Temperature
2.5. Measurement of Cell Death of Cold-Treated Plants
2.6. Determination of Reducing Sugar Contents
2.7. Selection of Cold-Tolerant Plants
2.8. Statistical Analysis
3. Results
3.1. Visual Inspection of Cold-Treated Plants
3.2. Measurement of Electrolyte Leakage Value
3.3. Prediction of Lethal Temperature
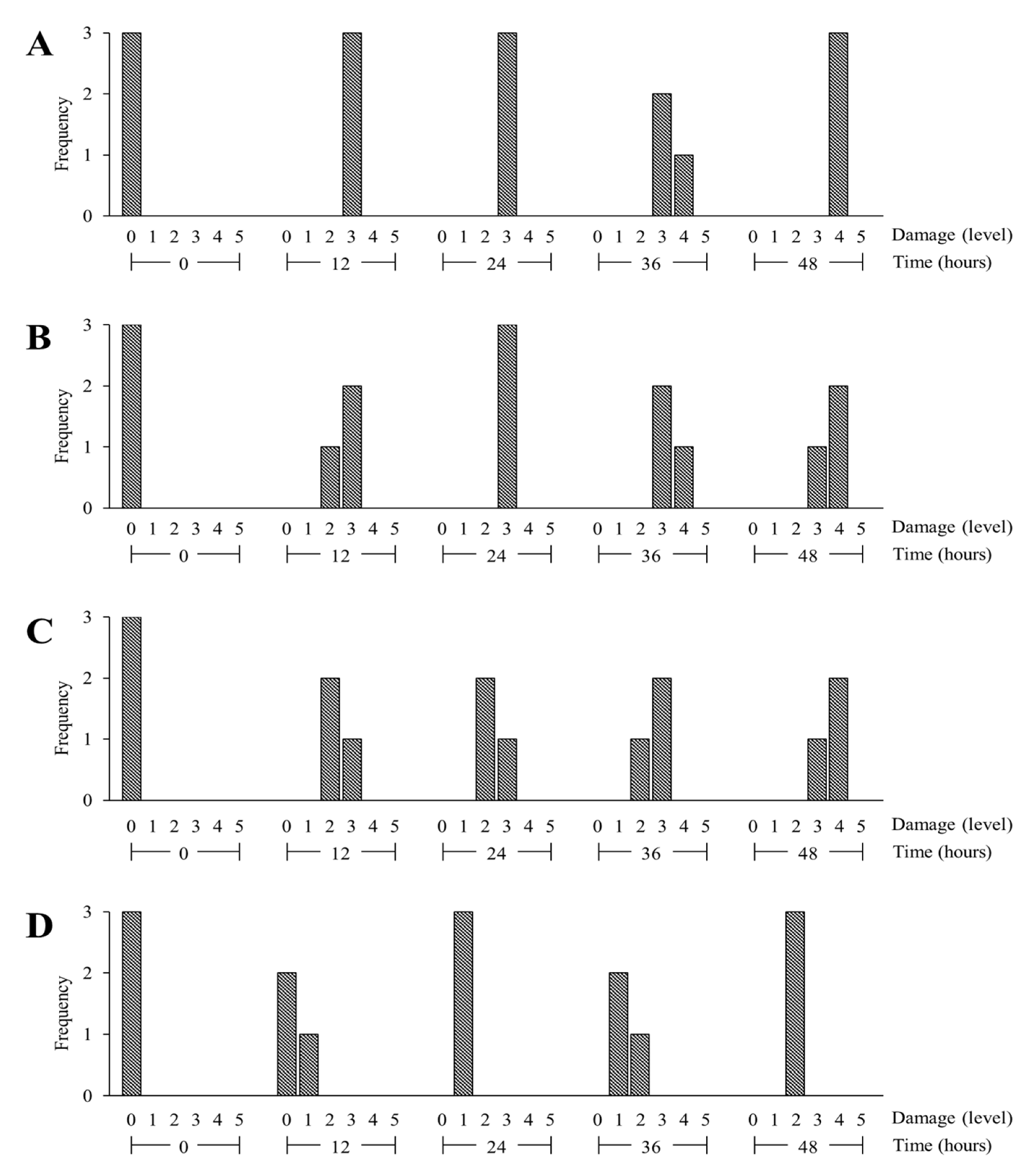
3.4. Determine the Degree of Cell Damage in Cold-Treated Plants
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, M.; Liao, W. Carbon monoxide as a signaling molecule in plants. Front. Plant Sci. 2016, 7, 572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zohner, C.M.; Renner, L.M.S.; Crowthe, T.W. Late-spring frost risk between 1959 and 2017 decreased in North America but increased in Europe and Asia. Proc. Natl. Acad. Sci. USA 2020, 117, 12192–12200. [Google Scholar] [CrossRef] [PubMed]
- Lichtenthaler, H.K. The stress concept in plants: An introduction. In Stress of Life: From Molecules to Man; Csermely, P., Ed.; New York Academy of Sciences: New York, NY, USA, 1998; Volume 851, pp. 187–198. [Google Scholar]
- Lee, S.Y.; Jung, J.A.; Sung, J.K.; Ha, S.K.; Lee, D.B.; Kim, T.W.; Song, B.H. Responses of nutrient uptake, carbohydrates and antioxidants against low temperature in plants. Korean J. Agric. Sci. 2014, 41, 75–83. [Google Scholar]
- Guo, X.; Liu, D.; Chong, K. Cold signaling in plants: Insights into mechanisms and regulation. J. Integr. Plant Biol. 2018, 60, 745–756. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Shi, Y.; Yang, S. Insights into the regulation of C-repeat binding factors in plant cold signaling. J. Integr. Plant Biol. 2018, 60, 780–795. [Google Scholar] [CrossRef] [Green Version]
- Murray, M.B.; Cape, J.N.; Fowler, D. Quantification of frost damage in plant tissues by rates of electrolyte leakage. New Phytol. 1989, 113, 307–311. [Google Scholar] [CrossRef]
- Kim, I.H. Development of Shallow-Extensive Green Roof System for Urban Greening. Ph.D. Thesis, Gyeongsang National University, Jinju, Republic of Korea, 2006. [Google Scholar]
- Dexter, S.T.; Tottingham, W.E.; Graber, L.F. Preliminary results in measuring the hardiness of plants. Plant Physiol. 1930, 5, 215–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, I.H.; Huh, K.Y.; Jung, H.J.; Choi, S.M.; Park, J.H. Modeling methodology for cold tolerance assessment of Pittosporum tobira. Korean J. Hortic. Sci. Technol. 2014, 32, 241–251. [Google Scholar] [CrossRef] [Green Version]
- Xie, H.; Sun, Y.; Cheng, B.; Xue, S.; Cheng, D.; Liu, L.; Meng, L.; Qiang, S. Variation in ICE1 methylation primarily determines phenotypic variation in freezing tolerance in Arabidopsis thaliana. Plant Cell Physiol. 2019, 60, 152–165. [Google Scholar] [CrossRef]
- Gonzalez-Mendoza, D.; Quiroz-Moreno, A.; Medrano, R.E.; Grimaldo-Juareza, O.; Zapata-Perezd, O. Cell viability and leakage of electrolytes in Avicennia germinans exposed to heavy metals. Z. Für Nat. C J. 2009, 64, 391–394. [Google Scholar] [CrossRef]
- Szalay, L.; Gyökös, I.G.; Békefi, Z. Cold hardiness of peach flowers at different phenological stages. Hortic. Sci. 2018, 45, 119–124. [Google Scholar] [CrossRef]
- Su, L.; Dai, Z.; Li, S.; Xin, H. A novel system for evaluating drought–cold tolerance of grapevines using chlorophyll fluorescence. BMC Plant Biol. 2015, 15, 82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindén, L. Measuring Cold Hardiness in Woody Plants. Ph.D. Thesis, Applied Biology, Horticulture, University of Helsinki, Helsinki, Finland, 2002. [Google Scholar]
- Du, Y.P.; He, H.B.; Wang, Z.X.; Wei, C.; Li, S.; Jia, G.X. Investigation and evaluation of the genus Lilium resources native to China. Genet. Resour. Crop Evol. 2013, 61, 395–412. [Google Scholar] [CrossRef]
- Suh, J.K.; Wu, X.W.; Lee, A.K.; Roh, M.S. Growth and flowering physiology, and developing new technologies to increase the flower numbers in the Genus Lilium. Hortic. Environ. Biotechnol. 2013, 54, 373–387. [Google Scholar] [CrossRef]
- Asano, Y. Study on crosses between distantly related species of Lilies. V. Characteristics of newly obtained hybrids through embryo culture. J. Jpn. Soc. Hortic. Sci. 1980, 49, 392–396. [Google Scholar] [CrossRef] [Green Version]
- Seo, C.B. Ilustrated Flora of Korea; Hyangmoonsa: Seoul, Republic of Korea, 1979; pp. 206–209. [Google Scholar]
- Jia, W.J.; Ma, L.L.; Ding, K.; Cui, G.F.; Wu, L.F.; Wang, X.N.; Wang, J.H. Characters of sap flow of Lilium and its relations with environment factors. Xibei Zhiwu Xuebao 2012, 32, 2498–2505. [Google Scholar]
- Pereira, A. Plant Abiotic Stress Challenges from the Changing Environment. Front. Plant Sci. 2016, 7, 1123. [Google Scholar] [CrossRef] [Green Version]
- Hong, Y.K.; Jang, Y.J.; Seong, M.H.; Moon, M.S.; Choi, D.C. Viability of barley, wheat and oat by low temperature treatment. Korean J. Med. Crop Sci. 2003, 48, 172–173. [Google Scholar]
- Seo, Y.H.; Park, Y.S.; Jo, B.W.; Kang, A.S.; Jung, B.C.; Jung, Y.S. Regional distribution of peach freezing damage and chilling days in 2010 in Gangwon province. Korean J. Agric. For. Meteorol. 2010, 12, 225–231. [Google Scholar] [CrossRef] [Green Version]
- Lim, B.S.; Jung, D.S.; Yun, H.K.; Hwang, Y.S.; Chun, J.P. Symptoms of freezing injury and mechanical injury-induced fruit rot in ‘Niitaka’ pear fruit (Pyrus pyrifolia Nakai) during low temperature storage. Korean J. Hortic. Sci. Technol. 2005, 23, 282–286. [Google Scholar]
- Kwon, E.Y.; Jung, J.E.; Chung, U.R.; Lee, S.J.; Song, G.C.; Choi, D.G.; Yun, J.I. A thermal time-driven dormancy index as a complementary criterion for grape vine freeze risk evaluation. Korean J. Agric. For. Meteorol. 2006, 8, 1–9. [Google Scholar]
- Xu, J.; Chai, N.; Zhang, T.; Zhu, T.; Cheng, Y.; Sui, S.; Li, M.; Liu, D. Prediction of temperature tolerance in Lilium based on distribution and climate data. iScience 2021, 24, 102794. [Google Scholar] [CrossRef] [PubMed]
- Lindén, L.; Palonen, P.; Lindén, M. Relating Freeze-induced Electrolyte Leakage Measurements to Lethal Temperature in Red Raspberry. J. Am. Soc. Hortic. Sci. 2000, 125, 429–435. [Google Scholar] [CrossRef]
- Walker, D.J.; Romero, P.; Hoyos, A.; Correal, E. Seasonal change in cold tolerance, water relations and accumulation of cations and compatible so-lutes in Atriplex halimus L. Environ. Exp. Bot. 2008, 64, 217–224. [Google Scholar] [CrossRef]
- Von Fircks, H.A.; Verwijst, T. Plant viability as a function of temperature stress. Plant Physiol. 1993, 103, 125–130. [Google Scholar] [CrossRef]
- Burr, K.E.; Tinus, R.W.; Wallner, S.J.; King, R.M. Comparison of three cold hardness tests for conifer seedlings. Tree Physiol. 1990, 6, 351–369. [Google Scholar] [CrossRef]
- Janacek, J.; Prasil, I. Quantification of plant frost injury by nonlinear fitting of an S-shaped function. CryoLetters 1991, 12, 47–52. [Google Scholar]
- Iles, J.K.; Agnew, N.H. Seasonal cold-acclimation patterns of Sedum spectabile × telephium L. ‘Autumn Joy’ and Sedum spectabile Boreau. ‘Brilliant’. HortScience 1995, 30, 1221–1224. [Google Scholar] [CrossRef] [Green Version]
- Ingram, D.L.; Buchanan, D.W. Measurement of direct heat injury of roots of three woody plants. HortScience 1981, 16, 769–771. [Google Scholar] [CrossRef]
- Miller, G.L. Use of Dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Foolad, M.; Lin, G.Y. Relationship between cold tolerance during seed germination and vegetative growth in tomato: Analysis of response and correlated response to selection. J. Am. Soc. Hortic. Sci. 2001, 126, 216–220. [Google Scholar] [CrossRef] [Green Version]
- Sakai, A.; Weiser, C.J. Freezing resistance of trees in North America with reference to tree regions. Ecology 1973, 54, 118–126. [Google Scholar] [CrossRef]
- Sanghera, G.S.; Wani, S.H.; Hussain, W.; Singh, N.B. Engineering cold stress tolerance in crop plants. Curr. Genom. 2011, 12, 30–43. [Google Scholar] [CrossRef] [Green Version]
- Hyun, U.J.; Yeo, S.M.; Lee, S.B.; Lee, J.H.; Jeong, J.M.; Seong, Y.K.; Seo, D.H.; Won, Y.J.; Ahn, E.K.; Lee, J.H.; et al. Optimization of temperature regime to screen cold tolerant rice seedlings. Plant Breed. Biotechnol. 2016, 4, 176–187. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.H.; Beak, J.K.; Kim, K.M.; Kim, K.H.; Kang, C.S.; Lee, G.E.; Choi, J.Y.; Son, J.Y.; Ko, J.M.; Choi, C.H. Assessment of cold tolerance traits of Wheat cultivars using RGB images. Korean J. Breed. Sci. 2022, 54, 171–176. [Google Scholar] [CrossRef]
- Kim, G.S.; Sa, J.G.; Huh, B.L. Response to low temperature of rice culti-vars for mid-northern area at rooting stage after transplanting. Korean J. Crop Sci. 1989, 34, 170–176. [Google Scholar]
- Kreyling, J.; Schmid, S.; Aas, G. Cold tolerance of tree species is related to the climate of their native ranges. J. Biogeogr. 2015, 42, 156–166. [Google Scholar] [CrossRef]
- Zhu, G.; Liu, Z.; Zhu, P. A study on determination of lethal temperature with logistic function. J. Nanjing Agric. Univ. 1986, 9, 11–16. [Google Scholar]
- Repo, T.; Lehto, T.; Finér, L. Delayed soil thawing affects root and shoot functioning and growth in Scots pine. Tree Physiol. 2003, 10, 1583–1591. [Google Scholar] [CrossRef] [Green Version]
- Nesbitt, M.L.; Ebel, R.C.; Findley, D.; Wilkins, B.; Woods, F.; Himelrick, D. Assays to assess freeze injury of Satsuma mandarin. HortScience 2002, 37, 871–877. [Google Scholar] [CrossRef] [Green Version]
- Smith, B.A.; Reider, M.L.; Fletcher, J.S. Relationship between vital staining and subculture growth during the senescence of plant tissue cultures. Plant Physiol. 1982, 70, 1228–1230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oprisko, M.J.; Green, R.L.; Beard, J.B.; Gates, C.E. Vital staining of root hairs in 12 warm-season perennial grasses. Crop Sci. 1990, 30, 947–950. [Google Scholar] [CrossRef]
- Yadav, S.K. Cold stress tolerance mechanisms in plants. Agron. Sustain. Dev. 2010, 30, 515–527. [Google Scholar] [CrossRef] [Green Version]
- Lyubushkina, I.V.; Grabelnych, O.I.; Pobezhimova, T.P.; Stepanov, A.V.; Fedyaeva, A.V.; Fedoseeva, I.V.; Voilikov, V.K. Winter wheat cells subjected to freezing temperature undergo death process with features of programmed cell death. Protoplasma 2014, 251, 615–623. [Google Scholar] [CrossRef]
- Koukalova, B.; Kovarik, A.; Fajkus, J.; Siroky, J. Chromatin fragmentation associated with apoptotic changes in tobacco cells exposed to cold stress. FEBS Lett. 1997, 414, 289–292. [Google Scholar]
- Jing, H.H.; Savina, M.; Jing, D.; Devendran, A.; Ramakanth, K.K.; Xin, T.; Wei, S.S.; Mironova, V.V.; Jian, X. A sacrifice-for-survival mechanism protects root stem cell niche from chilling stress. Cell 2017, 170, 102. [Google Scholar]
- Truernit, E.; Haseloff, J. A simple way to identify non-viable cells within living plant tissue using confocal microscopy. Plant Methods 2008, 4, 15. [Google Scholar] [CrossRef] [Green Version]
- Yin, H.; Chen, Q.; Yi, M. Effects of short-term heat stress on oxidative damage and responses of antioxidant system in Lilium longiflorum. Plant Growth Regul. 2007, 54, 45–54. [Google Scholar] [CrossRef]
- Xin, H.; Zhang, H.; Chen, L.; Li, X.; Lian, Q.; Yuan, X.; Hu, X.; Cao, L.; He, X.; Yi, M. Cloning and characterization of HsfA2 from lily (Lilium longiflorum). Plant Cell Rep. 2010, 29, 875–885. [Google Scholar] [CrossRef]
- Du, F.; Anderson, N. Bulb tissue cold-tolerance sensitivity differences in non-hardy and winter hardy Lilium. HortScience 2009, 44, 1013. [Google Scholar]
- Tian, X.; Xie, J.; Yu, J. Physiological and transcriptomic responses of Lanzhou Lily (Lilium davidii, var. unicolor) to cold stress. PLoS ONE 2020, 15, e0227921. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.A.; Kenna, M.P.; Taliaferro, C.M. Cold hardiness of ‘Midiron’ and ‘Tifgreen’ bermudagrass. HortScience 1988, 23, 748–750. [Google Scholar] [CrossRef]
- Cardona, C.A.; Duncan, R.R.; Lindstrom, O. Low temperature tolerance assessment in Paspalum. Crop Sci. 1997, 37, 1283–1291. [Google Scholar] [CrossRef]
- Kim, I.H.; Huh, K.Y.; Huh, M.R. Cold tolerance assessment of Sedum species for shallow-extensive green roof system. Korean J. Hortic. Sci. Technol. 2010, 28, 22–30. [Google Scholar]
- Ruelland, E.; Vaultier, M.N.; Zachowski, A.; Hurry, V. Cold signalling and cold acclimation in plants. Adv. Bot. Res. 2009, 49, 35–150. [Google Scholar]
- Tarkowski, L.P.; Ende, W.V. Cold tolerance triggered by soluble sugars: A multifaceted countermeasure. Front. Plant Sci. 2015, 6, 203. [Google Scholar] [CrossRef] [Green Version]
- Kooiker, M.; Drenth, J.; Glassop, D.; McIntyre, C.L.; Xue, G.P. TaMYB13-1, a R2R3 MYB transcription factor, regulates the fructan synthetic pathway and contributes to enhanced fructan accumulation in bread wheat. J. Exp. Bot. 2013, 64, 3681–3696. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.B. Korea Illustrated Plant Book; Hyangmunsa: Seoul, Republic of Korea, 1982; pp. 1–791. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Scientific Name | Electrolyte Leakage (Mean ± SE, %) | ||||
|---|---|---|---|---|---|
| 4 °C | 0 °C | −4 °C | −8 °C | −12 °C | |
| H. fulva | 3.9 ± 0.74 b* | 4.3 ± 0.59 b | 5.7 ± 0.83 a | 70.7 ± 9.33 a | 76.1 ± 13.46 a |
| H. longipes | 7 ± 1.16 a | 8.5 ± 0.89 a | 9.4 ± 5.89 a | 42.2 ± 24.16 b | 89.9 ± 3.75 a |
| H. plantaginea | 2.4 ± 0.37 b | 3.6 ± 0.39 b | 3.9 ± 1.04 a | 45.4 ± 8.25 ab | 76.1 ± 10.9 a |
| S. scilloides | 2.7 ± 0.55 b | 3.7 ± 0.85 b | 7.4 ± 1.53 a | 10 ± 2.4 c | 69.5 ± 14.35 a |
| Scientific Name | Estimated Lethal Temperature (°C) | Confidence Limits (95%) | |
|---|---|---|---|
| Lowest Temp. (°C) | Highest Temp. (°C) | ||
| Hemerocallis fulva | −7.15 | −8.44 | −5.86 |
| Hosta longipes | −8.87 | −9.71 | −8.03 |
| Hosta plantaginea | −11.14 | −11.71 | −10.58 |
| Scilla scilloides | −8.77 | −9.68 | −7.86 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, W.-H.; Yong, S.-H.; Park, D.-J.; Ahn, S.-J.; Kim, D.-H.; Park, K.-B.; Jin, E.-J.; Choi, M.-S. Efficient Cold Tolerance Evaluation of Four Species of Liliaceae Plants through Cell Death Measurement and Lethal Temperature Prediction. Horticulturae 2023, 9, 751. https://doi.org/10.3390/horticulturae9070751
Yang W-H, Yong S-H, Park D-J, Ahn S-J, Kim D-H, Park K-B, Jin E-J, Choi M-S. Efficient Cold Tolerance Evaluation of Four Species of Liliaceae Plants through Cell Death Measurement and Lethal Temperature Prediction. Horticulturae. 2023; 9(7):751. https://doi.org/10.3390/horticulturae9070751
Chicago/Turabian StyleYang, Woo-Hyeong, Seong-Hyeon Yong, Dong-Jin Park, Sung-Jin Ahn, Do-Hyun Kim, Kwan-Been Park, Eon-Ju Jin, and Myung-Suk Choi. 2023. "Efficient Cold Tolerance Evaluation of Four Species of Liliaceae Plants through Cell Death Measurement and Lethal Temperature Prediction" Horticulturae 9, no. 7: 751. https://doi.org/10.3390/horticulturae9070751