
Physiological and Molecular Screening of High Temperature Tolerance in Okra [Abelmoschus esculentus (L.) Moench]

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. DNA Isolation, Quantification and Qualification
2.3. SSR Primers and Polymerase-Chain Reaction (PCR)
2.4. Photosynthetic Parameters
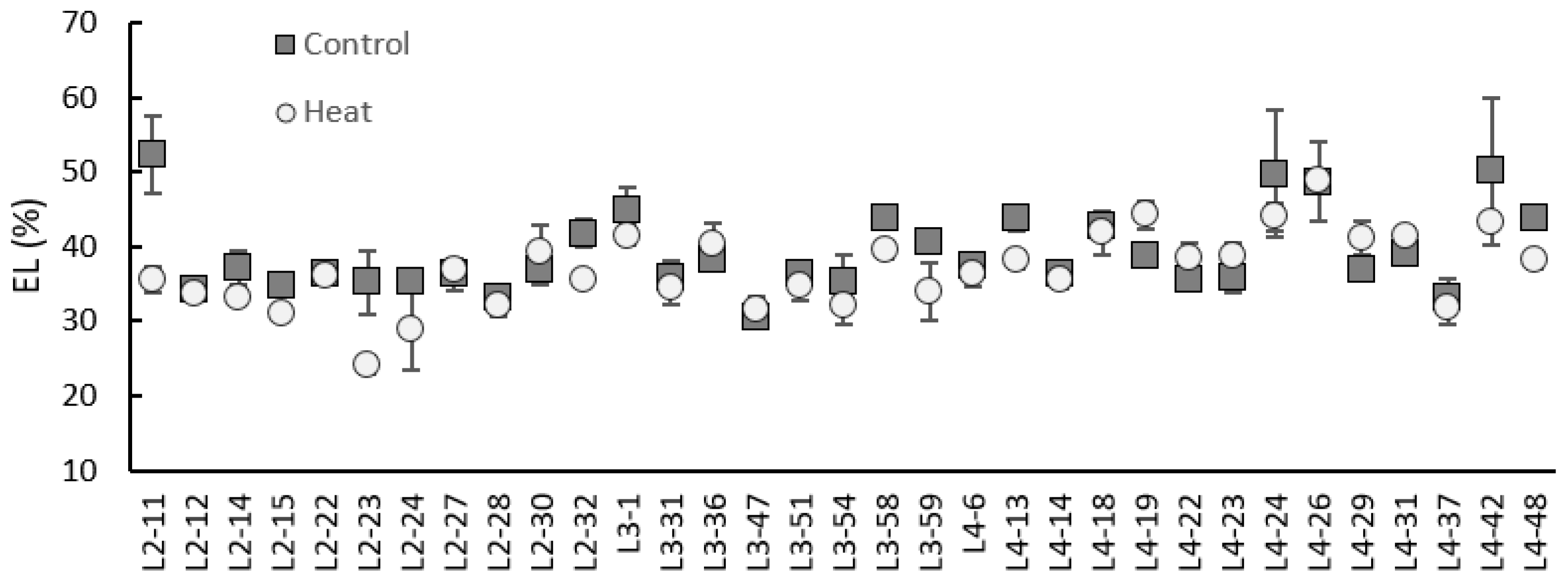
2.5. Electrolyte Leakage
2.6. Initial Screening of 104 Genotypes: Four-Hour Heat Shock (45 °C)
2.7. Advanced Screening of 33 Genotypes: Six-Hour Heat Shock (45 °C)
2.8. Statistical Data Analysis
3. Results
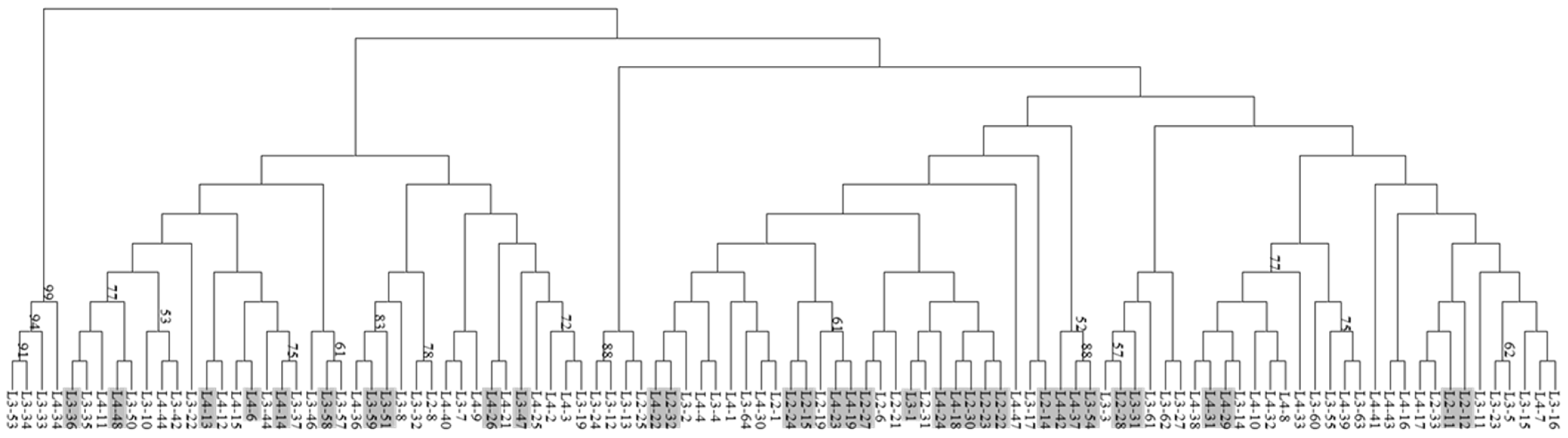
3.1. DNA Extraction and Dendrogram Generation Using SSR Primers
3.2. Initial Physiological Screening of 104 Genotypes after Four-Hour Heat Shock (45 °C)
- 45.5% with low Fv′/Fm′, low gs;
- 12.1% with low Fv′/Fm′, high gs;
- 6.1% with high Fv′/Fm′, high gs;
- 21.2% with similar Fv′/Fm′, low gs;
- 15.1% with similar Fv′/Fm′, high gs.
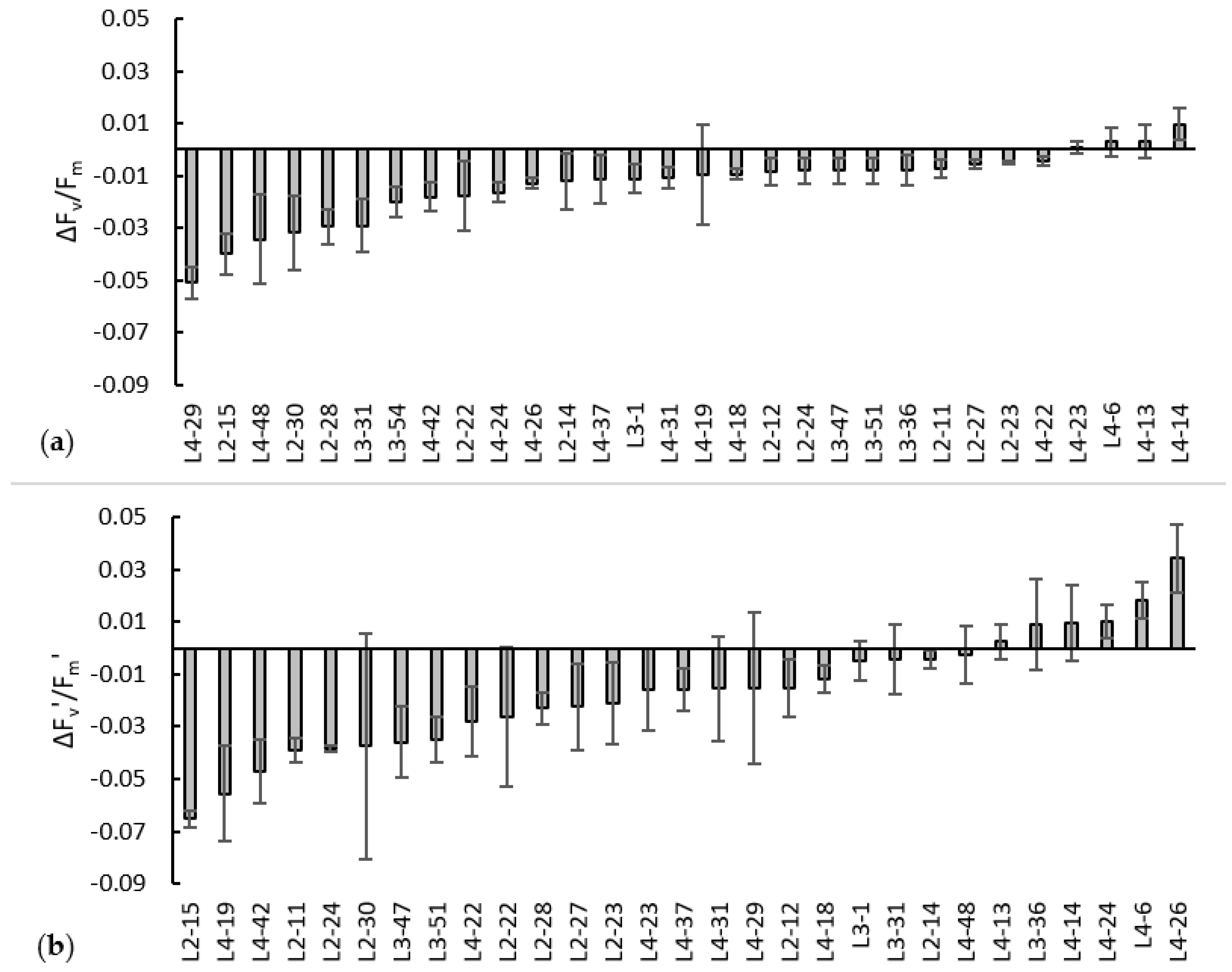
3.3. In-Depth Physiological Screening of 33 Genotypes after Six-Hour Heat Shock (45 °C)
4. Discussion
4.1. Modified DNA Extraction Protocol, SSR Markers and Polymorphism in Okra
4.2. Physiological Response of Okra to Heat Shock
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | Name (Marker) | ID | Repeated Motif | Forward Primer Sequences (5′-3′) | Reverse Primer Sequences (5′-3′) |
|---|---|---|---|---|---|
| 1 | AVRDC-Okra1 | 5200 | (AAG)13 | F:ATGGAGTGATTTTTGTGGAG | R:GACCCGAACTCACGTTACTA |
| 2 | AVRDC-Okra8 | 128713 | (AAG)8 | F:TGCTGTGGAAGGTTTTTACT | R:ATGACGAAAGTGGTGAAAAG |
| 3 | AVRDC-Okra9 | 89235 | (AAT)12 | F:ACCTTGAACACCAGGTACAG | R:TTGCTCTTATGAAGCAGTGA |
| 4 | AVRDC-Okra17 | 8461 | (AGA)7 | F:ACGAGAGTGAAGTGGAACTG | R:CTCCTCTTTCCTTTTTCCAT |
| 5 | AVRDC-Okra21 | 43380 | (AGA)9 | F:TCATGTCTTTCCACTCAACA | R:CCAAACAAAATATGCCTCTC |
| 6 | AVRDC-Okra28 | 151529 | (ATT)8 | F:CCTCTTCATCCATCTTTTCA | R:GGAAGATGCTGTGAAGGTAG |
| 7 | AVRDC-Okra39 | 51708 | (AG)16 | F:TGAGGTGATGATGTGAGAGA | R:TTGTAGATGAGGTTTGAACG |
| 8 | AVRDC-Okra52 | 126731 | (CAT)8-(TCA)9 | F:AACACATCCTCATCCTCATC | R:ACCGGAAGCTATTTACATGA |
| 9 | AVRDC-Okra54 | 87311 | (GAA)10 | F:CGAAAAGGAAACTCAACAAC | R:TGAACCTTATTTTCCTCGTG |
| 10 | AVRDC-Okra56 | 21030 | (GAA)44 | F:GGCAACTTCGTAATTTCCTA | R:TGAGTAAAAGTGGGGTCTGT |
| 11 | AVRDC-Okra57 | 151995 | (GAA)9-(GAG)7 | F:CGAGGAGACCATGGAAGAAG | R:ATGAGGAGGACGAGCAAGAA |
| 12 | AVRDC-Okra63 | 34632 | (TCT)12 | F:GTGTTTGAAAGGGACTGTGT | R:CTTCATCAAAACCATGCAG |
| 13 | AVRDC-Okra64 | 5886 | (TCT)22 | F:AAGGAGGAGAAAGAGAAGGA | R:ATTTACTTGAGCAGCAGCAG |
| 14 | AVRDC-Okra66 | 20291 | (TTC)12-(TTC)13 | F:CACCAGAATTTCCCTTTTG | R:ACTGTTGTTTGGCTTATGCT |
| 15 | AVRDC-Okra70 | 89044 | (TC)11 | F:GTAGCTGAACCCTTTGCTTA | R:CTATCATGGCGGATTCTTTA |
| 16 | AVRDC-Okra77 | 152270 | (GAAATA)4-(GAAACA)7 | F:CTGTTTGTTCGTCGTAATCA | R:AAAGTTTCTTCCTTTCCACC |
| 17 | AVRDC-Okra78 | 122488 | (TAT)11-(TATTGT)4-(TATCGT)4 | F:CTCCGACAATTCAAGAAAAG | R:CACCCAATCAAGCTATGTTA |
| 18 | AVRDC-Okra86 | 461 | (AGC)8 | F:ATGCAAACAAGCTAGTGGAT | R:ATTCTCTTCAGGGTTTCCTC |
| 19 | AVRDC-Okra89 | 129459 | (AGC)8 | F:TTTGAGTTCTTTCGTCCACT | R:GTATTTGGACATGGCGTTAT |
| 20 | 3 | (AAC)5 | F:TGGTGACGACATACAAGAAAAGA | R:CCCGGTGGTTTAGGAAGTTT | |
| 21 | 7 | (AAC)6 | F:ACCACTTCTCCATCCATCCA | R:AGCTTGCTGCATGAGTGCT | |
| 22 | 8 | (AAC)5-(AAC)6 | F:CAAAGGCACTTCATCAGCAA | R:GTGAGCGTCAATGTTGGATG | |
| 23 | 20 | (AAG)5 | F:TGAAGGTCAAATTGCCAAGA | R:TCCTTGTTTTTGAAGGTCACG | |
| 24 | 27 | (AAG)6 | F:CGATCGGAACGAGGACTTTA | R:CCCCGTTTTTCTTCTCTCCT | |
| 25 | 35 | (AAG)8 | F:GAAGAAGAAAAAGAGATAGATCTGTGG | R:GGCAGGAACAGATCCTTGAA | |
| 26 | 55 | (AAG)6 | F:CAGTTCGGGAAGAGGACAAA | R:ATCCCAAACCAGGTTCTTCA | |
| 27 | 62 | (AT)10 | F:TTCCGCCCATAGTCTTTGAC | R:TGAAAGGGCTTAGAGGGTTTT | |
| 28 | 74 | (AT)16 | F:GGTGGAAGGAACAACTCTGG | R:CCGGCATGATTAAGACACAC | |
| 29 | 82 | (TC)11 | F:CACTTTCCACACTCAAACCA | R:GAGAGGATTTCGGTGATGT | |
| 30 | 95 | (TCC)6 | F:AAAGGTGTTGGGTTTTGTGG | R:AGGAAGGAGAGGGACGAAAG | |
| 31 | 96 | (TCC)6 | F:CCAGTGGCAGCTACGGTACTA | R:GAGACGGAGGAGAAGTTGCTT | |
| 32 | 103 | (TG)5 | F:TGGGTTGTCCTTCTTTTTGG | R:GGGTGCAGAAGTTTGACCA | |
| 33 | 107 | (AC)5 | F:CAAACCATTTCCTCCATTGTG | R:TACGTAGCCCCTTGCTCATT | |
| 34 | 135 | (AG)10 | F:GCTGACTGGACGGATCTGAG | R:CCAAAGCATAAGCATTCATTCA | |
| 35 | 136 | (AT)5 | F:TTTGTGTCGAGAGATGCACA | R:CTTGAAACTTCAACGGCATT |
References
- Wahid, A.; Gelani, S.; Ashraf, M.; Foolad, M. Heat tolerance in plants: An overview. Environ. Exp. Bot. 2007, 61, 199–223. [Google Scholar] [CrossRef]
- Kakani, V.G.; Reddy, K.R.; Koti, S.; Wallace, T.P.; Prasad, P.V.; Reddy, V.R.; Zhao, D. Differences in in vitro pollen germination and pollen tube growth of cotton cultivars in response to high temperature. Ann. Bot. 2005, 96, 59–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Shaieny, A.; Bashandy, T. Effect of Planting Dates on Growth, Yield and Physiological Traits of Okra (Abelmoschus esculentus L. Moench.), and Field Evaluation for Heat Tolerance. J. Plant Prod. 2022, 13, 141–150. [Google Scholar] [CrossRef]
- Adeboye, O.; Oputa, C. Effects of galex on growth and fruit nutrient composition of okra (Abelnurschus esculentus). Ife J. Agric. 1996, 18, 1–9. [Google Scholar]
- Gunawardhana, M.; De Silva, C. Impact of temperature and water stress on growth yield and related biochemical parameters of okra. Trop. Agric. Res. 2012, 23, 77–83. [Google Scholar] [CrossRef] [Green Version]
- Arulrajah, T.; Ormrod, D. Responses of okra (Hibiscus esculentus L.) to photoperiod and temperature. Ann. Bot. 1973, 37, 331–340. [Google Scholar] [CrossRef]
- Jamala, G.; Boni, P.; Abraham, P.; Musa, A. Soil status and yield response of different varieties of okra (Abelmoschus esculentus (L.) Moench) grown at Mubi floodplain, North Eastern, Nigeria. J. Agric. Biotechnol. Sustain. Dev. 2011, 3, 120–125. [Google Scholar]
- Diizyaman, E. Okra: Botany and Horticulture. In Horticultural Reviews; Janick, J., Ed.; John Wiley & Sons: Hoboken, NJ, USA, 1997; Volume 21, pp. 41–72. [Google Scholar]
- Dhankhar, S.; Singh, S. Thermal requirements for flowering and fruit yield attainment in advance lines of okra. J. Agrometeorol. 2013, 15, 39–42. [Google Scholar] [CrossRef]
- Dhankhar, B.S.; Singh, R. Okra Handbook: Global Production, Processing, and Crop Improvement; HNB Publishing: New York, NY, USA, 2009. [Google Scholar]
- Hasanuzzaman, M.; Nahar, K.; Alam, M.M.; Roychowdhury, R.; Fujita, M. Physiological, biochemical, and molecular mechanisms of heat stress tolerance in plants. Int. J. Mol. Sci. 2013, 14, 9643–9684. [Google Scholar] [CrossRef]
- Van der Westhuizen, M.; Oosterhuis, D.; Berner, J.; Boogaers, N. Chlorophyll a fluorescence as an indicator of heat stress in cotton (Gossypium hirsutum L.). S. Afr. J. Plant Soil 2020, 37, 116–119. [Google Scholar] [CrossRef]
- Mercado Álvarez, K.; Bertero, H.D.; Paytas, M.J.; Ploschuk, E.L. Mesophyll conductance modulates photosynthetic rate in cotton crops exposed to heat stress under field conditions. J. Agron. Crop Sci. 2022, 208, 53–64. [Google Scholar] [CrossRef]
- Camejo, D.; Rodríguez, P.; Morales, M.A.; Dell’Amico, J.M.; Torrecillas, A.; Alarcón, J.J. High temperature effects on photosynthetic activity of two tomato cultivars with different heat susceptibility. J. Plant Physiol. 2005, 162, 281–289. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.-S.; Cheng, L. Photosystem 2 is more tolerant to high temperature in apple (Malus domestica Borkh.) leaves than in fruit peel. Photosynthetica 2009, 47, 112–120. [Google Scholar] [CrossRef]
- Teskey, R.; Wertin, T.; Bauweraerts, I.; Ameye, M.; McGuire, M.A.; Steppe, K. Responses of tree species to heat waves and extreme heat events. Plant Cell Environ. 2015, 38, 1699–1712. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Sharkey, T.D. Photosynthetic electron transport and proton flux under moderate heat stress. Photosynth. Res. 2009, 100, 29–43. [Google Scholar] [CrossRef] [PubMed]
- Martinazzo, E.G.; Ramm, A.; Bacarin, M.A. The chlorophyll a fluorescence as an indicator of the temperature stress in the leaves of Prunus persica. Braz. J. Plant Physiol. 2012, 24, 237–246. [Google Scholar] [CrossRef] [Green Version]
- Sinsawat, V.; Leipner, J.; Stamp, P.; Fracheboud, Y. Effect of heat stress on the photosynthetic apparatus in maize (Zea mays L.) grown at control or high temperature. Environ. Exp. Bot. 2004, 52, 123–129. [Google Scholar] [CrossRef]
- Ding, X.; Jiang, Y.; Hao, T.; Jin, H.; Zhang, H.; He, L.; Zhou, Q.; Huang, D.; Hui, D.; Yu, J. Effects of heat shock on photosynthetic properties, antioxidant enzyme activity, and downy mildew of cucumber (Cucumis sativus L.). PLoS ONE 2016, 11, e0152429. [Google Scholar] [CrossRef] [Green Version]
- Murchie, E.H.; Lawson, T. Chlorophyll fluorescence analysis: A guide to good practice and understanding some new applications. J. Exp. Bot. 2013, 64, 3983–3998. [Google Scholar] [CrossRef] [Green Version]
- Misra, A.N.; Misra, M.; Singh, R. Chlorophyll Fluorescence in Plant Biology; INTECH: London, UK, 2012. [Google Scholar]
- Strasser, R.J.; Tsimilli-Michael, M.; Srivastava, A. Analysis of the chlorophyll a fluorescence transient. Chlorophyll Fluoresc. 2004, 19, 321–362. [Google Scholar]
- Yamane, Y.; Shikanai, T.; Kashino, Y.; Koike, H.; Satoh, K. Reduction of QA in the dark: Another cause of fluorescence Fo increases by high temperatures in higher plants. Photosynth. Res. 2000, 63, 23–34. [Google Scholar] [CrossRef]
- Baker, N.R. Chlorophyll Fluorescence: A Probe of Photosynthesis In Vivo. Annu. Rev. Plant Biol. 2008, 59, 89–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, L.; Li, J.; Fan, Y.; Xu, S.; Zhang, Z. High temperature effects on photosynthesis, PSII functionality and antioxidant activity of two Festuca arundinacea cultivars with different heat susceptibility. Bot. Stud. 2006, 47, 61–69. [Google Scholar]
- Yamane, Y.; Kashino, Y.; Koike, H.; Satoh, K. Increases in the fluorescence Fo level and reversible inhibition of photosystem II reaction center by high-temperature treatments in higher plants. Photosynth. Res. 1997, 52, 57–64. [Google Scholar] [CrossRef]
- dos Santos Farias, D.B.; da Silva, P.S.O.; Tadeu Lucas, A.A.; de Freitas, M.I.; de Jesus Santos, T.; Nascimento Fontes, P.T.; de Oliveira Júnior, L.F.G. Physiological and productive parameters of the okra under irrigation levels. Sci. Hortic. 2019, 252, 1–6. [Google Scholar] [CrossRef]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence—A practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Roháček, K.; Soukupová, J.; Barták, M. Chlorophyll fluorescence: A wonderful tool to study plant physiology and plant stress. Plant Cell Compart.-Sel. Top. Res. Signpost Kerala India 2008, 41, 104. [Google Scholar]
- Reece, J.B.; Urry, L.A.; Cain, M.L.; Wasserman, S.A.; Minorsky, P.V.; Jackson, R.B. Campbell Biology; Pearson: Boston, MA, USA, 2011. [Google Scholar]
- Lu, Z.; Radin, J.W.; Turcotte, E.L.; Percy, R.; Zeiger, E. High yields in advanced lines of Pima cotton are associated with higher stomatal conductance, reduced leaf area and lower leaf temperature. Physiol. Plant. 1994, 92, 266–272. [Google Scholar] [CrossRef]
- Mc Daniel, R. The physiology of temperature effects on plants. In Breeding Plants for Less Favorable Environments; John Wiley and Sons: New York, NY, USA, 1982; pp. 13–45. [Google Scholar]
- Pilon, C.; Snider, J.L.; Oosterhuis, D.M.; Loka, D. The effects of genotype and irrigation regime on PSII heat tolerance in cotton. Sci. Int. 2016, 6, 1–11. [Google Scholar] [CrossRef]
- Lamont, W.J. Okra—A versatile vegetable crop. HortTechnology 1999, 9, 179–184. [Google Scholar] [CrossRef] [Green Version]
- Ariyo, O. Genetic diversity in West African okra (Abelmoschus caillei) (A. Chev.) Stevels—Multivariate analysis of morphological and agronomic characteristics. Genet. Resour. Crop Evol. 1993, 40, 25–32. [Google Scholar] [CrossRef]
- Aladele, S.E. Morphological distinctiveness and metroglyph analysis of fifty accessions of West African okra (Abelmoschus caillei) (A. Chev.) Stevels. J. Plant Breed. Crop Sci. 2009, 1, 273–280. [Google Scholar]
- Osawaru, M.; Ogwu, M.; Dania-Ogbe, F. Morphological assessment of the genetic variability among 53 Accessions of West African okra (Abelmoschus caillei (A. Chev.) Stevels) from South Western Nigeria. Niger. J. Basic Appl. Sci. 2013, 21, 227–238. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, F.J.; de Mendonca, T.F.N.; Siquieroli, A.C.S.; Maciel, G.; Clemente, A.A. Genetic and morphological descriptors to access Brazilian Okra genotypes diversity. Rev. Caatinga 2022, 35, 254–264. [Google Scholar] [CrossRef]
- Gulsen, O.; Karagul, S.; Abak, K. Diversity and relationships among Turkish okra germplasm by SRAP and phenotypic marker polymorphism. Biologia 2007, 62, 41–45. [Google Scholar] [CrossRef]
- Sawadogo, M.; Ouedraogo, J.T.; Balma, D.; Ouedraogo, M.; Gowda, B.S.; Botanga, C.; Timko, M.P. The use of cross species SSR primers to study genetic diversity of okra from Burkina Faso. Afr. J. Biotechnol. 2009, 8, 2476–2482. [Google Scholar]
- Schafleitner, R.; Kumar, S.; Lin, C.Y.; Hegde, S.G.; Ebert, A. The okra (Abelmoschus esculentus) transcriptome as a source for gene sequence information and molecular markers for diversity analysis. Gene 2013, 517, 27–36. [Google Scholar] [CrossRef]
- Kumar, S.; Parekh, M.J.; Fougat, R.S.; Patel, S.K.; Patel, C.B.; Kumar, M.; Patel, B.R. Assessment of genetic diversity among okra genotypes using SSR markers. J. Plant Biochem. Biotechnol. 2017, 26, 172–178. [Google Scholar] [CrossRef]
- Aladele, S.E.; Ariyo, O.; de Lapena, R. Genetic relationships among West African okra (Abelmoschus caillei) and Asian genotypes (Abelmoschus esculentus) using RAPD. Afr. J. Biotechnol. 2008, 7, 1426. [Google Scholar]
- Khan, A.A.; Azmat, M.A. Assessment of genetic diversity in okra (Abelmoschus esculentus L.) using RAPD markers. Pak. J. Agric. Sci. 2013, 50, 655–662. [Google Scholar]
- Martinello, G.; Leal, N.; Amaral, A., Jr.; Pereira, M.; Daher, R. Comparison of morphological characteristics and RAPD for estimating genetic diversity in Abelmoschus spp. Acta Hortic. (ISHS) 2000, 546, 101–104. [Google Scholar] [CrossRef]
- Akash, M.W.; Shiyab, S.M.; Saleh, M.I. Yield and AFLP analyses of inter-landrace variability in okra (Abelmoschus esculentus L.). Life Sci. J. 2013, 10, 771–779. [Google Scholar]
- Brake, V.; Irwin, J.; Park, R. Genetic variability in Australian isolates of Puccinia coronata f. sp. avenae assessed with molecular and pathogenicity markers. Australas. Plant Pathol. 2001, 30, 259–266. [Google Scholar] [CrossRef]
- Maréchal-Drouard, L.; Guillemaut, P. A powerful but simple technique to prepare polysaccharide-free DNA quickly and without phenol extraction. Plant Mol. Biol. Rep. 1995, 13, 26–30. [Google Scholar] [CrossRef]
- Sullivan, C.Y. Mechanisms of heat and drought resistance in grain sorghum and methods of measurement. In Sorghum in Seventies; Oxford & IBH Pub. Co.: New Delhi, India, 1972; p. 247. [Google Scholar]
- Lafuente, M.T.; Belver, A.; Guye, M.G.; Saltveit, M.E. Effect of temperature conditioning on chilling injury of cucumber cotyledons possible role of abscisic acid and heat shock proteins. Plant Physiol. 1991, 95, 443–449. [Google Scholar] [CrossRef]
- Nagy, S.; Poczai, P.; Cernák, I.; Gorji, A.M.; Hegedűs, G.; Taller, J. PICcalc: An online program to calculate polymorphic information content for molecular genetic studies. Biochem. Genet. 2012, 50, 670–672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Botstein, D.; White, R.L.; Skolnick, M.; Davis, R.W. Construction of a genetic linkage map in man using restriction fragment length polymorphisms. Am. J. Hum. Genet. 1980, 32, 314. [Google Scholar]
- Sneath, P.H.; Sokal, R.R. Numerical Taxonomy. The Principles and Practice of Numerical Classification; W H Freeman & Co.: New York, NY, USA, 1973. [Google Scholar]
- Rohlf, F. NTSYS-pc Numerical Taxonomy and Multivariate Analysis System, Version 2.02 e; EXETER Software Setauket: New York, NY, USA, 1998. [Google Scholar]
- Kankwatsa, P. Genetic Studies of Rust Resistance in the Triticum spp., and Puccinia graminis f. sp. Tritici Pathosystem; University of Sydeney, Faculty of Agriculture and Environment: Camperdown, Australia, 2016. [Google Scholar]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Nei, M.; Kumar, S. Prospects for inferring very large phylogenies by using the neighbor-joining method. Proc. Natl. Acad. Sci. USA 2004, 101, 11030–11035. [Google Scholar] [CrossRef] [Green Version]
- Fedoroff, N.; Battisti, D.; Beachy, R.; Cooper, P.; Fischhoff, D.; Hodges, C.; Knauf, V.; Lobell, D.; Mazur, B.; Molden, D. Radically rethinking agriculture for the 21st century. Science 2010, 327, 833–834. [Google Scholar] [CrossRef] [Green Version]
- Lobell, D.B.; Gourdji, S.M. The influence of climate change on global crop productivity. Plant Physiol. 2012, 160, 1686–1697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- IPCC. Climate Change 2007—The Physical Science Basis: Working Group I Contribution to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2007; Volume 4. [Google Scholar]
- Fang, G.; Hammar, S.; Grumet, R. A quick and inexpensive method for removing polysaccharides from plant genomic DNA. Biotechniques 1992, 13, 52–54, 56. [Google Scholar]
- Fougat, R.S.; Purohit, A.R.; Kumar, S.; Parekh, M.J.; Kumar, M. SSR based genetic diversity in Abelmoschus species. Indian J. Agric. Sci. 2015, 85, 1223–1228. [Google Scholar]
- Yildiz, M.; Koçak, M.; Baloch, F. Genetic bottlenecks in Turkish okra germplasm and utility of iPBS retrotransposon markers for genetic diversity assessment. Genet. Mol. Res. 2015, 14, 10588–10602. [Google Scholar] [CrossRef] [PubMed]
- Bibi, A.; Oosterhuis, D.; Gonias, E. Photosynthesis, quantum yield of photosystem II and membrane leakage as affected by high temperatures in cotton genotypes. J. Cotton Sci. 2008, 12, 150–159. [Google Scholar]
- Cottee, N.S.; Bange, M.P.; Wilson, I.W.; Tan, D.K. Developing controlled environment screening for high-temperature tolerance in cotton that accurately reflects performance in the field. Funct. Plant Biol. 2012, 39, 670–678. [Google Scholar] [CrossRef]
- Leigh, A.; Sevanto, S.; Close, J.D.; Nicotra, A.B. The influence of leaf size and shape on leaf thermal dynamics: Does theory hold up under natural conditions? Plant Cell Environ. 2017, 40, 237–248. [Google Scholar] [CrossRef]
- Zlatev, Z.S.; Yordanov, I.T. Effects of soil drought on photosynthesis and chlorophyll fluorescence in bean plants. Bulg. J. Plant Physiol. 2004, 30, 3–18. [Google Scholar]
- Jagadish, S.V.K.; Way, D.A.; Sharkey, T.D. Plant heat stress: Concepts directing future research. Plant Cell Environ. 2021, 44, 1992–2005. [Google Scholar] [CrossRef] [PubMed]
- Loka, D.A.; Oosterhuis, D.M.; Baxevanos, D.; Noulas, C.; Hu, W. Single and combined effects of heat and water stress and recovery on cotton (Gossypium hirsutum L.) leaf physiology and sucrose metabolism. Plant Physiol. Biochem. 2020, 148, 166–179. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R. Abiotic stress, the field environment and stress combination. Trends Plant Sci. 2006, 11, 15–19. [Google Scholar] [CrossRef]
- Rizhsky, L.; Liang, H.; Shuman, J.; Shulaev, V.; Davletova, S.; Mittler, R. When defense pathways collide. The response of Arabidopsis to a combination of drought and heat stress. Plant Physiol. 2004, 134, 1683–1696. [Google Scholar] [CrossRef] [Green Version]
- Drake, J.E.; Tjoelker, M.G.; Vårhammar, A.; Medlyn, B.E.; Reich, P.B.; Leigh, A.; Pfautsch, S.; Blackman, C.J.; López, R.; Aspinwall, M.J.; et al. Trees tolerate an extreme heatwave via sustained transpirational cooling and increased leaf thermal tolerance. Glob. Chang. Biol. 2018, 24, 2390–2402. [Google Scholar] [CrossRef]
- Osmond, C.; Grace, S. Perspectives on photoinhibition and photorespiration in the field: Quintessential inefficiencies of the light and dark reactions of photosynthesis? J. Exp. Bot. 1995, 46, 1351–1362. [Google Scholar] [CrossRef]
- Bukhov, N.G.; Wiese, C.; Neimanis, S.; Heber, U. Heat sensitivity of chloroplasts and leaves: Leakage of protons from thylakoids and reversible activation of cyclic electron transport. Photosynth. Res. 1999, 59, 81–93. [Google Scholar] [CrossRef]
- Feller, U.; Crafts-Brandner, S.J.; Salvucci, M.E. Moderately high temperatures inhibit ribulose-1, 5-bisphosphate carboxylase/oxygenase (Rubisco) activase-mediated activation of Rubisco. Plant Physiol. 1998, 116, 539–546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batcho, A.A.; Sarwar, M.B.; Rashid, B.; Hassan, S.; Husnain, T. Heat shock protein gene identified from Agave sisalana (AsHSP70) confers heat stress tolerance in transgenic cotton (Gossypium hirsutum). Theor. Exp. Plant Physiol. 2021, 33, 141–156. [Google Scholar] [CrossRef]
- Guo, Y.; Fan, R.; Sun, F.; Qu, Y.; Zheng, K.; Chen, Q.; Chen, Q. Identification of exogenous ABA and heat stress tolerance in various cotton genotypes. Plant Genet. Resour. 2020, 18, 404–416. [Google Scholar] [CrossRef]
- Kuo, C.; Chen, H.; Sun, H. Membrane thermostability and heat tolerance of vegetable leaves. In Adaptation of Food Crops to Temperature and Water Stress; Kuo, C.G., Ed.; AVRDC: Tainan City, Taiwan, 1992; pp. 160–168. [Google Scholar]




| Primer No. | Name | Repeat Motif | Primer Sequence (5′-3′) | Allele Size Range (bp) | No. of Alleles | PIC |
|---|---|---|---|---|---|---|
| 3 | AVRDC-Okra9 | (AAT)12 | F: ACCTTGAACACCAGGTACAG R: TTGCTCTTATGAAGCAGTGA | 150–250 | 4 | 0.56 |
| 6 | AVRDC-Okra28 | (ATT)8 | F: CCTCTTCATCCATCTTTTCA R: GGAAGATGCTGTGAAGGTAG | 200–300 | 3 | 0.53 |
| 9 | AVRDC-Okra54 | (GAA)10 | F: CGAAAAGGAAACTCAACAAC R: TGAACCTTATTTTCCTCGTG | 100–170 | 2 | 0.49 |
| 11 | AVRDC-Okra57 | (GAA)9- (GAG)7 | F: CGAGGAGACCATGGAAGAAG R: ATGAGGAGGACGAGCAAGAA | 170–310 | 4 | 0.43 |
| 12 | AVRDC-Okra63 | (TCT)12 | F: GTGTTTGAAAGGGACTGTGT R: CTTCATCAAAACCATGCAG | 200–300 | 2 | 0.27 |
| 13 | AVRDC-Okra64 | (TCT)22 | F: AAGGAGGAGAAAGAGAAGGA R: ATTTACTTGAGCAGCAGCAG | 100–300 | 7 | 0.71 |
| 18 | AVRDC-Okra86 | (AGC)8 | F: ATGCAAACAAGCTAGTGGAT R: ATTCTCTTCAGGGTTTCCTC | 250–400 | 4 | 0.65 |
| 19 | AVRDC-Okra89 | (AGC)8 | F: TTTGAGTTCTTTCGTCCACT R: GTATTTGGACATGGCGTTAT | 140–200 | 3 | 0.59 |
| Fv/Fm | Fv′/Fm′ | A | ΦPSII | gs | EL | ||
|---|---|---|---|---|---|---|---|
| Genotype | F | 2.14 | 6.23 | 6.41 | 3.09 | 9.94 | 4.34 |
| p-value | 0.003 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | |
| Treatment | F | 61.27 | 27.68 | 11.28 | 16.4 | 29.61 | 0.3 |
| p-value | <0.001 | <0.001 | 0.001 | <0.001 | <0.001 | 0.585 | |
| Genotype × Treatment | F | 2.27 | 1.8 | 4.88 | 2.99 | 5.29 | 1.62 |
| p-value | 0.001 | 0.017 | <0.001 | <0.001 | <0.001 | 0.031 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hayamanesh, S.; Trethowan, R.; Mahmood, T.; Ahmad, N.; Keitel, C. Physiological and Molecular Screening of High Temperature Tolerance in Okra [Abelmoschus esculentus (L.) Moench]. Horticulturae 2023, 9, 722. https://doi.org/10.3390/horticulturae9060722
Hayamanesh S, Trethowan R, Mahmood T, Ahmad N, Keitel C. Physiological and Molecular Screening of High Temperature Tolerance in Okra [Abelmoschus esculentus (L.) Moench]. Horticulturae. 2023; 9(6):722. https://doi.org/10.3390/horticulturae9060722
Chicago/Turabian StyleHayamanesh, Shahnoosh, Richard Trethowan, Tariq Mahmood, Nabil Ahmad, and Claudia Keitel. 2023. "Physiological and Molecular Screening of High Temperature Tolerance in Okra [Abelmoschus esculentus (L.) Moench]" Horticulturae 9, no. 6: 722. https://doi.org/10.3390/horticulturae9060722