
Integrated Control of Scales on Highbush Blueberry in Poland


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Taxonomic Identification of Scales Sampled from Blueberry Plantations
2.1.1. DNA Extraction, Amplification, and Sequencing
2.1.2. DNA Sequence Alignment and Phylogenetic Analysis
2.2. Assessment of Parthenolecanium spp. Parasites
2.3. Trials of Parthenolecanium sp. Control on Blueberry Plantations
2.4. Statistical Analysis
2.4.1. Phylogenetic Analysis
2.4.2. Field Data Analysis
3. Results
3.1. Taxonomic Identification of Highbush Blueberry Scale Specimens
3.2. Level of Parasitism of Parthenolecanium sp. under Field Conditions
3.3. Identification of Parthenolecanium spp. Parasites
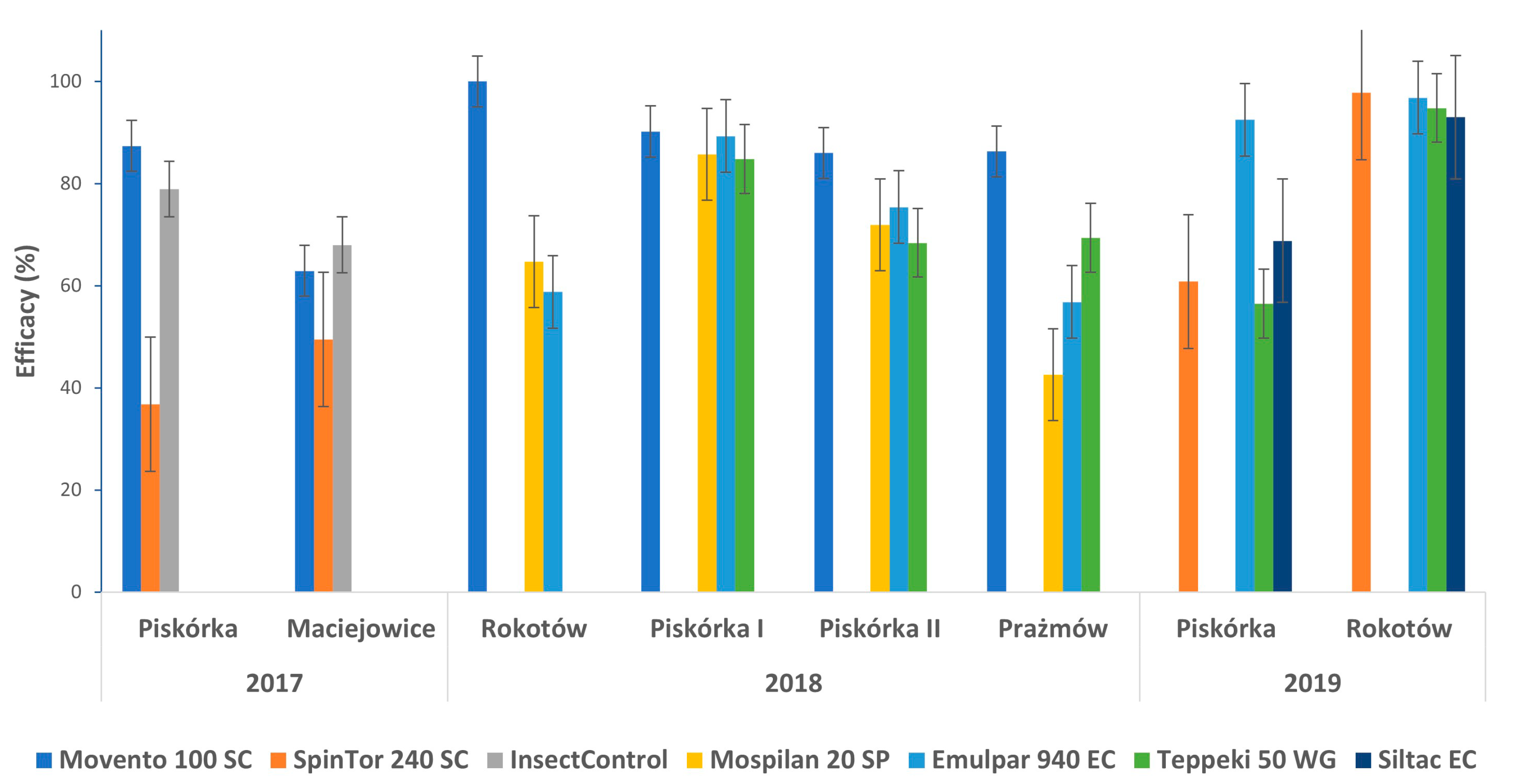
3.4. Control of Parthenolecanium sp. on Blueberry Plantations
4. Discussion
4.1. Phylogenetic Identification of the Specimens
4.2. Ecology of Parthenolecanium sp. Parasites
4.3. Is It Possible to Develop a Strategy for the Integrated Control of Parthenolecanium sp. in Highbush Blueberry Orchards with Low Environmental Impact?
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- International Blueberry Organization. Global State of the Blueberry Industry Report 2022. Available online: https://www.internationalblueberry.org/2022-report/ (accessed on 28 March 2023).
- Łabanowska, B.H.; Piotrowski, W. The spotted wing drosophila Drosophila suzukii (Matsumura, 1931)—Monitoring and first records in Poland. J. Hortic. Res. 2015, 23, 49–57. [Google Scholar] [CrossRef]
- Kalinowska, E.; Paduch-Cichal, E.; Chodorska, M.; Sala-Rejczak, K. First report of Blueberry red ringspot virus in highbush blueberry in Poland. J. Plant Pathol. 2011, 93, Supplement pp. S4.73. [Google Scholar]
- Tartanus, M.; Malusá, E.; Sas, D.; Łabanowska, B. Integrated control of Lecanium scale (Parthenolecanium spp.) on highbush blueberry in open field and protected crops. J. Plant Prot. Res. 2018, 58, 297–303. [Google Scholar] [CrossRef]
- Łagowska, B. Czerwce (Coccoidea), Zabielicowate (Ortheziidae), Czerwcowate (Margarodidae), Czerwce mączyste (Pseudococcidae), Pilśnikowate (Eriococcidae), Kermesowate (Kermesidae), Miłkowate (Cerococcidae), Misecznikowate (Coccidae), Gwiazdosze (Asterolecaniidae), Tarczniki (Diaspididae). In Fauna Polski—Charakterystyka i wykaz gatunków; Bogdanowicz, W., Chudzicka, E., Pilipiuk, I., Skibińska, E., Eds.; Muzeum and Instytut Zoologii PAN: Warszawa, Poland, 2004; pp. 266–269. [Google Scholar]
- Vranjic, J.A. Effects on host plant. In Soft Scale Insects: Their Biology, Natural Enemies and Control; Ben-Dov, Y., Hodgson, C.J., Eds.; Elsevier Science B.V.: Amsterdam, The Netherlands, 1997; Volume 7A, pp. 323–336. [Google Scholar]
- Robayo Camacho, E.; Chong, J.-H. General biology and current management approaches of soft scale pests (Hemiptera: Coccidae). J. Integr. Pest. Manag. 2015, 6, 1–22. [Google Scholar]
- Stark, J.D.; Vargas, R.; Banks, J.E. Incorporating ecologically relevant measures of pesticide effect for estimating the compatibility of pesticides and biocontrol agents. J. Econ. Entomol. 2007, 100, 1027–1032. [Google Scholar] [CrossRef]
- Trdan, S.; Laznik, Ž.; Bohinc, T. Thirty years of research and professional work in the field of biological control (predators, parasitoids, entomopathogenic and parasitic nematodes) in Slovenia: A review. Appl. Sci. 2020, 10, 7468. [Google Scholar] [CrossRef]
- Ponsonby, D.J.; Copland, M.J.W. Coccinellidae and other Coleoptera. In Soft Scale Insects: Their Biology, Natural Enemies and Control; Ben-Dov, Y., Hodgson, C.J., Eds.; Elsevier Science B.V.: Amsterdam, The Netherlands, 1997; Volume 7A, pp. 29–60. [Google Scholar]
- Kapranas, A.; Tena, A. Encyrtid Parasitoids of Soft Scale Insects: Biology, Behavior, and Their Use in Biological Control. Ann. Rev. Entomol. 2015, 60, 195–211. [Google Scholar] [CrossRef]
- Price, P.W.; Denno, R.F.; Eubanks, M.D.; Finke, D.L.; Kaplan, I. Insect Ecology: Behavior, Populations and Communities. Cambridge University Press: New York, NY, USA, 2011. [Google Scholar]
- Raupp, M.J.; Holmes, J.J.; Sadof, C.; Shrewsbury, P.; Davidson, J.A. Effects of cover sprays and residual pesticides on scale insects and natural enemies in urban forests. J. Arboric. 2001, 27, 203–214. [Google Scholar] [CrossRef]
- Gullan, P.J.; Cook, L.G. Phylogeny and higher classification of the scale insects (Hemiptera: Sternorrhyncha: Coccoidea). Zootaxa 2007, 425, 413–425. [Google Scholar] [CrossRef]
- Andersen, J.C.; Wu, J.; Gruwell, M.E.; Gwiazdowski, R.; Santana, S.H.; Feliciano, N.H.; Morse, G.E.; Normark, B.B. A phylogenetic analysis of armored scale insects (Hemiptera: Diaspididae), based upon nuclear, mitochondrial, and endosymbiont gene sequences. Mol. Phylogenet. Evol. 2010, 57, 992–1003. [Google Scholar] [CrossRef]
- Wang, X.-B.; Deng, J.; Zhang, J.-T.; Zhou, Q.-S.; Zhang, Y.-Z.; Wu, S.-A. DNA barcoding of common soft scales (Hemiptera: Coccoidea: Coccidae) in China. Bull. Entomol. Res. 2015, 105, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem Bull. 1987, 19, 11–15. [Google Scholar]
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Park, D.S.; Suh, S.J.; Oh, H.W.; Hebert, P.D.N. Recovery of the mitochondrial COI barcode region in diverse Hexapoda through tRNA-based primers. BMC Genom. 2010, 11, 423. [Google Scholar] [CrossRef] [PubMed]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA 11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Zhang, Z.; Schwartz, S.; Wagner, L.; Miller, W. A greedy algorithm for aligning DNA sequences. J. Comput. Biol. 2000, 7, 203–214. [Google Scholar] [CrossRef]
- Ronquist, F.M.; Teslenko, P.; van der Mark, D.; Ayres, A.; Darling, S.; Höhna, B.; Larget, L.; Liu, M.; Suchard, A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Abbott, W.S. A Method of Computing the Effectiveness of an Insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- Łagowska, B.; Golan, K.; Kot, I.; Kmieć, K.; Górska-Drabik, E.; Goliszek, K. Alien and invasive scale insect species in Poland and their threat to native plants. Bull. Insectology 2015, 68, 13–22. [Google Scholar]
- Stepaniuk, K.; Łagowska, B. Number and arrangement variation of submarginal tubercles in adult females Parthenolecanium corni group (Hemiptera, Coccidae) and its value as a taxonomic character. Pol. J. Entomol. 2006, 75, 293–301. [Google Scholar]
- Schultz, P.B. Natural enemies of oak lecanium (Homoptera: Coccidae) in eastern Virginia. Environ. Entomol. 1984, 13, 1515–1518. [Google Scholar] [CrossRef]
- Noyes, J.S. Universal Chalcidoidea Database; World Wide Web Electronic Publication, UK: 2019. Available online: http://www.nhm.ac.uk/chalcidoids (accessed on 10 March 2023).
- Meineke, E.K.; Dunn, R.R.; Frank, S.D. Early pest development and loss of biological control are associated with urban warming. Biol. Lett. 2014, 10, 20140586. [Google Scholar] [CrossRef] [PubMed]
- Robayo Camacho, E.; Chong, J.-H.; Braman, S.K.; Frank, S.D.; Schultz, P.B. Natural enemy communities and biological control of Parthenolecanium spp. (Hemiptera: Coccidae) in the Southeastern United States. J. Econ. Entomol. 2018, 111, 1558–1568. [Google Scholar] [CrossRef]
- Moglan, I. Complexes parasitaires de quelques espèces de coccides (Homoptera, Coccidae) en Roumanie. Entomol. Rom. 2007, 12, 267–275. [Google Scholar]
- Arnaoudov, V.; Olszak, R.; Kutinkova, H. Natural enemies of plum brown scale Parthenolecanium corni Bouché (Homoptera: Coccidae) in plum orchards in the region of Plovdiv. IOBC/Wprs Bull. 2006, 29, 105–109. [Google Scholar]
- Koteja, J. Notes on the Polish scale insect fauna (Homoptera, Coccoidea). IV. Polskie Pismo Entomol. 1972, 42, 565–571. [Google Scholar]
- Goliszek, K.; Łagowska, B.; Golan, K. Scale insects (Hemiptera, Sternorrhyncha, Coccoidea) on ornamental plants in the field in Poland. Acta Sci. Pol. Hortorum Cultus 2011, 10, 75–84. [Google Scholar]
- Graora, D.; Spasić, R.; Mihajlović, L. Bionomy of spruce bud scale, Physokermes piceae (Schrank) (Hemiptera: Coccidae) in the Belgrade area, Serbia. Arch. Biol. Sci. 2012, 64, 337–343. [Google Scholar] [CrossRef]
- Kosztarab, M.; Kozar, F. Introduction of Anthribus nebulosus (Coleoptera: Anthribidae) in Virginia for control of scale insects: A review. Virginia J. Sci. 1983, 34, 223–236. [Google Scholar]
- Kosztarab, M.; Kozár, F. Scale Insects of Central Europe; Series Entomologica; Akademiai Kiado: Budapest, Hungary, 1988; Volume 41, 456p. [Google Scholar]
- Dervisevic, M.; Graora, D. The life cycle and efficacy of Anthribus nebulosus Forster in reducing soft scale populations in Belgrade Fresenius Environ. Bull. 2019, 28, 1981–1985. [Google Scholar]
- Kidd, N.A.C.; Jervis, M.A. Population Dynamics. In Insects as Natural Enemies; Jervis, M.A., Ed.; Springer: Dordrecht, Germany, 2007. [Google Scholar] [CrossRef]
- Masner, L. Revisionary notes and keys to world genera of Scelionidae (Hymenoptera: Proctotrupoidea). Mem. Ent. Soc. Canada 1976, 108, 1–87. [Google Scholar] [CrossRef]
- Greathead, D.J. Opportunities for biological control of insect pests in tropical Africa. Rev. Zool. Afr. 1986, 100, 85–96. [Google Scholar]
- Falagiarda, M.; Carnio, V.; Chiesa, S.G.; Pignalosa, A.; Anfora, G.; Angeli, G.; Ioriatti, C.; Mazzoni, V.; Schmidt, S.; Zapponi, L. Factors influencing short-term parasitoid establishment and efficacy for the biological control of Halyomorpha halys with the samurai wasp Trissolcus japonicus. Pest Manag. Sci. 2023. [Google Scholar] [CrossRef]
- Seok-Min, L.; Bu-Keun, C.; Dong-Wan, K.; Kyung-Mi, P.; In-Young, H.; Jin-Hyeuk, K.; Heung-Su, L. Seasonal development and control of Parthenolecanium corni in blueberry Shrubs. Korean J. App. Entomol. 2021, 60, 403–415. [Google Scholar]
- Bacci, L.; Lupi, D.; Savoldelli, S.; Rossaro, B. A review of Spinosyns, a derivative of biological acting substances as a class of insecticides with a broad range of action against many insect pests. J. Entamol. Acarol. Res. 2016, 48, 40–52. [Google Scholar] [CrossRef]
- Duso, C.; Pozzebon, A.; Lorenzon, M.; Fornasiero, D.; Tirello, P.; Simoni, S.; Bagnoli, B. The Impact of Microbial and Botanical Insecticides on Grape Berry Moths and Their Effects on Secondary Pests and Beneficials. Agronomy 2022, 12, 217. [Google Scholar] [CrossRef]
- Williams, T.; Valle, J.; Viñuela, E. Is the naturally derived insecticide Spinosad® compatible with insect natural enemies? Biocontrol Sci. Technol. 2003, 13, 459–475. [Google Scholar] [CrossRef]
- Santana, V.; Santos, V.; Barbosa Pereira, B. Properties, toxicity and current applications of the biolarvicide spinosad. J. Toxicol. Environ. Health Part B 2020, 23, 13–26. [Google Scholar] [CrossRef]
- Biondi, A.; Mommaerts, V.; Smagghe, G.; Viñuela, E.; Zappalà, L.; Desneux, N. The non-target impact of spinosyns on beneficial arthropods. Pest Manag. Sci. 2012, 68, 1523–1536. [Google Scholar] [CrossRef] [PubMed]
- Suma, P.; Zappalà, L.; Mazzeo, G.; Siscaro, G. Lethal and sub-lethal effects of insecticides on natural enemies of citrus scale pests. BioControl 2009, 54, 651–661. [Google Scholar] [CrossRef]
- Somasundaran, P.; Mehta, S.C.; Purohit, P. Silicone emulsions. Adv. Colloid Interface Sci. 2006, 128-130, 103–109. [Google Scholar] [CrossRef]
- Hrastar, R.; Petrisic, M.G.; Ogrinc, N.; Kosir, I.J. Fatty acid and stable carbon isotope characterization of Camelina sativa oil: Implications for authentication. J. Agric. Food Chem. 2009, 57, 579–585. [Google Scholar] [CrossRef]
- Copping, L.G.; Duke, S.O. Natural products that had been used commercially as crop protection agents. Pest Manag. Sci, 2007, 63, 524–554. [Google Scholar] [CrossRef]
- Bakkali, F.; Averbeck, S.; Averbeck, D.; Idaomar, M. Biological effects of essential oils—A review. Food Chem. Toxicol. 2008, 46, 446–475. [Google Scholar] [CrossRef]
- Skalský, M.; Niedobová, J.; Popelka, J. The efficacy of European fruit lecanium, Parthenolecanium corni (Bouché, 1844) control using natural products. Hortic. Sci. 2019, 46, 195–200. [Google Scholar] [CrossRef]
- Gantner, M.; Jaśkiewicz, B.; Golan, K. Occurrence of Parthenolecanium corni (Bouché) on 18 cultivars of hazelnut. Folia Hortic. 2004, 16, 95–100. [Google Scholar]
- Vončina, A.; Novljan, M. Effect of selected insecticides on European fruit lecanium population (Parthenolecanium corni Buche) in a northern highbush blueberry (Vaccinium corymbosum L.) orchard. In Proceedings of the 14. Slovensko Posvetovanje o Varstvu Rastlin z Mednarodno Udelezbo, Maribor, Slovenija, 5–6 March 2019; pp. 247–251. [Google Scholar]
- Taylor-Wells, J.; Gross, A.D.; Jiang, S.; Demares, F.; Clements, J.S.; Carlier, P.R.; Bloomquist, J.R. Toxicity, mode of action, and synergist potential of flonicamid against mosquitoes Pestic. Biochem. Physiol. 2018, 151, 3–9. [Google Scholar]
- Morita, M.; Ueda, T.; Yoneda, T.; Koyanagi, T.; Haga, T. Flonicamid, a novel insecticide with a rapid inhibitory effect on aphid feeding. Pest Manag. Sci. 2007, 63, 969–973. [Google Scholar] [CrossRef] [PubMed]
- Karmakar, P.; Shera, P.S. Lethal and sublethal effects of insecticides used in cotton crop on the mealybug endoparasitoid Aenasius arizonensis. Int. J. Pest Manag. 2020, 66, 13–22. [Google Scholar] [CrossRef]
- Serrão, J.E.; Plata-Rueda, A.; Martínez, L.C.; Zanuncio, J.C. Side-effects of pesticides on non-target insects in agriculture: A mini-review. Sci. Nat. 2022, 109, 17. [Google Scholar] [CrossRef] [PubMed]
- Orr, D.B. Scelionid wasps as biological control agents: A review. Fla. Entomol. 1988, 71, 506–528. Available online: https://www.jstor.org/stable/3495011 (accessed on 10 March 2023). [CrossRef]
- Sial, A. Identification and Management of Scale Insects in Blueberries. 2022. Available online: https://site.extension.uga.edu/ipm/2022/08/05/identification-management-of-scale-insects-in-blueberries/ (accessed on 10 March 2023).
- Bahde, B.W.; Poojari, S.; Alabi, O.J.; Naidu, R.A.; Walsh, D.B. Pseudococcus maritimus (Hemiptera: Pseudococcidae) and Parthenolecanium corni (Hemiptera: Coccidae) are capable of transmitting Grapevine leafroll-associated virus between Vitis × labruscana and Vitis vinifera. Environ. Entomol. 2013, 42, 1292–1298. [Google Scholar] [CrossRef]
- Hommay, G.; Komar, V.; Lemaire, O.; Herrbach, E. Grapevine virus A transmission by larvae of Parthenolecanium corni. Eur. J. Plant Pathol. 2008, 121, 185–188. [Google Scholar] [CrossRef]
- Szyndel, M.S.; Paduch-Cichal, E. Detection of Blueberry red ringspot virus in different stages of Parthenolecanium corni in Poland. Ann. Wars. Univ. LifeSci.–SGGW Hortic. Landsc. Archit. 2020, 41, 77–81. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Scale Insect Species | Host | Locality | Isolate | GenBank No. Sequences |
|---|---|---|---|---|
| Parthenolecanium corni (Bouché) | No data | Chile | 24245 | KY085297 |
| No data | China | S3_499 | KP189846 | |
| Parthenolecanium fletcheri (Cockerell) | No data | No data | PARFLE | MZ 567176 |
| Platycladus orientalis | Chungcheongbuk-do, Korea | MK543920 | ||
| Parthenolecanium persicae (Fabricius) | No data | Australia | WIL | KY362203 |
| No data | China | S4_079c | KP189853 | |
| Parthenolecanium pomeranicum (Kawecki) | Taxus sp. | Poland | PAR_POM | MN603162 |
| Parthenolecanium pruinosum (Coquillett) | Vitis sp. L. | Australia | Mudgee_2a | KC784924 |
| Vitis sp. L. | Australia | Gumeracha_b | KC784923 | |
| Parthenolecanium sp. | Vaccinium corymbosum ‘Bluecrop’ | Piskórka, Poland | CBr3 | ON899817 |
| Vaccinium corymbosum ‘Bluecrop’ | Piskórka, Poland | CBr5 | ON899818 | |
| Vaccinium corymbosum ‘Bluecrop’ | Piskórka, Poland | CBr9 | ON899819 | |
| Vaccinium corymbosum ‘Bluecrop’ | Piskórka, Poland | CBr10 | ON899820 | |
| Vaccinium corymbosum ‘Bluecrop’ | Maciejowice, Poland | CBm11 | ON899821 | |
| Vaccinium corymbosum ‘Bluecrop’ | Maciejowice, Poland | CBm13 | ON899822 | |
| Vaccinium corymbosum ‘Bluecrop’ | Maciejowice, Poland | CBm14 | ON899823 |
| Gene | Primer | F or R | Primer Sequence 5′ to 3′ | References |
|---|---|---|---|---|
| COI | PcoF1 | F | CCTTCAACTAATCATAAAAATATYAG | [19] |
| HCO | R | TAAACTTCAGGGTGACCAAAAAATCA | [20] |
| Active Substance | Product | Mechanism of Action According to IRAC | Activity on the Pest |
|---|---|---|---|
| Camelina oil (Camelina sativa (L.) Crantz) | Emulpar 940 EC | N/A | Mechanical action and suffocation |
| Spinosad | SpinTor 240 EC | Nicotinic acetylcholine receptor (nAChR) allosteric modulators (Site I) | Contact and stomach poisoning, and ovicidal |
| Silicon polymers | Insect Control | N/A | Mechanical action and suffocation. |
| Silicon polymers | Siltac EC | N/A | Mechanical action, suffocation |
| Acetamipryd | Mospilan 20 SP Stonkat 20 SP | Nicotinic acetylcholine receptor (nAChR) competitive modulators | Contact and stomach poisoning |
| Flonicamid | Tepekki 50 WG | Chordotonal organ and Modulators—undefined target sites | Systemic |
| Spirotetramat | Movento 100 SC | Inhibitors of acetyl CoA carboxylase | Systemic |
| Parasite Species or Genera | Location of the Plantation | |||
|---|---|---|---|---|
| Jakubów | Piskórka | Prażmów | Rokotów | |
| Blastothrix spp. including B. brittanica (Girault, 1917) | X | X | X * | X |
| Coccophagus lycimnia (Walker, 1839) | X | X | X | X |
| Encyrtus infelix (Embleton, 1902) | X | X | X | X |
| Metaphycus spp. including M. insidiosus (Mercet, 1921) | X | X * | X | |
| Syrphophagus spp. including S. taeniatus (Förster, 1861) | X * | X | X | |
| Anthribidae spp. | X | |||
| Scelionidae spp. | X | |||
| Total | 4 | 5 | 6 | 5 |
| Treatment | Dose Applied | Living Larvae per Shoot (n) | |
|---|---|---|---|
| Piskórka | Maciejowice | ||
| Before treatment | - | 9.3 ± 26.9 | 10.0 ± 8.5 |
| Control | - | 7.9 ± 2.2 b | 34.3 ± 16.6 b |
| Spinosad | 0.4 L/ha | 5.0 ± 6.7 b | 17.3 ± 17.3 a |
| Silicon polymers | 0.2% | 1.7 ± 1.1 ab | 11.0 ± 15.4 a |
| Spirotetramat | 0.75 L/ha | 1.0 ± 2.1 a | 12.7 ± 11.4 a |
| Treatment | Dose Applied | Living Larvae per Shoot (n) | |||
|---|---|---|---|---|---|
| Rokotów | Piskórka I | Piskórka II (Tunel) | Prażmów | ||
| Before treatment | - | 2.5 ± 3.0 | 19.5 ± 18.2 | 20.7 ± 14.2 | 79.2 ± 33.2 |
| Control | - | 1.4 ± 1.3 b | 9.3 ± 5.1 b | 4.7 ± 3.1 b | 15.2 ±6.6 b |
| Camelina oil | 1.2% | 0.6 ± 0.8 ab | 1.0 ± 2.1 a | 1.2 ±3.5 a | 6.6 ± 13.5 ab |
| Acetamipryd | 0.2 kg/ha | 0.5 ± 1.5 ab | 1.3 ± 6.4 a | 1.3 ± 5.8 a | 8.7 ±20.9 ab |
| Flonicamid | 0.14 kg/ha | 1.6 ± 4.2 b | 1.4 ± 1.5 a | 1.5 ± 7.8 a | 4.7 ± 9.3 ab |
| Spirotetramat | 0.75 L/ha | 0.0 ± 0.0 a | 0.9 ± 3.7 a | 0.7 ± 2.9 a | 2.1 ± 6.8 a |
| Treatment | Dose Applied | Living Larvae per Shoot (n) | |
|---|---|---|---|
| Piskórka | Rokotów | ||
| Before treatment | - | 187.5 ± 36.7 | 336.0 ± 32.0 |
| Control | - | 15.5 ± 7.4 c | 248.2 ± 30.4 c |
| Spinosad | 0.4 L/ha | 6.1 ± 2.9 bc | 5.5 ± 9.1 a |
| Silicon polymers | 0.15% | 4.8 ± 7.8 bc | 17.3 ± 20.6 b |
| Acetamipryd | 0.2 L/ha | 1.2 ± 2.9 a | 8.0 ± 12.5 ab |
| Flonicamid | 0.14 kg/ha | 6.7 ± 5.4 bc | 12.9 ± 9.1 ab |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tartanus, M.; Sobieszek, B.; Furmańczyk-Gnyp, A.; Malusà, E. Integrated Control of Scales on Highbush Blueberry in Poland. Horticulturae 2023, 9, 604. https://doi.org/10.3390/horticulturae9050604
Tartanus M, Sobieszek B, Furmańczyk-Gnyp A, Malusà E. Integrated Control of Scales on Highbush Blueberry in Poland. Horticulturae. 2023; 9(5):604. https://doi.org/10.3390/horticulturae9050604
Chicago/Turabian StyleTartanus, Małgorzata, Barbara Sobieszek, Agnieszka Furmańczyk-Gnyp, and Eligio Malusà. 2023. "Integrated Control of Scales on Highbush Blueberry in Poland" Horticulturae 9, no. 5: 604. https://doi.org/10.3390/horticulturae9050604