
Using High-Density SNP Array to Investigate Genetic Relationships and Structure of Apple Germplasm in Bosnia and Herzegovina
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. SNP Data Genotyping
2.2. SSR Data
2.3. Statistical Analyses
3. Results and Discussion
3.1. SNP Polymorphism
3.2. SSR Polymorphism
3.3. Genetic Diversity
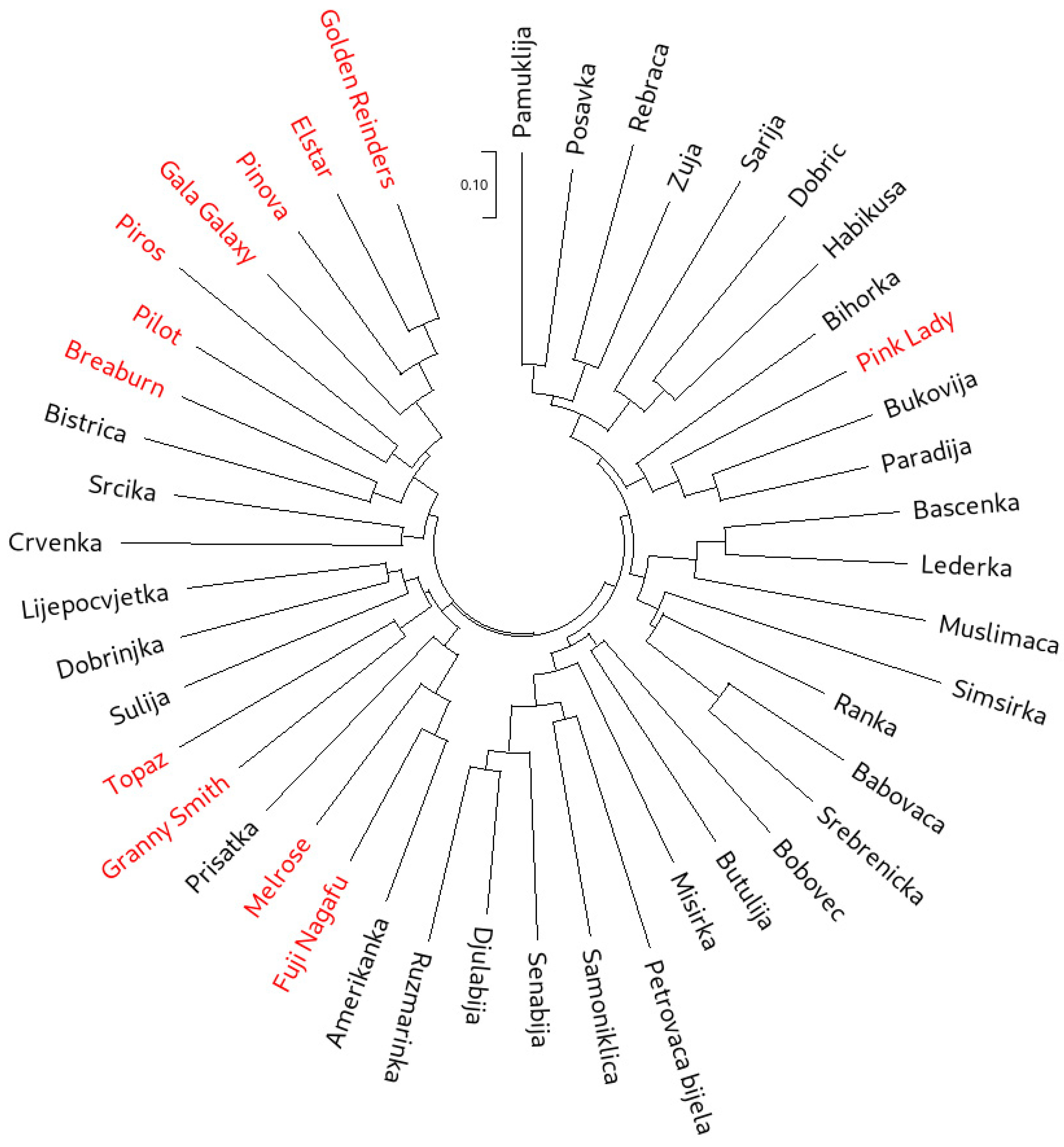
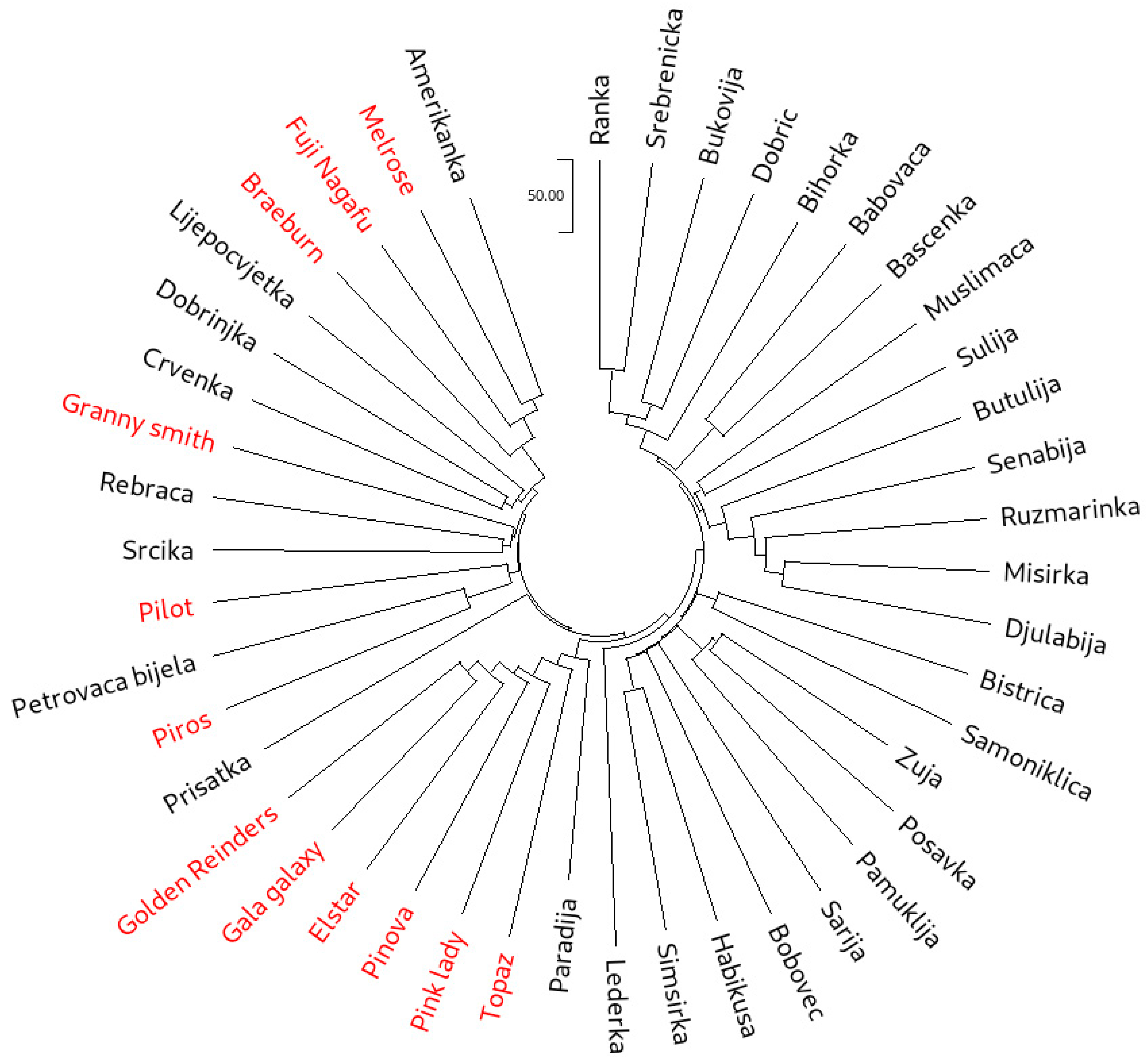
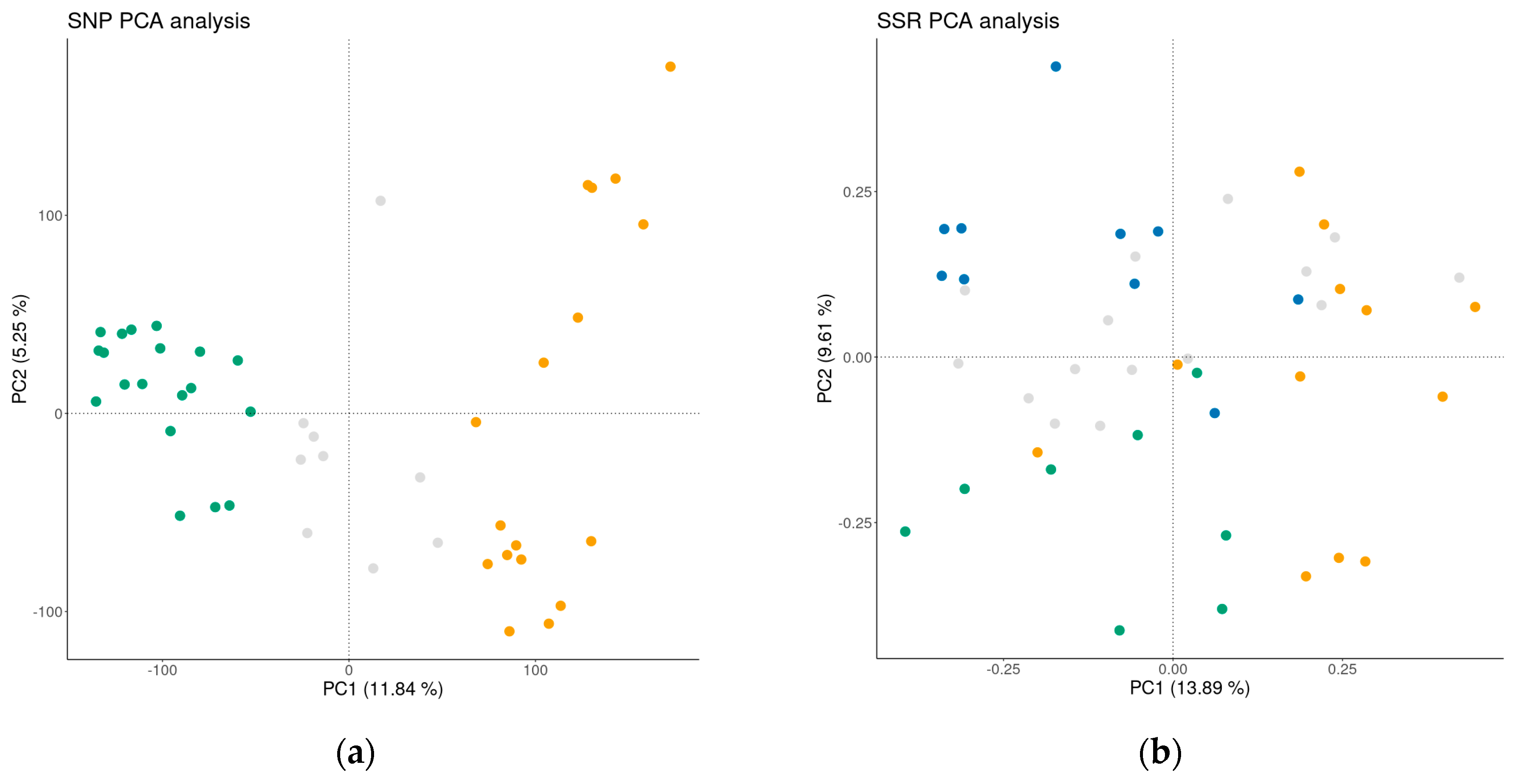
3.4. Genetic Relationships
3.4.1. Cluster Analyses
3.4.2. Identifying First-Degree Relationships
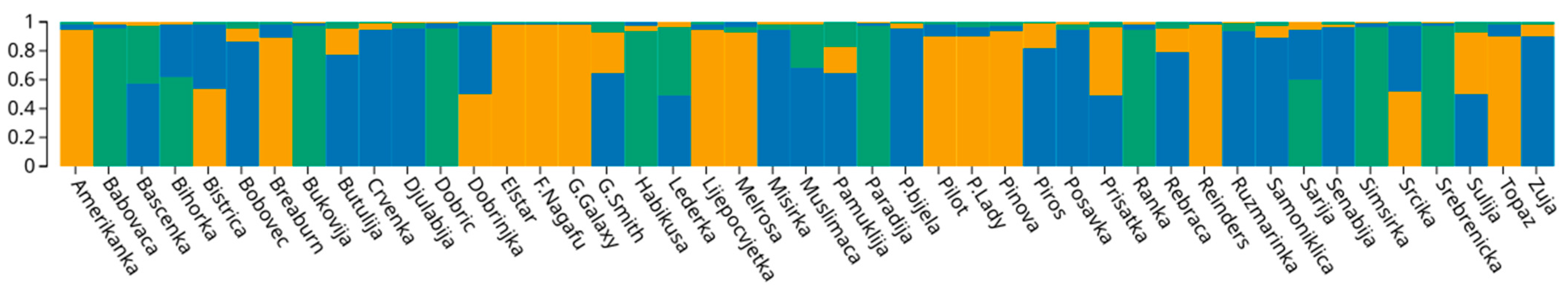
3.4.3. Genetic Structure
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nybom, H.; Lācis, G. Recent Large-Scale Genotyping and Phenotyping of Plant Genetic Resources of Vegetatively Propagated Crops. Plants 2021, 10, 415. [Google Scholar] [CrossRef] [PubMed]
- Rao, N.K. Plant Genetic Resources: Advancing Conservation and Use through Biotechnology. Afr. J. Biotechnol. 2004, 3, 136–145. [Google Scholar] [CrossRef]
- Nybom, H.; Weising, K. DNA-Based Identification of Clonally Propagated Cultivars. In Plant Breeding Reviews; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2010; Volume 34, pp. 221–295. [Google Scholar] [CrossRef]
- Liang, W.; Dondini, L.; De Franceschi, P.; Paris, R.; Sansavini, S.; Tartarini, S. Genetic diversity, population structure and construction of a core collection of apple cultivars from Italian germplasm. Plant Mol. Biol. Rep. 2015, 33, 458–473. [Google Scholar] [CrossRef]
- Frankel, O.H. Genetic Perspectives of Germplasm Conservation. In Genetic Manipulation: Impact on Man and Society; Arber, W.K., Llimensee, K., Peacock, W.J., Stralinger, P., Eds.; Cambridge University Press: Cambridge, UK, 1984; pp. 161–170. [Google Scholar]
- Nayak, S.N.; Song, J.; Villa, A.; Pathak, B.; Ayala-Silva, T.; Yang, X.; Todd, J.; Glynn, N.C.; Kuhn, D.N.; Glaz, B.; et al. Promoting Utilization of Saccharum spp. Genetic Resources through Genetic Diversity Analysis and Core Collection Construction. PLoS ONE 2014, 9, e110856. [Google Scholar] [CrossRef] [PubMed]
- Hokansson, S.C.; Szewc-McFadden, A.K.; Lamboy, W.F.; McFerson, J.R. Microsatellite (SSR) markers reveal genetic identities, genetic diversity and relationship in a Malus domestica Borkh. core subset collection. Theor. Appl. Genet. 1998, 97, 671–683. [Google Scholar] [CrossRef]
- Gharghani, A.; Zamani, Z.; Talaie, A.; Oraguzie, C.N.; Fatahi, R.; Hajnajari, H.; Wiedow, C.; Gardiner, S.E. Genetic identity and relationships of Iranian apple (Malus × domestica Borkh.) cultivars and landraces, wild Malus species and representative old apple cultivars based on simple sequence repeat (SSR) markers analysis Genet. Resour. Crop. Evol. 2009, 56, 829–842. [Google Scholar] [CrossRef]
- Van Treuren, R.; Kemp, H.; Ernsting, G.; Jongejans, B.; Houtman, H.; Visser, L. Microsatellite genotyping of apple (Malus × domestica Borkh.) genetic resources in the Netherlands: Application in collection management and variety identification. Genet. Resour. Crop Evol. 2010, 57, 853–865. [Google Scholar] [CrossRef]
- Ramos-Cabrer, A.M.; Diaz-Hernandez, M.B.; Pereira-Lorenzo, S. Use of microsatellites in the management of genetic resources of Spanish apple cultivars. J. Hort. Sci. Biotechnol. 2007, 82, 257–265. [Google Scholar] [CrossRef]
- Pereira-Lorenzo, S.; Ramos-Cabrer, A.M.; González-Díaz, A.J.; Díaz-Hernández, M.B. Genetic assessment of local apple cultivars from La Palma, Spain, using simple sequence repeats (SSRs). Sci. Hortic. 2008, 117, 160–166. [Google Scholar] [CrossRef]
- Urrestarazu, J.; Miranda, C.; Santesteban, L.; Royo, J. Genetic diversity and structure of local apple cultivars from Northeastern Spain assessed by microsatellite markers. Tree Genet. Genomes 2012, 8, 1163–1180. [Google Scholar] [CrossRef]
- Garkava-Gustavsson, L.; Kolodinska Brantestam, A.; Sehic, J.; Nybom, H. Molecular characterisation of indigenous Swedish apple cultivars based on SSR and S-allele analysis. Hereditas 2008, 145, 99–112. [Google Scholar] [CrossRef]
- Lassois, L.; Denancé, C.; Ravon, E.; Guyader, A.; Guisnel, R.; Hibrand-Saint-Oyant, L.; Poncet, C.; Lasserre-Zuber, P.; Feugey, L.; Durel, C.-E. Genetic Diversity, Population Structure, Parentage Analysis, and Construction of Core Collections in the French Apple Germplasm Based on SSR Markers. Plant Mol. Biol. Rep. 2016, 34, 827–844. [Google Scholar] [CrossRef]
- Larsen, A.S.; Asmussen, C.B.; Coart, E.; Olrik, D.C.; Kjær, E.D. Hybridization and genetic variation in Danish populations of European crab apple (Malus sylvestris). Tree Genet. Genomes 2006, 2, 86–97. [Google Scholar] [CrossRef]
- Cmejlova, J.; Rejlova, M.; Paprstein, F.; Cmejla, R. A new one-tube reaction kit for the SSR genotyping of apple (Malus × domestica Borkh.). Plant Sci. 2021, 303, 110768. [Google Scholar] [CrossRef] [PubMed]
- Bakir, M.; Dumanoglu, H.; Aygun, A.; Erdogan, V.; Efe Dost, S.; Gülsen, O.; Serdar, U.; Kalkisim, O.; Bastas, K. Genetic diversity and population structure of apple germplasm from Eastern Black Sea region of Turkey by SSRs. Sci. Hortic. 2022, 294, 110793. [Google Scholar] [CrossRef]
- Pikunova, A.V.; Sedov, E.N.; Tokmakov, S.V.; Suprun, I.I.; Gorbatchova, N.G.; Dolzhikova, M.A.; Yanchuk, T.V.; Serova, Z.M. Microsatellite Loci Polymorphism of Apple (Malus domestica Borkh.) Genotypes with Different Ploidy Level. Russ. J. Genet. 2018, 54, 442–450. [Google Scholar] [CrossRef]
- Gaši, F.; Kanlić, K.; Stroil, B.K.; Pojskić, N.; Asdal, Å.; Rasmussen, M.; Kaiser, C.; Meland, M. Redundancies and Genetic Structure among ex situ Apple Collections in Norway Examined with Microsatellite Markers. HortScience 2016, 51, 1458–1462. [Google Scholar] [CrossRef]
- Meland, M.; Aksic, M.F.; Frøynes, O.; Konjic, A.; Lasic, L.; Pojskic, N.; Gaši, F. Genetic Identity and Diversity of Apple Accessions within a Candidate Collection for the Norwegian National Clonal Germplasm Repository. Horticulturae 2022, 8, 630. [Google Scholar] [CrossRef]
- Gaši, F.; Simon, S.; Pojskic, N.; Kurtovic, M.; Pejic, I. Genetic assessment of apple germplasm in Bosnia and Herzegovina using microsatellite and morphologic markers. Sci. Hort. 2010, 126, 164–171. [Google Scholar] [CrossRef]
- Gaši, F.; Simon, S.; Pojskic, N.; Kurtovic, M.; Pejic, I.; Meland, M.; Kaiser, C. Evaluation of Apple (Malus × domestica) Genetic Resources in Bosnia and Herzegovina Using Microsatellite Markers. HortScience 2013, 48, 13–21. [Google Scholar] [CrossRef]
- Gaši, F.; Žulj Mihaljević, M.; Simon, S.; Grahić, J.; Pojskic, N.; Kurtović, M.; Nikolić, D.; Pejić, I. Genetic structure of apple accessions maintained ex situ in Bosnia and Herzegovina examined by microsatellite markers. Genetika 2013, 45, 467–478. [Google Scholar] [CrossRef]
- You, Q.; Yang, X.; Peng, Z.; Xu, L.; Wang, J. Development and Applications of a High Throughput Genotyping Tool for Polyploid Crops: Single Nucleotide Polymorphism (SNP) Array. Front. Plant Sci. 2018, 9, 104. [Google Scholar] [CrossRef] [PubMed]
- Chagné, D.; Crowhurst, R.N.; Troggio, M.; Davey, M.W.; Gilmore, B.; Lawley, C.; Vanderzande, S.; Hellens, R.P.; Kumar, S.; Cestaro, A.; et al. Genome-Wide SNP Detection, Validation, and Development of an 8K SNP Array for Apple. PLoS ONE 2012, 7, e31745. [Google Scholar] [CrossRef] [PubMed]
- Bianco, L.; Cestaro, A.; Sargent, D.J.; Banchi, E.; Derdak, S.; Di Guardo, M.; Salvi, S.; Jansen, J.; Viola, R.; Gut, I.; et al. Development and validation of a 20K single nucleotide polymorphism (SNP) whole genome genotyping array for apple (Malus × domestica Borkh). PLoS ONE 2014, 9, e110377. [Google Scholar] [CrossRef]
- Chagné, D.; Gašic, K.; Crowhurst, R.N.; Han, Y.; Bassett, H.C.; Bowatte, D.R.; Lawrence, T.J.; Rikkerink, E.H.; Gardiner, S.E.; Korban, S.S. Development of a set of SNP markers present in expressed genes of the apple. Genomics 2008, 92, 353–358. [Google Scholar] [CrossRef]
- Bianco, L.; Cestaro, A.; Linsmith, G.; Muranty, H.; Denancé, C.; Théron, A.; Poncet, C.; Micheletti, D.; Kerschbamer, E.; Di Pierro, E.A.; et al. Development and validation of the Axiom(®) Apple480K SNP genotyping array. Plant Mol. Biol. 2016, 86, 62–74. [Google Scholar] [CrossRef] [PubMed]
- Howard, N.P.; Van de Weg, E.; Bedford, D.S.; Peace, C.P.; Vanderzande, S.; Clark, M.D.; Teh, S.L.; Cai, L.; Luby, J.J. Elucidation of the ‘Honeycrisp’ pedigree through haplotype analysis with a multi-family integrated SNP linkage map and a large apple (Malus × domestica) pedigree-connected SNP data set. Hortic. Res. 2017, 4, 17003. [Google Scholar] [CrossRef]
- Chagné, D.; Kirk, C.; Whitworth, C.; Erasmuson, S.; Bicknell, R.; Sargent, D.J.; Kumar, S.; Troggio, M. Polyploid and aneuploid detection in apple using a single nucleotide polymorphism array. Tree Genet. Genomes 2015, 11, 94. [Google Scholar] [CrossRef]
- Minamikawa, M.F.; Kunihisa, M.; Noshita, K.; Moriya, S.; Abe, K.; Hayashi, T.; Katayose, Y.; Matsumoto, T.; Nishitani, C.; Terakami, S.; et al. Tracing founder haplotypes of Japanese apple varieties: Application in genomic prediction and genome-wide association study. Hortic. Res. 2021, 8, 49. [Google Scholar] [CrossRef]
- Volk, G.M.; Peace, C.P.; Henk, A.D.; Howard, N.P. DNA profiling with the 20K apple SNP array reveals Malus domestica hybridization and admixture in M. sieversii, M. orientalis, and M. sylvestris genebank accessions. Front. Plant Sci. 2022, 13, 1015658. [Google Scholar] [CrossRef]
- Muranty, H.; Denancé, C.; Feugey, L.; Crépin, J.-L.; Barbier, Y.; Tartarini, S.; Ordidge, M.; Troggio, M.; Lateur, M.; Nybom, H.; et al. Using whole-genome SNP data to reconstruct a large multi-generation pedigree in apple germplasm. BMC Plant Biol. 2020, 20, 2. [Google Scholar] [CrossRef]
- Urrestarazu, J.; Muranty, H.; Denancé, C.; Leforestier, D.; Ravon, E.; Guyader, A.; Guisnel, R.; Feugey, L.; Aubourg, S.; Celton, J.-M.; et al. Genome-Wide Association Mapping of Flowering and Ripening Periods in Apple. Front. Plant Sci. 2017, 8, 1923. [Google Scholar] [CrossRef] [PubMed]
- Gritsenko, D.; Pozharskiy, A.; Dolgikh, S.; Aubakirova, K.; Kenzhebekova, R.; Galiakparov, N.; Karimov, N.; Sadykov, S. Apple varieties from Kazakhstan and their relation to foreign cultivars assessed with RosBREED 10K SNP array. Eur. J. Hortic. Sci. 2022, 87, 1–8. [Google Scholar] [CrossRef]
- Larsen, B.; Gardner, K.; Pedersen, C.; Ørgaard, M.; Migicovsky, Z.; Myles, S.; Toldam-Andersen, T.B. Population structure, relatedness and ploidy levels in an apple gene bank revealed through genotyping-by-sequencing. PLoS ONE 2018, 13, e0201889. [Google Scholar] [CrossRef]
- Skytte af Sätra, J.; Troggio, M.; Odilbekov, F.; Sehic, J.; Mattisson, H.; Hjalmarsson, I.; Ingvarsson, P.K.; Garkava-Gustavsson, L. Genetic Status of the Swedish Central collection of heirloom apple cultivars. Sci. Hortic. 2020, 272, 109599. [Google Scholar] [CrossRef]
- Gianfranceschi, L.; Seglia, N.; Tarchini, R.; Komjanc, M.; Gessler, C. Simple sequence repeats for the genetic analyses of apple. Theor. Appl. Genet. 1998, 96, 1069–1079. [Google Scholar] [CrossRef]
- Liebhard, R.; Gianfranceschi, L.; Koller, B.; Ryder, C.D.; Tarchini, R.; Van De Weg, E.; Gessler, C. Development and characterization of 140 new microsatellite in apple (Malus × domestica Borkh.). Mol. Breed. 2002, 10, 217–241. [Google Scholar] [CrossRef]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.R.; Bender, D.; Maller, J.; Sklar, P.; de Bakker, P.I.W.; Daly, M.J.; et al. PLINK: A toolset for whole-genome association and population-based linkage analysis. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef]
- R Studio Team. RStudio: Integrated Development for R; RStudio, PBC: Boston, MA, USA, 2020; Available online: http://www.rstudio.com/ (accessed on 12 May 2022).
- Jombart, T. adegenet: A R package for the multivariate analysis of genetic markers. Bioinformatics 2008, 24, 1403–1405. [Google Scholar] [CrossRef]
- Kamvar, Z.N.; Tabima, J.F.; Grünwald, N.J. Poppr: An R package for genetic analysis of populations with clonal, partially clonal, and/or sexual reproduction. PeerJ 2014, 2, e281. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GENALEX 6: Genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research—An update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [PubMed]
- Statology. Two Proportion Z-Test Calculator. Available online: https://www.statology.org/two-proportion-z-test-calculator/ (accessed on 4 April 2023).
- Kalinowski, S.T.; Wagner, A.P.; Taper, M.L. ML-Relate: A computer program for maximum likelihood estimation of relatedness and relationship. Mol. Ecol. 2006, 6, 576–579. [Google Scholar] [CrossRef]
- Liu, K.; Muse, S.V. PowerMarker: An integrated analysis environment for genetic marker analysis. Bioinformatics 2005, 21, 2128–2129. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef]
- Mantel, N. The detection of disease clustering and a generalized regression approach. Cancer Res. 1967, 27, 209–220. [Google Scholar] [PubMed]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; ISBN 978-3-319-24277-4. Available online: https://ggplot2.tidyverse.org (accessed on 15 May 2022).
- Raj, A.; Stephens, M.; Pritchard, J.K. fastSTRUCTURE: Variational Inference of Population Structure in Large SNP Data Sets. Genetics 2014, 197, 573–589. [Google Scholar] [CrossRef]
- Li, Y.L.; Liu, J.X. StructureSelector: A web based software to select and visualize the optimal number of clusters using multiple methods. Mol. Ecol. Resour. 2018, 18, 176–177. [Google Scholar] [CrossRef]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef]
- Earl, D.A.; Von Holdt, B.M. Structure harvester: A Website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2011, 4, 359–361. [Google Scholar] [CrossRef]
- Marconi, G.; Ferradini, N.; Russi, L.; Concezzi, L.; Veronesi, F.; Albertini, E. Genetic Characterization of the Apple Germplasm Collection in Central Italy: The Value of Local Varieties. Front. Plant Sci. 2018, 9, 1460. [Google Scholar] [CrossRef] [PubMed]
- Pereira-Lorenzo, S.; Ramos-Cabrer, A.M.; Ferreira, V.; Díaz-Hernández, M.B.; Carnide, V.; Pinto-Carnide, O.; Rodrigues, R.; Velázquez Barrera, M.E.; Rios-Mesa, D.; Ascasíbar-Errasti, J.; et al. Genetic diversity and core collection of Malus × domestica in northwestern Spain, Portugal and the Canary Islands by SSRs. Sci. Hortic. 2018, 240, 49–56. [Google Scholar] [CrossRef]
- Ramasamy, R.; Ramasamy, S.; Bindroo, B.; Naik, V. STRUCTURE PLOT: A program for drawing elegant STRUCTURE bar plots in user friendly interface. SpringerPlus 2014, 3, 431. [Google Scholar] [CrossRef]
- Calus, M.P.L.; Vandenplas, J. SNPrune: An efficient algorithm to prune large SNP array and sequence datasets based on high linkage disequilibrium. Genet. Sel. Evol. 2018, 50, 34. [Google Scholar] [CrossRef] [PubMed]
- García, C.; Guichoux, E.; Hampe, A. A comparative analysis between SNPs and SSRs to investigate genetic variation in a juniper species (Juniperus phoenicea ssp. Turbinate). Tree Genet. Genomes 2018, 14, 87. [Google Scholar] [CrossRef]
- Emanuelli, F.; Lorenzi, S.; Grzeskowiak, L.; Catalano, V.; Stefanini, M.; Troggio, M.; Myles, S.; Martinez-Zapater, J.M.; Zyprian, E.; Moreira, F.M.; et al. Genetic diversity and population structure assessed by SSR and SNP markers in a large germplasm collection of grape. BMC Plant Biol. 2013, 13, 39. [Google Scholar] [CrossRef]
- Kanlić, K. Autohtone Sorte Voćaka Istočne Bosne; Štamparija Fojnica: Fojnica, Bosnia and Herzegovina, 2010. [Google Scholar]
- Amar, M.H.; Biswas, M.K.; Zhang, Z.; Guo, W.-W. Exploitation of SSR, SRAP and CAPS-SNP markers for genetic diversity of Citrus germplasm collection. Sci. Hortic. 2011, 128, 220–227. [Google Scholar] [CrossRef]
- Varshney, R.; Salem, K.; Baum, M.; Roder, M.; Graner, A.; Börner, A. SSR and SNP diversity in a barley germplasm collection. Plant Genet. Res. 2008, 6, 167–174. [Google Scholar] [CrossRef]
- Li, Y.-H.; Li, W.; Zhang, C.; Yang, L.; Chang, R.-Z.; Gaut, B.S.; Qiu, L.-J. Genetic diversity in domesticated soybean (Glycine max) and its wild progenitor (Glycine soja) for simple sequence repeat and single-nucleotide polymorphism loci. New Phytol. 2010, 188, 242–253. [Google Scholar] [CrossRef]
- Oras, A.; Akagić, A.; Spaho, N.; Gaši, F.; Žuljević, S.O.; Meland, M. Distribution and Stability of Polyphenols in Juices Made from Traditional Apple Cultivars Grown in Bosnia and Herzegovina. Molecules 2023, 28, 230. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accessions | GCs—SNP | GCs—SSR | Ex Situ Collection | |
|---|---|---|---|---|
| Traditional accessions | ||||
| 1 | ‘Amerikanka’ | 2 | 1 | GO |
| 2 | ‘Babovaca’ | admx | 2 | GO |
| 3 | ‘Bascenka’ | 1 | admx | GO |
| 4 | ‘Bihorka’ | 1 | admx | GO |
| 5 | ‘Bistrica’ | admx | admx | GO |
| 6 | ‘Bobovec’ | admx | 3 | SR |
| 7 | ‘Bukovija’ | 1 | 2 | SR |
| 8 | ‘Butulija’ | 1 | admx | GO |
| 9 | ‘Crvenka’ | 2 | 3 | GO |
| 10 | ‘Djulabija’ | 1 | 3 | SR |
| 11 | ‘Dobric’ | 1 | 2 | SR |
| 12 | ‘Dobrinjka’ | 2 | admx | GO |
| 13 | ‘Habikusa’ | 1 | 2 | SR |
| 14 | ‘Lederka’ | admx | admx | SR |
| 15 | ‘Lijepocvjetka’ | 2 | 1 | SR |
| 16 | ‘Misirka’ | 1 | 3 | GO |
| 17 | ‘Muslimaca’ | 1 | admx | GO |
| 18 | ‘Pamuklija’ | 1 | admx | SR |
| 19 | ‘Paradija’ | admx | 2 | SR |
| 20 | ‘Petrovaca bijela’ | admx | 3 | SR |
| 21 | ‘Posavka’ | 1 | 3 | GO |
| 22 | ‘Prisatka’ | admx | admx | GO |
| 23 | ‘Ranka’ | 1 | 2 | GO |
| 24 | ‘Rebraca’ | 2 | admx | SR |
| 25 | ‘Ruzmarinka’ | 1 | 3 | SR |
| 26 | ‘Samoniklica’ | 1 | 3 | SR |
| 27 | ‘Sarija’ | admx | admx | SR |
| 28 | ‘Senabija’ | 1 | 3 | SR |
| 29 | ‘Simsirka’ | 1 | 2 | GO |
| 30 | ‘Srcika’ | 2 | admx | GO |
| 31 | ‘Srebrenicka’ | 1 | 2 | SR |
| 32 | ‘Sulija’ | 1 | admx | GO |
| 33 | ‘Zuja’ | 1 | 3 | SR |
| International reference cultivars | ||||
| 1 | ‘Braeburn’ | 2 | 1 | SR |
| 2 | ‘Elstar’ | 2 | 1 | SR |
| 3 | ‘Fuji Nagafu’ | 2 | 1 | SR |
| 4 | ‘Gala Galaxy’ | 2 | 1 | SR |
| 5 | ‘Granny Smith’ | 2 | admx | SR |
| 6 | ‘Melrose’ | 2 | 1 | SR |
| 7 | ‘Pilot’ | 2 | 1 | SR |
| 8 | ‘Pink Lady’ | 2 | 1 | SR |
| 9 | ‘Pinova’ | 2 | 1 | SR |
| 10 | ‘Piros’ | admx | 3 | SR |
| 11 | ‘Golden Reinders’ | 2 | 1 | SR |
| 12 | ‘Topaz’ | 2 | 1 | SR |
| N | MAF | Ho | He | F | |
|---|---|---|---|---|---|
| Pruned SNP data | 263,406 | 0.24 | 0.29 | 0.33 | 0.11 |
| Non-pruned SNP data | 487,163 | 0.25 | 0.31 | 0.34 | 0.07 |
| Locus | N | Na | Ne | I | Ho | He | uHe | F |
|---|---|---|---|---|---|---|---|---|
| CH01H01 | 45 | 10 | 7.09 | 2.04 | 0.87 | 0.86 | 0.87 | −0.01 |
| CH05E03 | 45 | 15 | 7.61 | 2.34 | 0.6 | 0.87 | 0.88 | 0.31 |
| CH05E04 | 45 | 12 | 5.39 | 1.99 | 0.73 | 0.81 | 0.82 | 0.1 |
| CH01H02 | 5 | 9 | 3.96 | 1.63 | 0.71 | 0.75 | 0.76 | 0.05 |
| CH02C02a | 45 | 19 | 12.02 | 2.67 | 0.91 | 0.92 | 0.93 | 0.01 |
| CH04E02 | 45 | 9 | 2.32 | 1.29 | 0.53 | 0.57 | 0.58 | 0.06 |
| CH01H10 | 45 | 13 | 4.51 | 1.89 | 0.8 | 0.78 | 0.79 | −0.03 |
| CH02D08 | 45 | 11 | 6 | 2.03 | 0.87 | 0.83 | 0.84 | −0.04 |
| CH02C02b | 45 | 5 | 2.32 | 0.99 | 0.49 | 0.57 | 0.58 | 0.14 |
| CH02C06 | 45 | 14 | 9.35 | 2.41 | 0.89 | 0.89 | 0.9 | 0.01 |
| Average | 11.7 | 6.06 | 1.93 | 0.74 | 0.79 | 0.8 | 0.06 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Konjić, A.; Kurtović, M.; Grahić, J.; Pojskić, N.; Kalajdžić, A.; Gaši, F. Using High-Density SNP Array to Investigate Genetic Relationships and Structure of Apple Germplasm in Bosnia and Herzegovina. Horticulturae 2023, 9, 527. https://doi.org/10.3390/horticulturae9050527
Konjić A, Kurtović M, Grahić J, Pojskić N, Kalajdžić A, Gaši F. Using High-Density SNP Array to Investigate Genetic Relationships and Structure of Apple Germplasm in Bosnia and Herzegovina. Horticulturae. 2023; 9(5):527. https://doi.org/10.3390/horticulturae9050527
Chicago/Turabian StyleKonjić, Almira, Mirsad Kurtović, Jasmin Grahić, Naris Pojskić, Abdurahim Kalajdžić, and Fuad Gaši. 2023. "Using High-Density SNP Array to Investigate Genetic Relationships and Structure of Apple Germplasm in Bosnia and Herzegovina" Horticulturae 9, no. 5: 527. https://doi.org/10.3390/horticulturae9050527