
Morphological, Phytochemical, and Photosynthetic Performance of Grafted Tomato Seedlings in Response to Different LED Light Qualities under Protected Cultivation
 ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
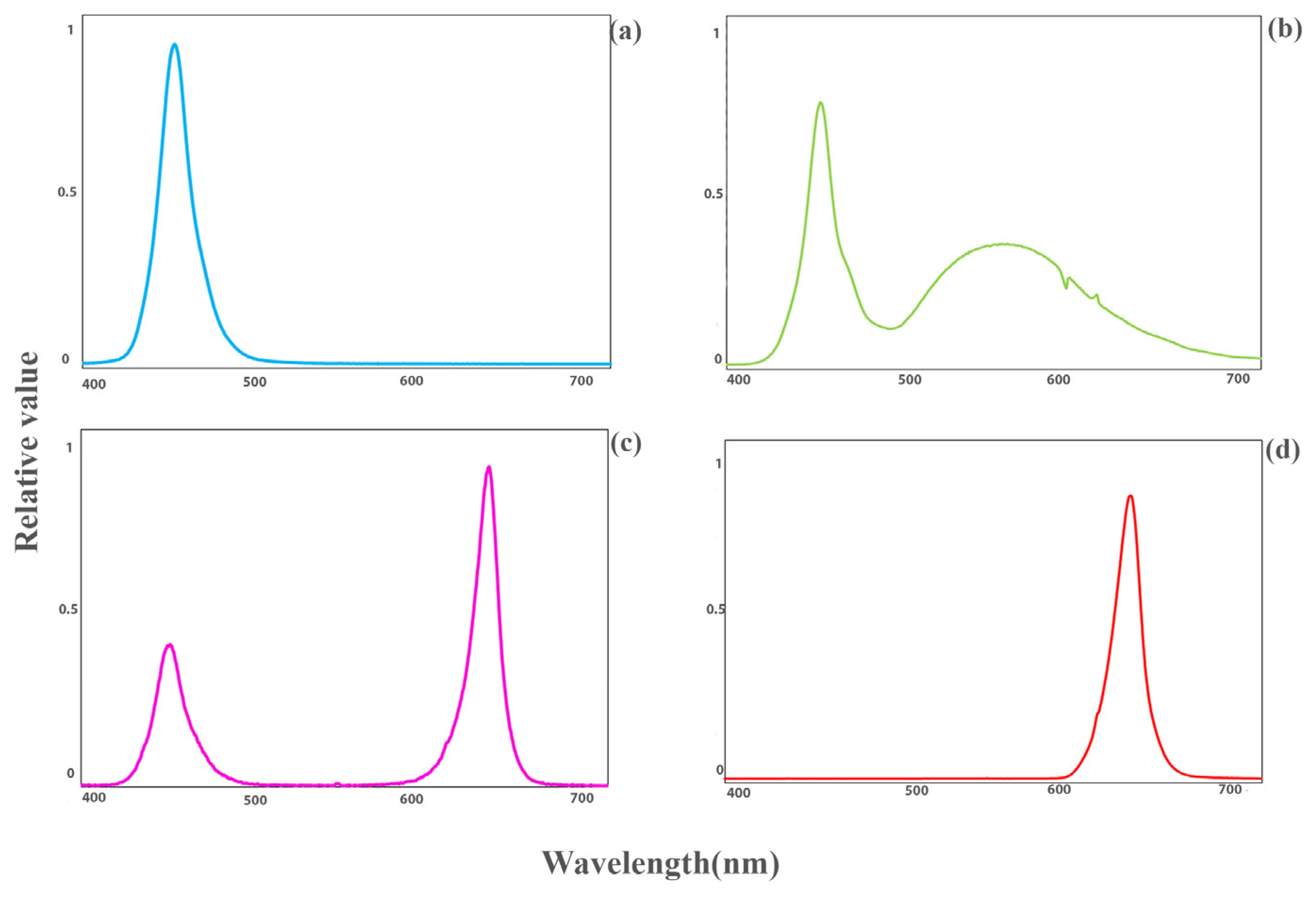
2.1. Plant Materials and Environmental Conditions
2.2. Morphological Parameters
2.3. Determination of Soluble Carbohydrates
2.4. Antioxidant Enzymes Activity
2.5. Determination of Hydrogen Peroxide Content
2.6. SPAD Determination
2.7. Chlorophyll Fluorescence Imaging
2.8. Analysis of the OJIP Test
2.9. Statistical Analysis
3. Results
3.1. Morphology of Grafted Seedlings Was Improved by RB Light Regime
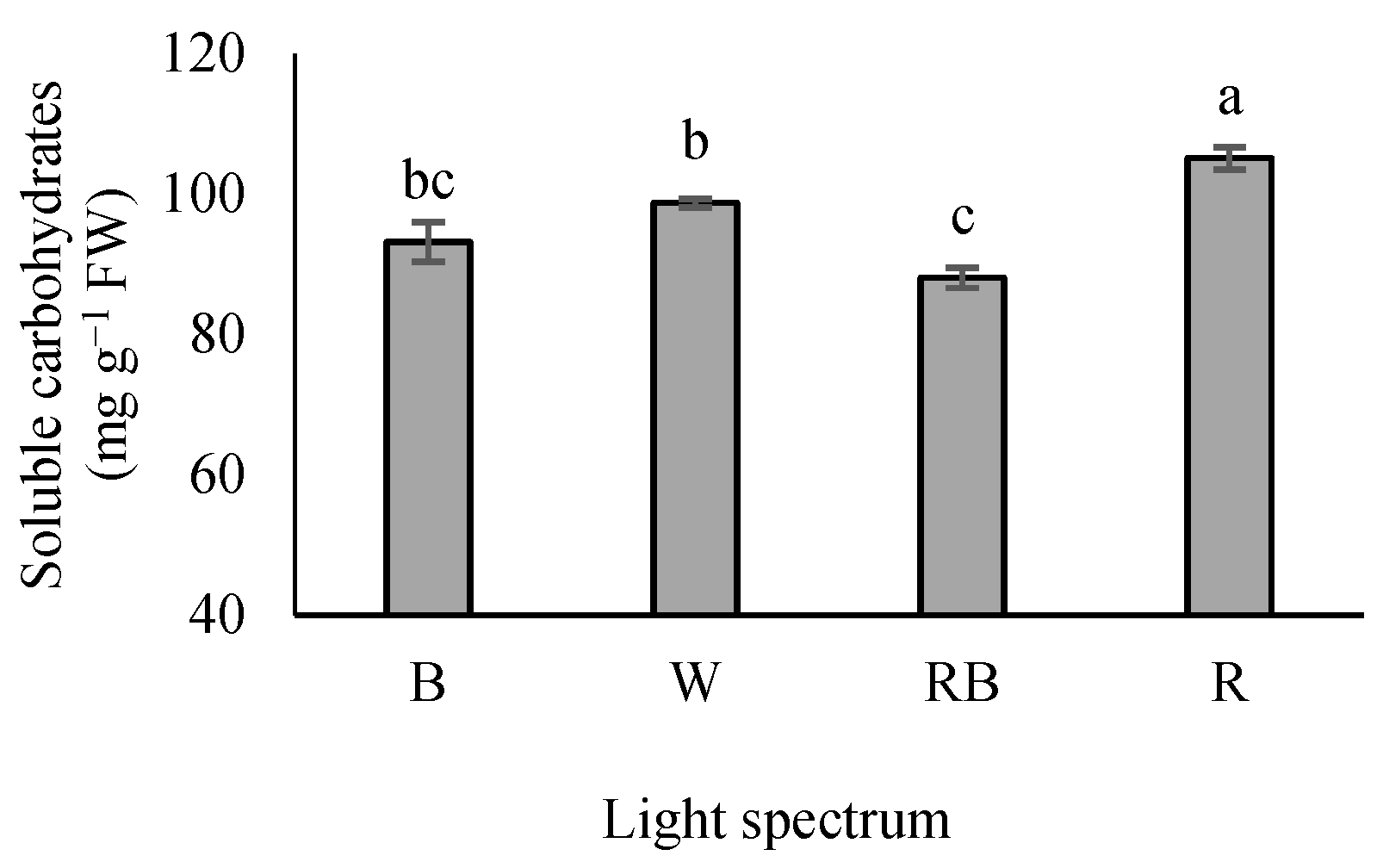
3.2. Leaf-Soluble Carbohydrates
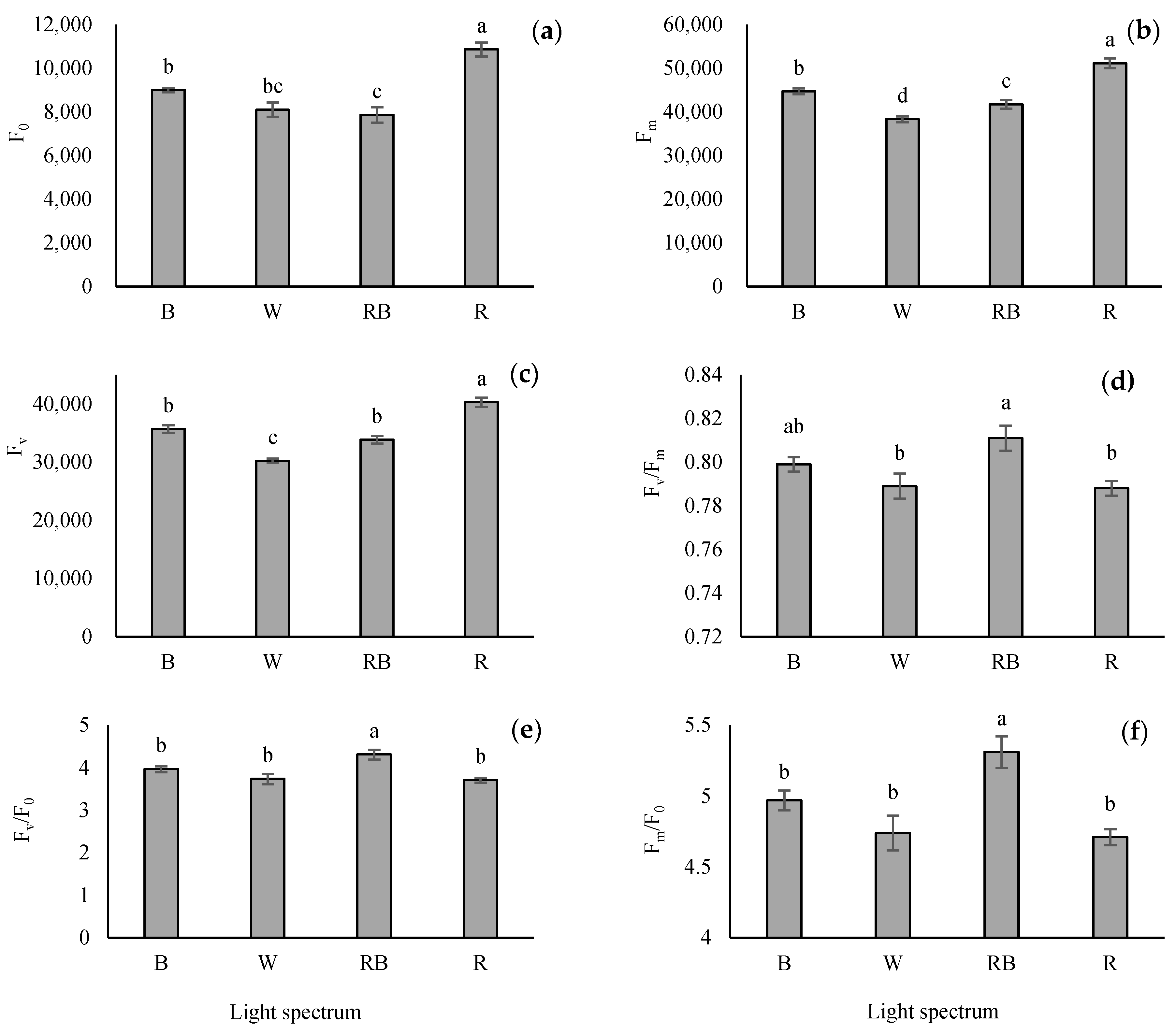
3.3. Chlorophyll Fluorescence Imaging
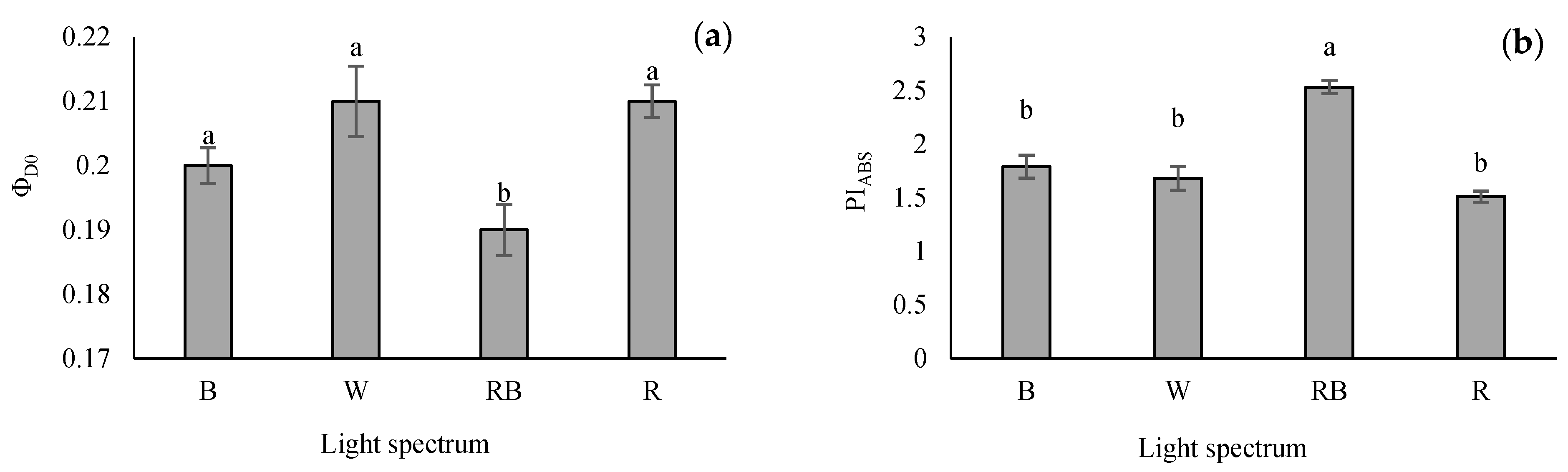
3.4. Evaluation of Parameters Obtained from the OJIP Protocol
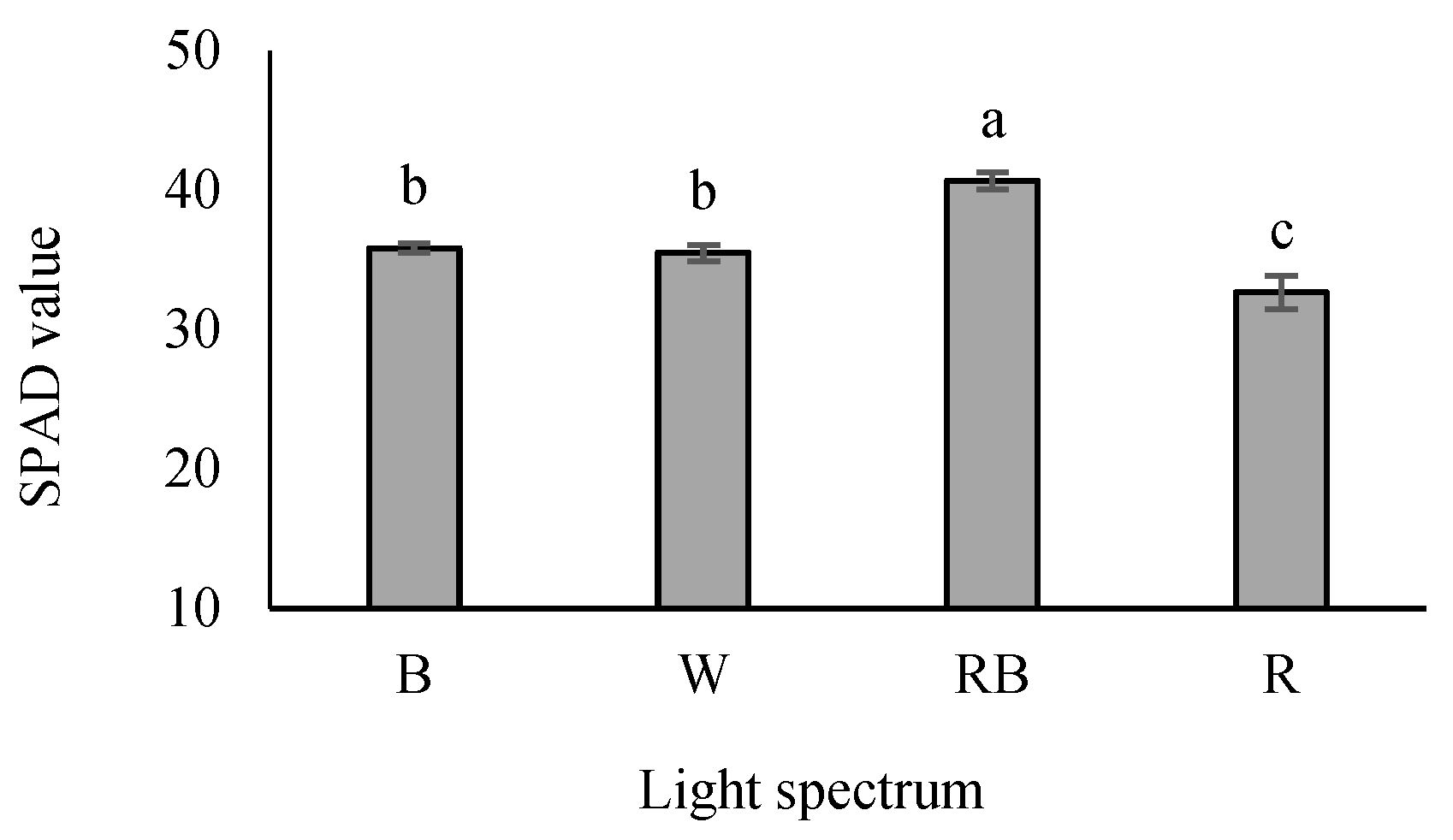
3.5. Chlorophyll Index
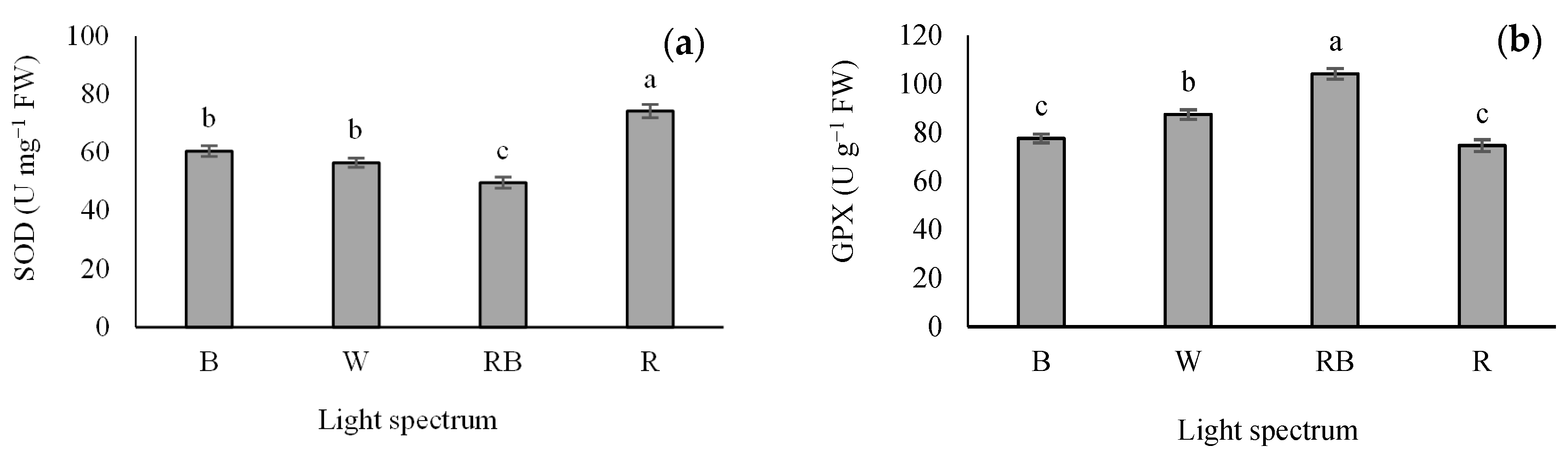
3.6. Antioxidant Enzymes Activity
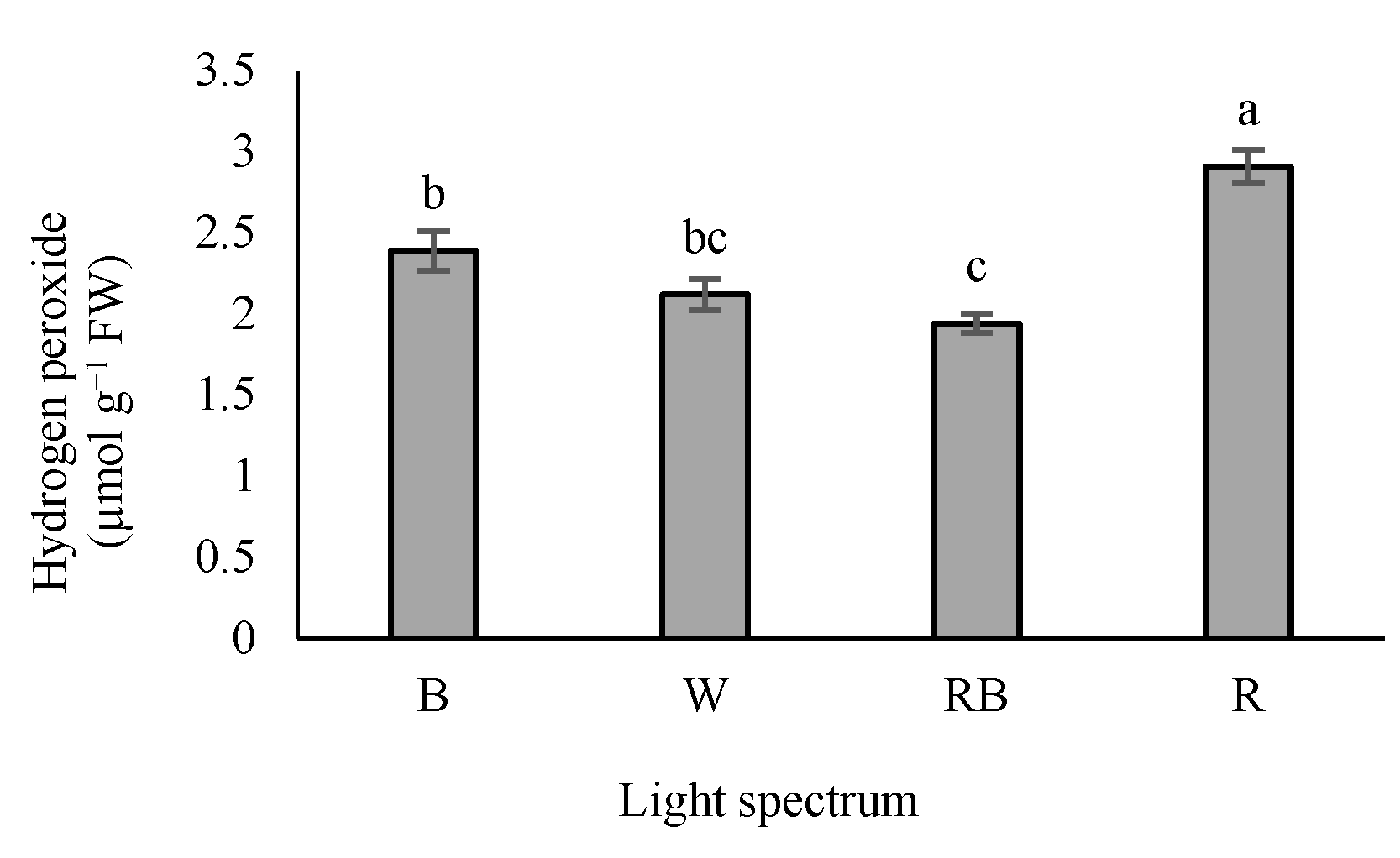
3.7. Determination of Hydrogen Peroxide
4. Discussion
4.1. Plant Fresh and Dry Weight and Stem Diameter Reduced in R Light
4.2. Red Light-Induced Carbohydrate Accumulation
4.3. Red Light-Induced SOD Enzymes Activity and H2O2 Accumulation
4.4. Photosynthetic Functionality of Tomato Grafted Seedlings Enhanced by RB Light
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| B | blue |
| R | red |
| RB | red and blue |
| W | white |
| D | darkness |
| OJIP | polyphasic chlorophyll fluorescence induction curve |
| FW | Fresh weight |
| PPFD | photosynthetic photon flux density |
| PSII | photosystem II |
| SOD | superoxide dismutase |
| GPX | guaiacol peroxidase |
| H2O2 | hydrogen peroxide |
| RC | reaction center |
| NBT | nitro blue tetrazolium |
| KI | potassium iodide |
| TCA | Trichloroacetic acid |
References
- King, S.R.; Davis, A.R.; Zhang, X.; Crosby, K. Genetics, Breeding and Selection of Rootstocks for Solanaceae and Cucurbitaceae. Sci. Hortic. 2010, 127, 106–111. [Google Scholar] [CrossRef]
- Rouphael, Y.; Kyriacou, M.C.; Colla, G. Vegetable Grafting: A Toolbox for Securing Yield Stability under Multiple Stress Conditions. Front. Plant Sci. 2018, 8, 2255. [Google Scholar] [CrossRef] [Green Version]
- Gruda, N.; Savvas, D.; Colla, G.; Rouphael, Y. Impacts of Genetic Material and Current Technologies on Product Quality of Selected Greenhouse Vegetables—A Review. Eur. J. Hortic. Sci. 2018, 83, 319–328. [Google Scholar] [CrossRef]
- Bantis, F.; Koukounaras, A.; Siomos, A.; Menexes, G.; Dangitsis, C.; Kintzonidis, D. Assessing Quantitative Criteria for Characterization of Quality Categories for Grafted Watermelon Seedlings. Horticulturae 2019, 5, 16. [Google Scholar] [CrossRef] [Green Version]
- Abdelmageed, A.H.A.; Gruda, N. Influence of Grafting on Growth, Development and Some Physiological Parameters of Tomatoes under Controlled Heat Stress Conditions. Eur. J. Hortic. Sci. 2009, 74, 16–20. [Google Scholar]
- Colla, G.; Pérez-Alfocea, F.; Schwarz, D. Vegetable Grafting: Principles and Practices; CAB International: Oxfordshire, UK, 2017; ISBN 9781780648972. [Google Scholar]
- Moosavi-Nezhad, M.; Salehi, R.; Aliniaeifard, S.; Winans, K.S.; Nabavi-Pelesaraei, A. An Analysis of Energy Use and Economic and Environmental Impacts in Conventional Tunnel and LED-Equipped Vertical Systems in Healing and Acclimatization of Grafted Watermelon Seedlings. J. Clean. Prod. 2022, 361, 132069. [Google Scholar] [CrossRef]
- Bantis, F.; Koukounaras, A.; Siomos, A.S.; Fotelli, M.N.; Kintzonidis, D. Bichromatic Red and Blue LEDs during Healing Enhance the Vegetative Growth and Quality of Grafted Watermelon Seedlings. Sci. Hortic. 2020, 261, 109000. [Google Scholar] [CrossRef]
- Dong, W.; Zhou, Z.C.; Bu, Y.L.; Zhuo, J.Q.; Chen, L.Z.; Li, Y.Z. Research and Application of Grafted Seedlings Healing Room. Acta Hortic. 2015, 1086, 51–58. [Google Scholar] [CrossRef]
- Lee, K.M.; Lim, C.S.; Muneer, S.; Jeong, B.R. Functional Vascular Connections and Light Quality Effects on Tomato Grafted Unions. Sci. Hortic. 2016, 201, 306–317. [Google Scholar] [CrossRef]
- Lang, K.M.; Nair, A.; Litvin, A.G. An Alternative Healing Method for Grafted Tomato Transplants: The Effect of Light Exclusion and Substrate Temperature on Plant Survival and Growth. Horttechnology 2020, 30, 677–684. [Google Scholar] [CrossRef]
- Klem, K.; Gargallo-Garriga, A.; Rattanapichai, W.; Oravec, M.; Holub, P.; Veselá, B.; Sardans, J.; Peñuelas, J.; Urban, O. Distinct Morphological, Physiological, and Biochemical Responses to Light Quality in Barley Leaves and Roots. Front. Plant Sci. 2019, 10, 1026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huber, M.; Nieuwendijk, N.M.; Pantazopoulou, C.K.; Pierik, R. Light Signalling Shapes Plant–Plant Interactions in Dense Canopies. Plant Cell Environ. 2021, 44, 1014–1029. [Google Scholar] [CrossRef]
- Fu, W.; Li, P.; Wu, Y. Effects of Different Light Intensities on Chlorophyll Fluorescence Characteristics and Yield in Lettuce. Sci. Hortic. 2012, 135, 45–51. [Google Scholar] [CrossRef]
- Kozai, T.; Niu, G.; Takagaki, M. Plant Factory: An Indoor Vertical Farming System for Efficient Quality Food Production, 2nd ed.; Academic Press: San Diego, CA, USA, 2019; ISBN 9780128166918. [Google Scholar]
- Li, H.; Tang, C.; Xu, Z. The Effects of Different Light Qualities on Rapeseed (Brassica napus L.) Plantlet Growth and Morphogenesis in Vitro. Sci. Hortic. 2013, 150, 117–124. [Google Scholar] [CrossRef]
- Taulavuori, K.; Pyysalo, A.; Taulavuori, E.; Julkunen-Tiitto, R. Responses of Phenolic Acid and Flavonoid Synthesis to Blue and Blue-Violet Light Depends on Plant Species. Environ. Exp. Bot. 2018, 150, 183–187. [Google Scholar] [CrossRef] [Green Version]
- Aliniaeifard, S.; Hajilou, J.; Tabatabaei, S.J. Photosynthetic and Growth Responses of Olive to Proline and Salicylic Acid under Salinity Condition. Not. Bot. Horti Agrobot. Cluj-Napoca 2016, 44, 579–585. [Google Scholar] [CrossRef] [Green Version]
- Moosavi-Nezhad, M.; Salehi, R.; Aliniaeifard, S.; Tsaniklidis, G.; Woltering, E.J.; Fanourakis, D.; Żuk-Gołaszewska, K.; Kalaji, H.M. Blue Light Improves Photosynthetic Performance during Healing and Acclimatization of Grafted Watermelon Seedlings. Int. J. Mol. Sci. 2021, 22, 8043. [Google Scholar] [CrossRef]
- Viršile, A.; Olle, M.; Duchovskis, P. LED Lighting in Horticulture. In Light Emitting Diodes for Agriculture: Smart Lighting; Springer Singapore: Singapore, 2017; pp. 113–147. ISBN 9789811058073. [Google Scholar]
- Meiramkulova, K.; Tanybayeva, Z.; Kydyrbekova, A.; Turbekova, A.; Aytkhozhin, S.; Zhantasov, S.; Taukenov, A. The Efficiency of Led Irradiation for Cultivating High-Quality Tomato Seedlings. Sustainability 2021, 13, 9426. [Google Scholar] [CrossRef]
- Seif, M.; Aliniaeifard, S.; Arab, M.; Mehrjerdi, M.Z.; Shomali, A.; Fanourakis, D.; Li, T.; Woltering, E. Monochromatic Red Light during Plant Growth Decreases the Size and Improves the Functionality of Stomata in Chrysanthemum. Funct. Plant Biol. 2021, 48, 515–528. [Google Scholar] [CrossRef]
- Chen, Y.; Zhou, B.; Li, J.; Tang, H.; Tang, J.; Yang, Z. Formation and Change of Chloroplast-Located Plant Metabolites in Response to Light Conditions. Int. J. Mol. Sci. 2018, 19, 654. [Google Scholar] [CrossRef] [Green Version]
- Shin, K.S.; Murthy, H.N.; Heo, J.W.; Hahn, E.J.; Paek, K.Y. The Effect of Light Quality on the Growth and Development of in Vitro Cultured Doritaenopsis Plants. Acta Physiol. Plant. 2008, 30, 339–343. [Google Scholar] [CrossRef]
- Li, Y.; Xin, G.; Wei, M.; Shi, Q.; Yang, F.; Wang, X. Carbohydrate Accumulation and Sucrose Metabolism Responses in Tomato Seedling Leaves When Subjected to Different Light Qualities. Sci. Hortic. 2017, 225, 490–497. [Google Scholar] [CrossRef]
- Whitelam, G.C.; Halliday, K.J. Light and Plant Development; Garry, C., Whitelam, K.J.H., Eds.; Wiley: Hoboken, NJ, USA, 2007; ISBN 9780470988893. [Google Scholar]
- Strasser, R.J.; Tsimilli-Michael, M.; Srivastava, A. Analysis of the Chlorophyll a Fluorescence Transient. In Chlorophyll a Fluorescence: A Signature of Photosynthesis. Advances in Photosynthesis and Respiration; Papageorgiou, G., Govindjee, Eds.; Springer: Dordrecht, The Netherlands, 2004; pp. 321–362. [Google Scholar]
- Kalaji, H.M.; Govindjee; Bosa, K.; Kościelniak, J.; Zuk-Gołaszewska, K. Effects of Salt Stress on Photosystem II Efficiency and CO2 Assimilation of Two Syrian Barley Landraces. Environ. Exp. Bot. 2011, 73, 64–72. [Google Scholar] [CrossRef]
- Strasser, R.J.; Srivastava, A.; Tsimilli-Michael, M. The Fluorescence Transient as a Tool to Characterize and Screen Photosynthetic Samples. In Probing Photosynthesis: Mechanism, Regulation & Adaptation; Taylor and Francis: London, UK, 2000; pp. 443–480. ISBN 0748408215. [Google Scholar]
- Kalaji, H.M.; Jajoo, A.; Oukarroum, A.; Brestic, M.; Zivcak, M.; Samborska, I.A.; Cetner, M.D.; Łukasik, I.; Goltsev, V.; Ladle, R.J. Chlorophyll a Fluorescence as a Tool to Monitor Physiological Status of Plants under Abiotic Stress Conditions. Acta Physiol. Plant. 2016, 38, 102. [Google Scholar] [CrossRef] [Green Version]
- Rapacz, M.; Sasal, M.; Kalaji, H.M.; Kościelniak, J. Is the OJIP Test a Reliable Indicator of Winter Hardiness and Freezing Tolerance of Common Wheat and Triticale under Variable Winter Environments? PLoS ONE 2015, 10, e0134820. [Google Scholar] [CrossRef] [Green Version]
- Kinoshita, T.; Doi, M.; Suetsugu, N.; Kagawa, T.; Wada, M.; Shimazaki, K.I. Phot1 and Phot2 Mediate Blue Light Regulation of Stomatal Opening. Nature 2001, 414, 656–660. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Ballesta, M.C.; Alcaraz-López, C.; Muries, B.; Mota-Cadenas, C.; Carvajal, M. Physiological Aspects of Rootstock-Scion Interactions. Sci. Hortic. 2010, 127, 112–118. [Google Scholar] [CrossRef]
- Estaji, A.; Kalaji, H.M.; Karimi, H.R.; Roosta, H.R.; Moosavi-Nezhad, S.M. How Glycine Betaine Induces Tolerance of Cucumber Plants to Salinity Stress? Photosynthetica 2019, 57, 753–761. [Google Scholar] [CrossRef] [Green Version]
- Moosavi-Nezhad, M.; Alibeigi, B.; Estaji, A.; Gruda, N.S.; Aliniaeifard, S. Growth, Biomass Partitioning, and Photosynthetic Performance of Chrysanthemum Cuttings in Response to Different Light Spectra. Plants 2022, 11, 3337. [Google Scholar] [CrossRef] [PubMed]
- Van Doorn, W.G. Water Relations of Cut Flowers: An Update. In Horticultural Reviews; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2012; Volume 40, pp. 55–106. [Google Scholar]
- Dhindsa, R.S.; Plumb-dhindsa, P.; Thorpe, T.A. Leaf Senescence: Correlated with Increased Levels of Membrane Permeability and Lipid Peroxidation, and Decreased Levels of Superoxide Dismutase and Catalase. J. Exp. Bot. 1981, 32, 93–101. [Google Scholar] [CrossRef]
- Flocco, C.G.; Giulietti, A.M. In Vitro Hairy Root Cultures as a Tool for Phytoremediation Research. In Phytoremediation: Methods and Reviews; Springer: Berlin/Heidelberg, Germany, 2007; pp. 161–173. [Google Scholar]
- Patterson, B.D.; MacRae, E.A.; Ferguson, I.B. Estimation of Hydrogen Peroxide in Plant Extracts Using Titanium(IV). Anal. Biochem. 1984, 139, 487–492. [Google Scholar] [CrossRef] [PubMed]
- Hoel, B.O.; Solhaug, K.A. Effect of Irradiance on Chlorophyll Estimation with the Minolta SPAD-502 Leaf Chlorophyll Meter. Ann. Bot. 1998, 82, 389–392. [Google Scholar] [CrossRef] [Green Version]
- Aliniaeifard, S.; Van Meeteren, U. Natural Variation in Stomatal Response to Closing Stimuli among Arabidopsis Thaliana Accessions after Exposure to Low VPD as a Tool to Recognize the Mechanism of Disturbed Stomatal Functioning. J. Exp. Bot. 2014, 65, 6529–6542. [Google Scholar] [CrossRef]
- Ashrostaghi, T.; Aliniaeifard, S.; Shomali, A.; Azizinia, S.; Koohpalekani, J.A.; Moosavi-Nezhad, M.; Gruda, N.S. Light Intensity: The Role Player in Cucumber Response to Cold Stress. Agronomy 2022, 12, 201. [Google Scholar] [CrossRef]
- Moradi, S.; Kafi, M.; Aliniaeifard, S.; Moosavi-Nezhad, M.; Pedersen, C.; Gruda, N.S.; Salami, S.A. Monochromatic Blue Light Enhances Crocin and Picrocrocin Content by Upregulating the Expression of Underlying Biosynthetic Pathway Genes in Saffron (Crocus sativus L.). Front. Hortic. 2022, 1, 960423. [Google Scholar] [CrossRef]
- Gorbe, E.; Calatayud, A. Applications of Chlorophyll Fluorescence Imaging Technique in Horticultural Research: A Review. Sci. Hortic. 2012, 138, 24–35. [Google Scholar] [CrossRef]
- Stirbet, A.; Lazár, D.; Kromdijk, J. Govindjee Chlorophyll a Fluorescence Induction: Can Just a One-Second Measurement Be Used to Quantify Abiotic Stress Responses? Photosynthetica 2018, 56, 86–104. [Google Scholar] [CrossRef]
- Kalaji, M.H.; Goltsev, V.N.; Żuk-Golaszewska, K.; Zivcak, M.; Brestic, M. Chlorophyll Fluorescence: Understanding Crop Performance—Basics and Applications; CRC Press: Boca Raton, FL, USA, 2017; ISBN 9781498764506. [Google Scholar]
- Yu, X.; Liu, H.; Klejnot, J.; Lin, C. The Cryptochrome Blue Light Receptors. Arab. Book 2010, 8, e0135. [Google Scholar] [CrossRef] [Green Version]
- Yousef, A.F.; Ali, M.M.; Rizwan, H.M.; Ahmed, M.A.A.; Ali, W.M.; Kalaji, H.M.; Elsheery, N.; Wróbel, J.; Xu, Y.; Chen, F. Effects of Light Spectrum on Morphophysiological Traits of Grafted Tomato Seedlings. PLoS ONE 2021, 16, e0250210. [Google Scholar] [CrossRef]
- Nguyen, D.T.P.; Kitayama, M.; Lu, N.; Takagaki, M. Improving Secondary Metabolite Accumulation, Mineral Content, and Growth of Coriander (Coriandrum sativum L.) by Regulating Light Quality in a Plant Factory. J. Hortic. Sci. Biotechnol. 2020, 95, 356–363. [Google Scholar] [CrossRef]
- Neff, M.M.; Fankhauser, C.; Chory, J. Eight: An Indicator of Time and Place. Genes Dev. 2000, 14, 257–271. [Google Scholar] [CrossRef]
- Yang, F.O.; Mao, J.F.; Wang, J.; Zhang, S.; Li, Y. Transcriptome Analysis Reveals That Red and Blue Light Regulate Growth and Phytohormone Metabolism in Norway Spruce [Picea abies (L.) Karst.] . PLoS ONE 2015, 10, e0127896. [Google Scholar] [CrossRef] [Green Version]
- Dierck, R.; Dhooghe, E.; Van Huylenbroeck, J.; Van Der Straeten, D.; De Keyser, E. Light Quality Regulates Plant Architecture in Different Genotypes of Chrysanthemum Morifolium Ramat. Sci. Hortic. 2017, 218, 177–186. [Google Scholar] [CrossRef]
- Fang, L.; Ma, Z.; Wang, Q.; Nian, H.; Ma, Q.; Huang, Q.; Mu, Y. Plant Growth and Photosynthetic Characteristics of Soybean Seedlings Under Different LED Lighting Quality Conditions. J. Plant Growth Regul. 2021, 40, 668–678. [Google Scholar] [CrossRef]
- Lau, O.S.; Deng, X.W. Plant Hormone Signaling Lightens up: Integrators of Light and Hormones. Curr. Opin. Plant Biol. 2010, 13, 571–577. [Google Scholar] [CrossRef]
- El Kayal, W.; Allen, C.C.G.; Ju, C.J.T.; Adams, E.; King-Jones, S.; Zaharia, L.I.; Abrams, S.R.; Cooke, J.E.K. Molecular Events of Apical Bud Formation in White Spruce, Picea Glauca. Plant Cell Environ. 2011, 34, 480–500. [Google Scholar] [CrossRef] [Green Version]
- Kurepin, L.V.; Walton, L.J.; Hayward, A.; Emery, R.J.N.; Pharis, R.P.; Reid, D.M. Interactions between Plant Hormones and Light Quality Signaling in Regulating the Shoot Growth of Arabidopsis Thaliana Seedlings. Botany 2012, 90, 237–246. [Google Scholar] [CrossRef]
- Li, Y.; Liu, Z.; Shi, Q.; Yang, F.; Wei, M. Mixed Red and Blue Light Promotes Tomato Seedlings Growth by Influencing Leaf Anatomy, Photosynthesis, CO2 Assimilation and Endogenous Hormones. Sci. Hortic. 2021, 290, 110500. [Google Scholar] [CrossRef]
- Ouzounis, T.; Fretté, X.; Rosenqvist, E.; Ottosen, C.O. Spectral Effects of Supplementary Lighting on the Secondary Metabolites in Roses, Chrysanthemums, and Campanulas. J. Plant Physiol. 2014, 171, 1491–1499. [Google Scholar] [CrossRef] [PubMed]
- Xiao-ying, L.; Zhi-gang, X.; Tao-tao, C.; Shi-rong, G. Growth and Photosynthesis of Cherry Tomato Seedling Exposed to Different Low Light of LED Light Quality. Acta Bot. Boreali-Occident. Sincia 2010, 30, 725–732. [Google Scholar]
- Chen, C.C.; Huang, M.Y.; Lin, K.H.; Wong, S.L.; Huang, W.D.; Yang, C.M. Effects of Light Quality on the Growth, Development and Metabolism of Rice Seedlings (Oryza sativa L.). Res. J. Biotechnol. 2014, 9, 15–24. [Google Scholar]
- Wang, L.; Chen, X.; Wang, Q.; Hao, J.; Lan, J. Effect of Different Light of LED Light Quality on Growth and Antioxidant Enzyme Activities of Ganoderma Lucidum. Zhongguo Zhongyao Zazhi 2011, 36, 2471–2474. [Google Scholar] [CrossRef]
- Kaiser, E.; Ouzounis, T.; Giday, H.; Schipper, R.; Heuvelink, E.; Marcelis, L.F.M. Adding Blue to Red Supplemental Light Increases Biomass and Yield of Greenhouse-Grown Tomatoes, but Only to an Optimum. Front. Plant Sci. 2019, 9, 2002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, S.; Liu, X.; Liu, Y.; Cao, B.; Chen, Z.; Xu, K. Photosynthetic Characteristics and Chloroplast Ultrastructure of Welsh Onion (Allium fistulosum L.) Grown under Different LED Wavelengths. BMC Plant Biol. 2020, 20, 78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, L.; Hong, Y.; Zhang, X. Effect of Light Quality on Calluses Induction and Differentiation of Capsicum Annuum. J. Hunan Agric. Univ. 2009, 35, 615–617. [Google Scholar]
- Vu, N.T.; Kim, Y.S.; Kang, H.M.; Kim, I.S. Influence of Short-Term Irradiation during Pre- and Post-Grafting Period on the Graft-Take Ratio and Quality of Tomato Seedlings. Hortic. Environ. Biotechnol. 2014, 55, 27–35. [Google Scholar] [CrossRef]
- Zavala, J.A.; Ravetta, D.A. Allocation of Photoassimilates to Biomass, Resin and Carbohydrates in Grindelia Chiloensis as Affected by Light Intensity. Field Crops Res. 2001, 69, 143–149. [Google Scholar] [CrossRef]
- Smith, A.M.; Stitt, M. Coordination of Carbon Supply and Plant Growth. Plant Cell Environ. 2007, 30, 1126–1149. [Google Scholar] [CrossRef]
- Fan, J.; Yu, L.; Xu, C. A Central Role for Triacylglycerol in Membrane Lipid Breakdown, Fatty Acid β-Oxidation, and Plant Survival under Extended Darkness. Plant Physiol. 2017, 174, 1517–1530. [Google Scholar] [CrossRef] [Green Version]
- Usadel, B.; Bläsing, O.E.; Gibon, Y.; Poree, F.; Höhne, M.; Günter, M.; Trethewey, R.; Kamlage, B.; Poorter, H.; Stitt, M. Multilevel Genomic Analysis of the Response of Transcripts, Enzyme Activities and Metabolites in Arabidopsis Rosettes to a Progressive Decrease of Temperature in the Non-Freezing Range. Plant Cell Environ. 2008, 31, 518–547. [Google Scholar] [CrossRef]
- Pham, M.D.; Hwang, H.; Park, S.W.; Cui, M.; Lee, H.; Chun, C. Leaf Chlorosis, Epinasty, Carbohydrate Contents and Growth of Tomato Show Different Responses to the Red/Blue Wavelength Ratio under Continuous Light. Plant Physiol. Biochem. 2019, 141, 477–486. [Google Scholar] [CrossRef]
- Sæbø, A.; Krekling, T.; Appelgren, M. Light Quality Affects Photosynthesis and Leaf Anatomy of Birch Plantlets in Vitro. Plant Cell Tissue Organ Cult. 1995, 41, 177–185. [Google Scholar] [CrossRef]
- Jeannette, E.; Reyss, A.; Grégory, N.; Gantet, P.; Prioul, J.L. Carbohydrate Metabolism in a Heat-Girdled Maize Source Leaf. Plant Cell Environ. 2000, 23, 61–69. [Google Scholar] [CrossRef]
- Wang, H.; Gu, M.; Cui, J.; Shi, K.; Zhou, Y.; Yu, J. Effects of Light Quality on CO2 Assimilation, Chlorophyll-Fluorescence Quenching, Expression of Calvin Cycle Genes and Carbohydrate Accumulation in Cucumis Sativus. J. Photochem. Photobiol. B Biol. 2009, 96, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Bhandari, R.; Sharma, P.K. High-Light–Induced Changes on Photosynthesis, Pigments, Sugars, Lipids and Antioxidant Enzymes in Freshwater (Nostoc Spongiaeforme) and Marine (Phormidium Corium) Cyanobacteria. Photochem. Photobiol. 2006, 82, 702–710. [Google Scholar] [CrossRef]
- Distelbarth, H.; Nägele, T.; Heyer, A.G. Responses of Antioxidant Enzymes to Cold and High Light Are Not Correlated to Freezing Tolerance in Natural Accessions of Arabidopsis Thaliana. Plant Biol. 2013, 15, 982–990. [Google Scholar] [CrossRef]
- Xu, F.; Shi, L.; Chen, W.; Cao, S.; Su, X.; Yang, Z. Effect of Blue Light Treatment on Fruit Quality, Antioxidant Enzymes and Radical-Scavenging Activity in Strawberry Fruit. Sci. Hortic. 2014, 175, 181–186. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Zulfiqar, F.; Raza, A.; Mohsin, S.M.; Al Mahmud, J.; Fujita, M.; Fotopoulos, V. Reactive Oxygen Species and Antioxidant Defense in Plants under Abiotic Stress: Revisiting the Crucial Role of a Universal Defense Regulator. Antioxidants 2020, 9, 681. [Google Scholar] [CrossRef]
- Bayat, L.; Arab, M.; Aliniaeifard, S.; Seif, M.; Lastochkina, O.; Li, T. Effects of Growth under Different Light Spectra on the Subsequent High Light Tolerance in Rose Plants. AoB Plants 2018, 10, ply052. [Google Scholar] [CrossRef]
- Lee, T.Y.; Woo, S.Y.; Kwak, M.J.; Inkyin, K.; Lee, K.E.; Jang, J.H.; Kim, I.R. Photosynthesis and Chlorophyll Fluorescence Responses of Populus Sibirica to Water Deficit in a Desertification Area in Mongolia. Photosynthetica 2016, 54, 317–320. [Google Scholar] [CrossRef]
- Falqueto, A.R.; da Silva Júnior, R.A.; Gomes, M.T.G.; Martins, J.P.R.; Silva, D.M.; Partelli, F.L. Effects of Drought Stress on Chlorophyll a Fluorescence in Two Rubber Tree Clones. Sci. Hortic. 2017, 224, 238–243. [Google Scholar] [CrossRef]
- Chen, Y.E.; Mao, H.T.; Wu, N.; Din, A.M.U.; Khan, A.; Zhang, H.Y.; Yuan, S. Salicylic Acid Protects Photosystem Ii by Alleviating Photoinhibition in Arabidopsis Thaliana under High Light. Int. J. Mol. Sci. 2020, 21, 1229. [Google Scholar] [CrossRef] [Green Version]
- Hogewoning, S.W.; Trouwborst, G.; Maljaars, H.; Poorter, H.; van Ieperen, W.; Harbinson, J. Blue Light Dose-Responses of Leaf Photosynthesis, Morphology, and Chemical Composition of Cucumis Sativus Grown under Different Combinations of Red and Blue Light. J. Exp. Bot. 2010, 61, 3107–3117. [Google Scholar] [CrossRef]
- Miao, Y.; Chen, Q.; Qu, M.; Gao, L.; Hou, L. Blue Light Alleviates ‘Red Light Syndrome’ by Regulating Chloroplast Ultrastructure, Photosynthetic Traits and Nutrient Accumulation in Cucumber Plants. Sci. Hortic. 2019, 257, 108680. [Google Scholar] [CrossRef]
- Wang, L.W.; Li, Y.; Xin, G.F.; Wei, M.; Mi, Q.H.; Yang, Q.C. Effects of Different Proportions of Red and Blue Light on the Growth and Photosynthesis of Tomato Seedlings. Chin. J. Appl. Ecol. 2017, 28, 1595–1602. [Google Scholar] [CrossRef]
- Moradi, S.; Kafi, M.; Aliniaeifard, S.; Salami, S.A.; Shokrpour, M.; Pedersen, C.; Moosavi-Nezhad, M.; Wróbel, J.; Kalaji, H.M. Blue Light Improves Photosynthetic Performance and Biomass Partitioning toward Harvestable Organs in Saffron (Crocus sativus L.). Cells 2021, 10, 1994. [Google Scholar] [CrossRef]
- Mathur, S.; Mehta, P.; Jajoo, A. Effects of Dual Stress (High Salt and High Temperature) on the Photochemical Efficiency of Wheat Leaves (Triticum Aestivum). Physiol. Mol. Biol. Plants 2013, 19, 179–188. [Google Scholar] [CrossRef] [Green Version]
- Bukhov, N.G.; Drozdova, I.S.; Bondar, V.V.; Mokronosov, A.T. Blue, Red and Blue plus Red Light Control of Chlorophyll Content and CO2 Gas Exchange in Barley Leaves: Quantitative Description of the Effects of Light Quality and Fluence Rate. Physiol. Plant. 1992, 85, 632–638. [Google Scholar] [CrossRef]
- Appenroth, K.J.; Keresztes, Á.; Sárvári, É.; Jaglarz, A.; Fischer, W. Multiple Effects of Chromate on Spirodela Polyrhiza: Electron Microscopy and Biochemical Investigations. Plant Biol. 2003, 5, 315–323. [Google Scholar] [CrossRef] [Green Version]
- Oukarroum, A.; Madidi, S.E.; Schansker, G.; Strasser, R.J. Probing the Responses of Barley Cultivars (Hordeum vulgare L.) by Chlorophyll a Fluorescence OLKJIP under Drought Stress and Re-Watering. Environ. Exp. Bot. 2007, 60, 438–446. [Google Scholar] [CrossRef]
- Aliniaeifard, S.; Seif, M.; Arab, M.; Mehrjerdi, M.Z.; Li, T.; Lastochkina, O. Growth and Photosynthetic Performance of Calendula Officinalis under Monochromatic Red Light. Int. J. Hortic. Sci. Technol. 2018, 5, 123–132. [Google Scholar] [CrossRef]
- Trouwborst, G.; Hogewoning, S.W.; van Kooten, O.; Harbinson, J.; van Ieperen, W. Plasticity of Photosynthesis after the “red Light Syndrome” in Cucumber. Environ. Exp. Bot. 2016, 121, 75–82. [Google Scholar] [CrossRef]
- Ouzounis, T.; Rosenqvist, E.; Ottosen, C.O. Spectral Effects of Artificial Light on Plant Physiology and Secondary Metabolism: A Review. HortScience 2015, 50, 1128–1135. [Google Scholar] [CrossRef] [Green Version]
- Zheng, L.; Van Labeke, M.C. Effects of Different Irradiation Levels of Light Quality on Chrysanthemum. Sci. Hortic. 2018, 233, 124–131. [Google Scholar] [CrossRef]
- Krapp, A.; Quick, W.P.; Stitt, M. Ribulose-1,5-Bisphosphate Carboxylase-Oxygenase, Other Calvin-Cycle Enzymes, and Chlorophyll Decrease When Glucose Is Supplied to Mature Spinach Leaves via the Transpiration Stream. Planta 1991, 186, 58–69. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Light Regime | Survival Ratio (%) | Scion Length (cm) | Node Number | Scion Stem Diameter (mm) | Leaf Area (cm2) | Root Fresh Weight (g) | Shoot Fresh Weight (g) | Plant Fresh Weight (g) | Root Dry Weight (g) | Shoot Dry Weight (g) | Plant Dry Weight (g) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| B | 100 | 14.66 c | 5.23 | 5.42 ab | 235.12 b | 1.39 | 3.94 | 5.33 b | 0.057 ab | 0.233 b | 0.290 b |
| W | 100 | 19.54 b | 5.20 | 5.40 ab | 217.94 b | 1.35 | 4.22 | 5.57 ab | 0.043 bc | 0.237 b | 0.280 b |
| RB | 100 | 19.70 b | 5.22 | 5.97 a | 294.57 a | 1.36 | 4.90 | 6.26 a | 0.063 a | 0.293 a | 0.357 a |
| R | 100 | 24.16 a | 5.21 | 4.90 b | 182.15 c | 0.97 | 3.95 | 4.92 b | 0.029 c | 0.174 c | 0.203 c |
| p | - | <0.0001 | 0.6765 | 0.0499 | 0.0002 | 0.1129 | 0.0710 | 0.0241 | 0.0055 | 0.0069 | 0.0049 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soltani, S.; Arouiee, H.; Salehi, R.; Nemati, S.H.; Moosavi-Nezhad, M.; Gruda, N.S.; Aliniaeifard, S. Morphological, Phytochemical, and Photosynthetic Performance of Grafted Tomato Seedlings in Response to Different LED Light Qualities under Protected Cultivation. Horticulturae 2023, 9, 471. https://doi.org/10.3390/horticulturae9040471
Soltani S, Arouiee H, Salehi R, Nemati SH, Moosavi-Nezhad M, Gruda NS, Aliniaeifard S. Morphological, Phytochemical, and Photosynthetic Performance of Grafted Tomato Seedlings in Response to Different LED Light Qualities under Protected Cultivation. Horticulturae. 2023; 9(4):471. https://doi.org/10.3390/horticulturae9040471
Chicago/Turabian StyleSoltani, Seyedreza, Hossein Arouiee, Reza Salehi, Seyed Hossein Nemati, Moein Moosavi-Nezhad, Nazim S. Gruda, and Sasan Aliniaeifard. 2023. "Morphological, Phytochemical, and Photosynthetic Performance of Grafted Tomato Seedlings in Response to Different LED Light Qualities under Protected Cultivation" Horticulturae 9, no. 4: 471. https://doi.org/10.3390/horticulturae9040471