
Functional Characterization of Ammonium Transporter MhAMT1;2 in Malus hupehensis
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Treatment
2.2. Determination of NO3−, NH4+ and Activities of Enzymes Involved in N Assimilation
2.3. Expression Analysis of AMTs
2.4. Generation of MhAMT1;2 cDNA
2.5. Bioinformatics Analysis of MhAMT1;2
2.6. Localization of MhAMT1;2 by GFP Fusion
2.7. Functional Complementation of MhAMT1;2 in NH4+ Uptake-Defective Yeast Mutant
2.8. Functional Complementation of MhAMT1;2 in NH4+ Uptake-Defective Arabidopsis Mutants
2.9. Statistical and Graphical Analyses
3. Results
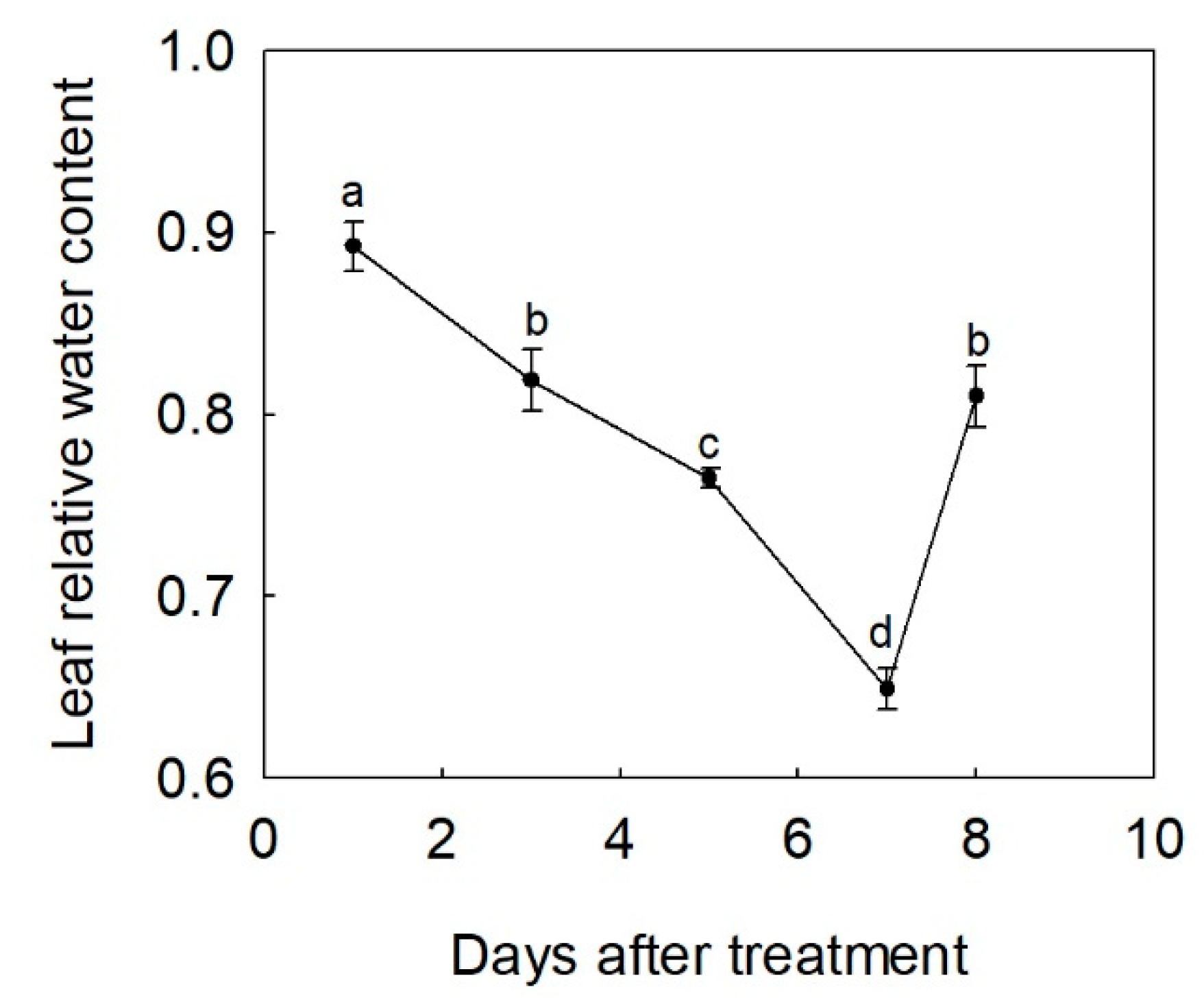
3.1. Changes of Leaf Relative Water Content under Drought and Rewatering
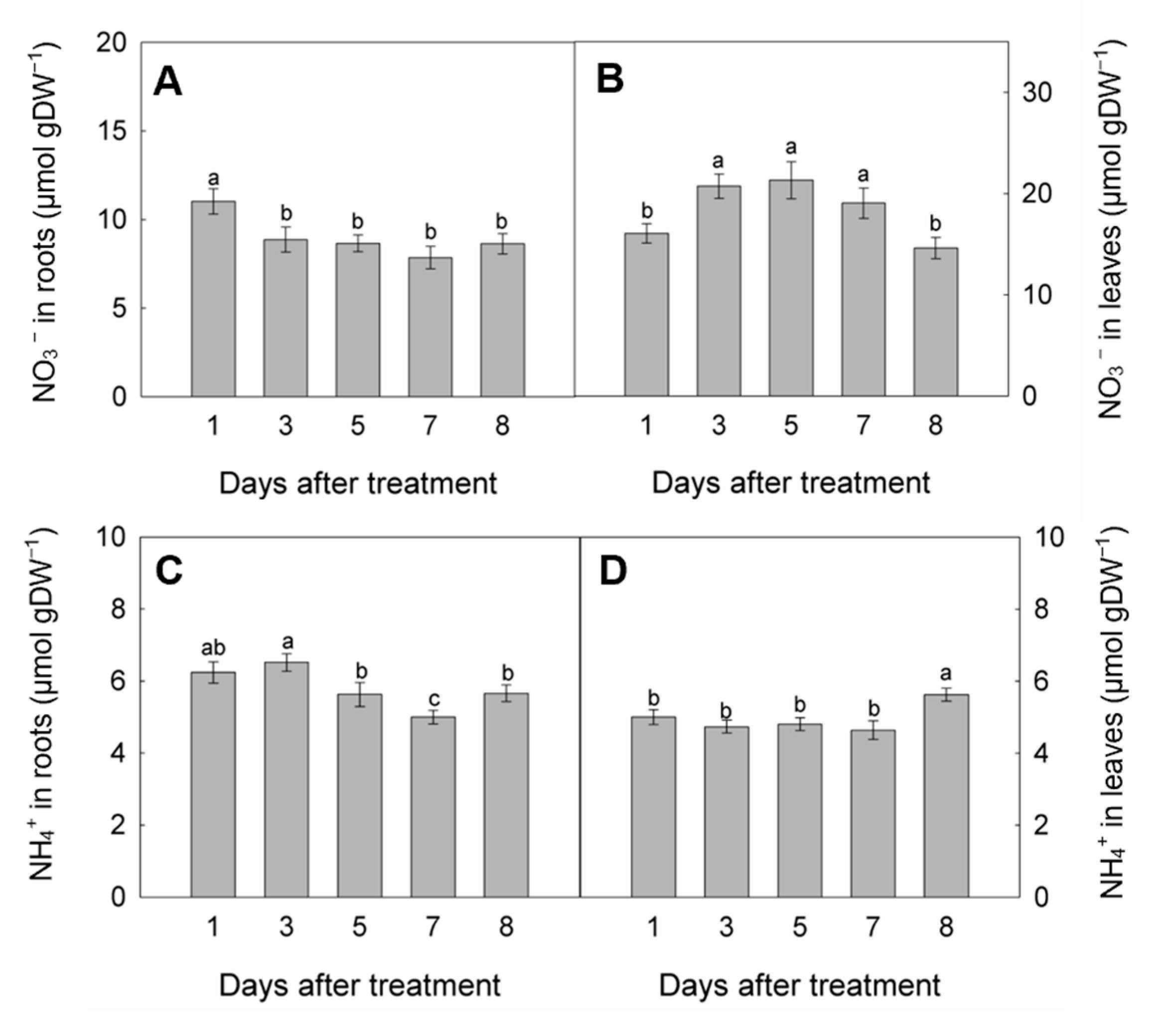
3.2. Concentrations of NH4+ and NO3− in Roots and Leaves
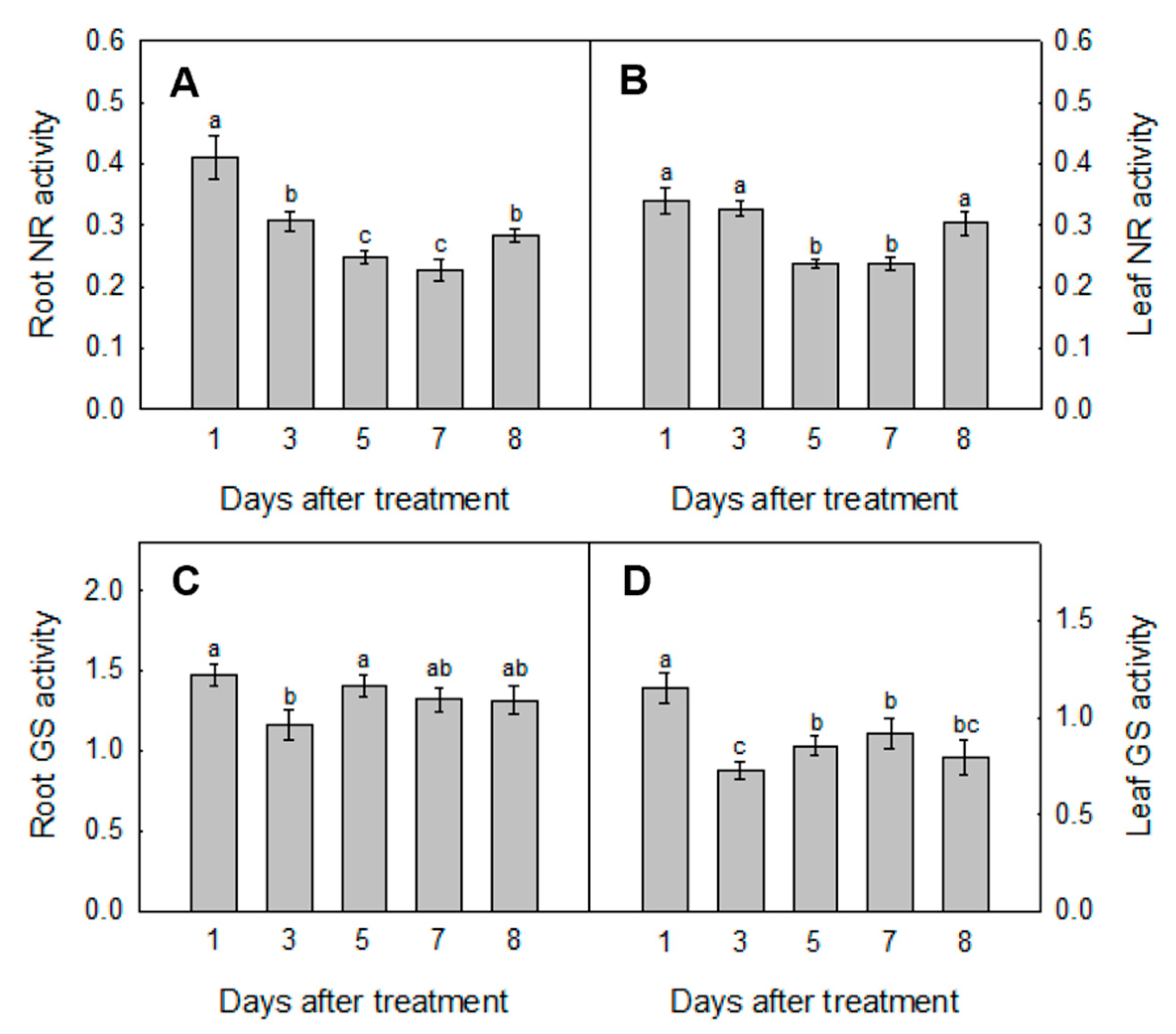
3.3. Activities Analysis of Key Enzymes Involved in N Assimilation
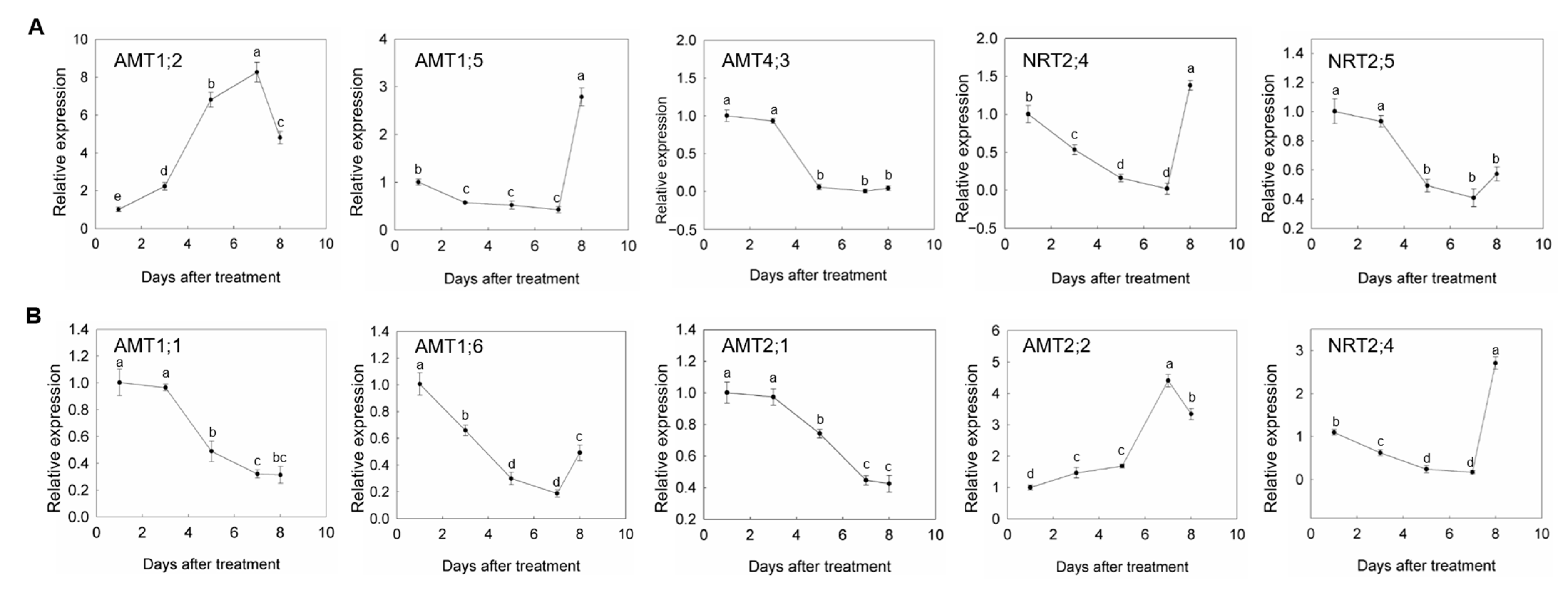
3.4. Expression Changes of AMTs and NRTs under Drought and Rewatering
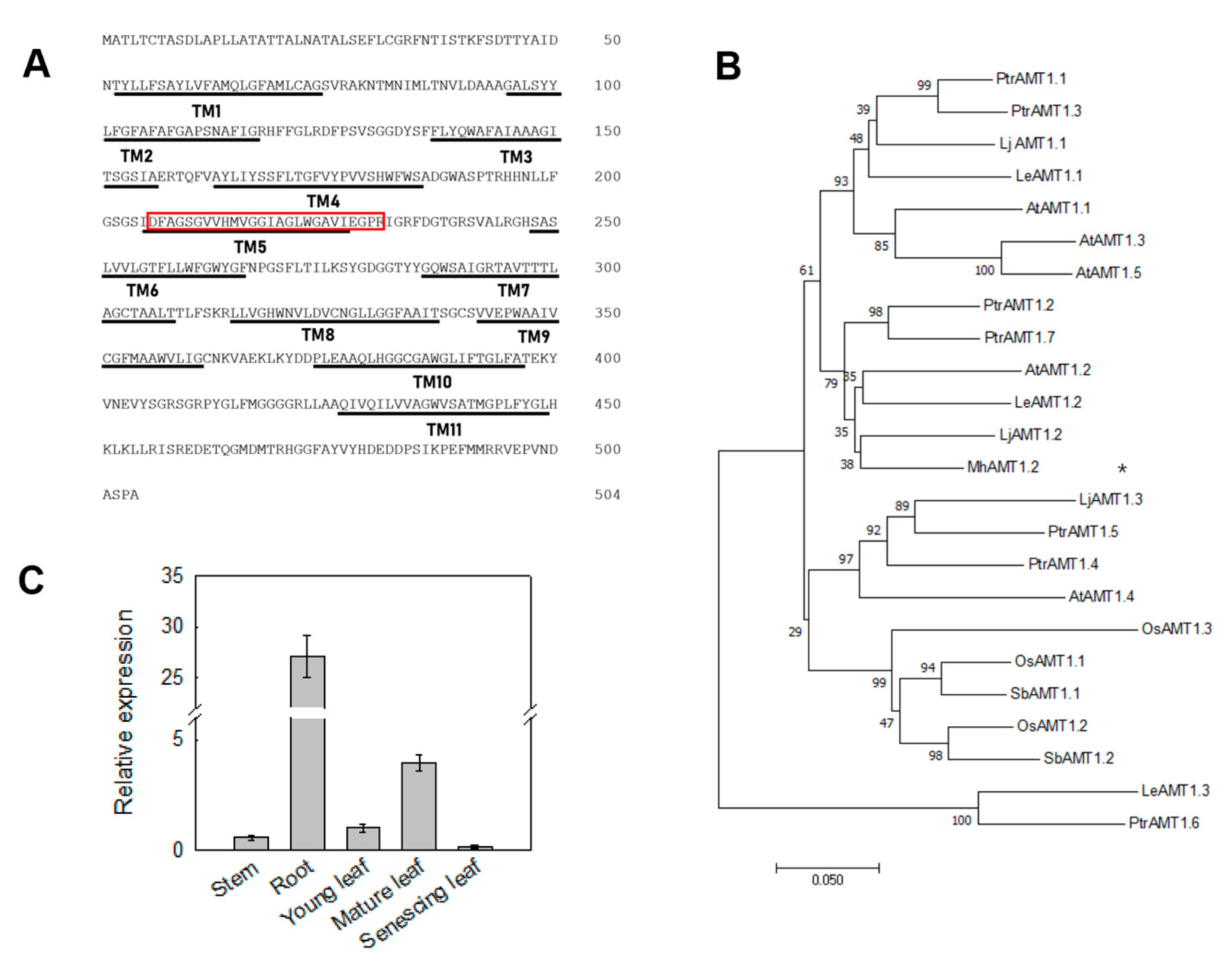
3.5. Isolation of MhAMT1;2 Gene from Malus hupehensis
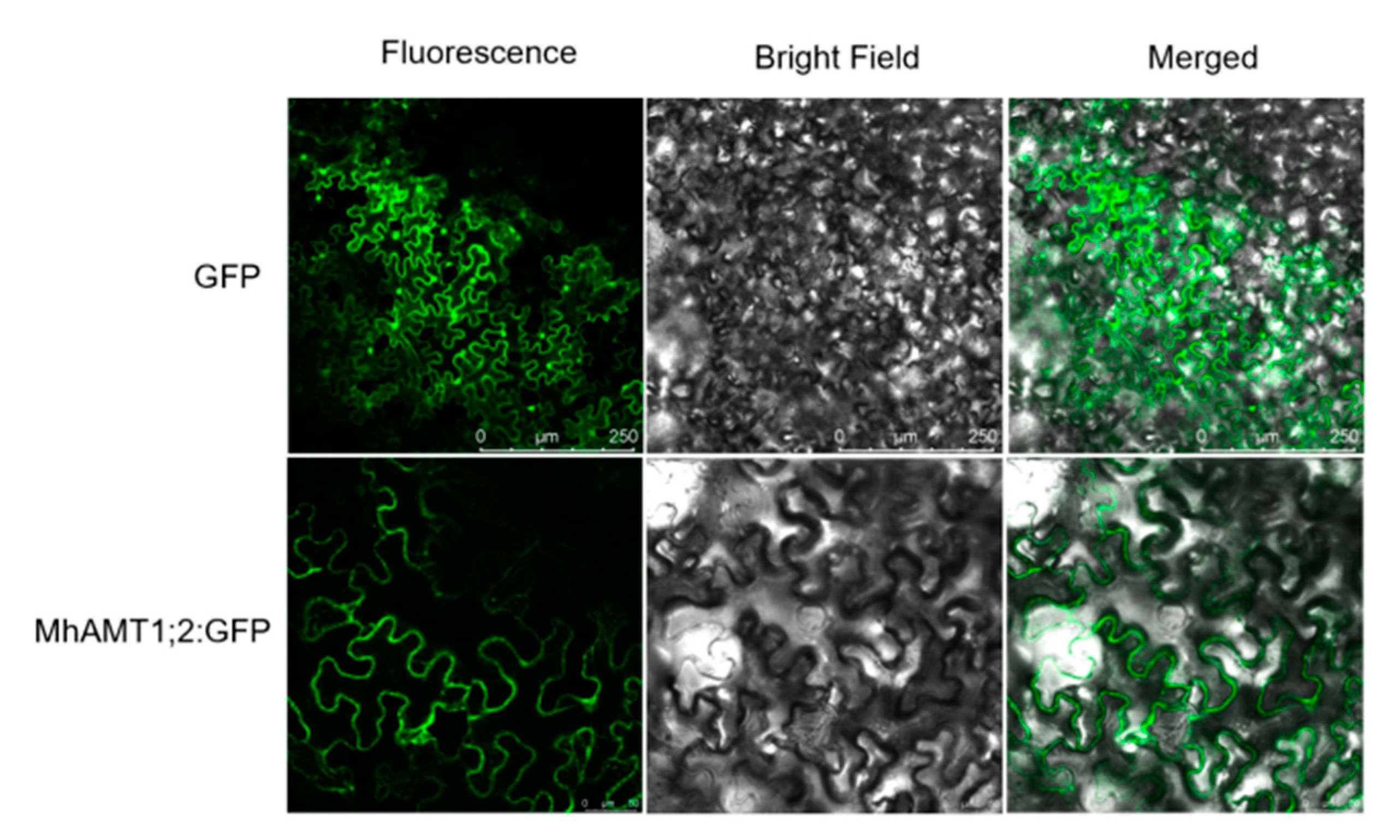
3.6. Localization of MhAMT1;2 in Tobacco Epidermal Cells
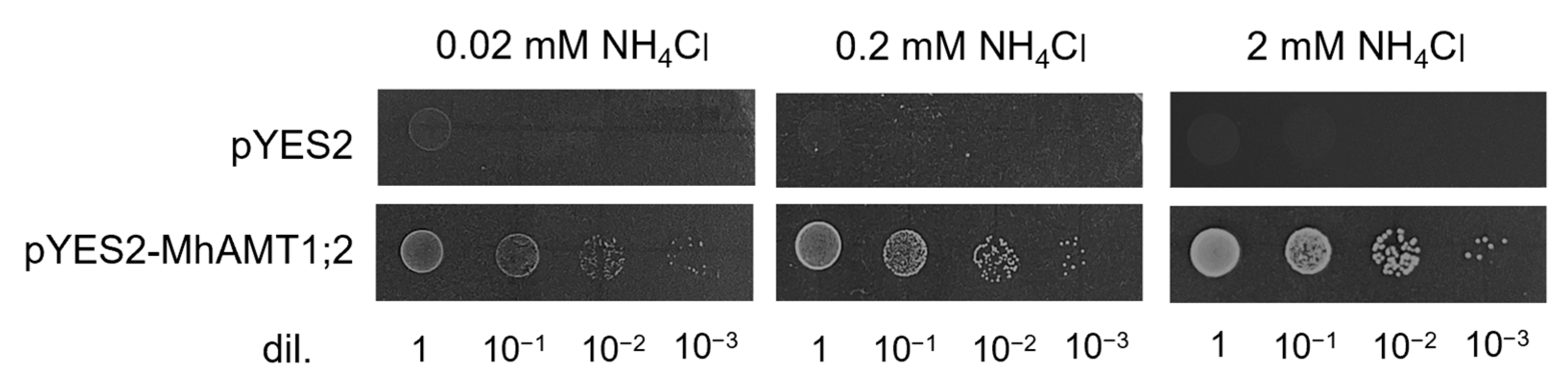
3.7. Transport Capacity of MhAMT1;2
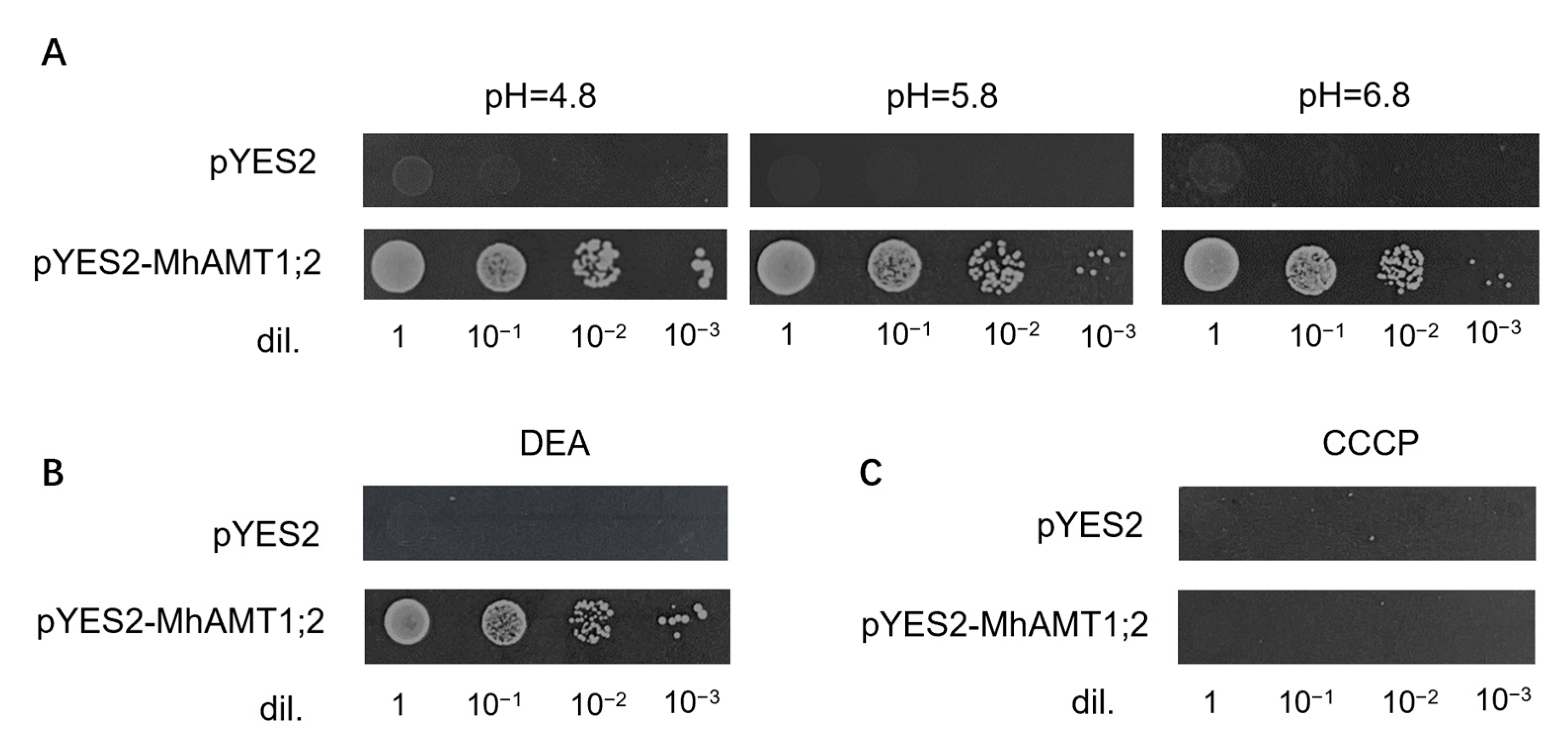
3.8. Proton and ATP Dependence of MhAMT1;2-Mediated Transport Process
3.9. Functional Complementation of MhAMT1;2 in NH4+ Uptake-Defective Arabidopsis Mutants
4. Discussion
4.1. Changes of N Uptake and Assimilation under Drought and Rewatering
4.2. MhAMT1;2 Is a ATP-Dependent High Affinity NH4+ Transporter
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, Y.F.; Wang, Y.; Wu, W.H. Membrane Transporters for Nitrogen, Phosphate and Potassium Uptake in Plants. J. Integr. Plant Biol. 2008, 50, 835–848. [Google Scholar] [CrossRef] [PubMed]
- Miller, A.J.; Cramer, M.D. Root nitrogen acquisition and assimilation. Plant Soil 2004, 274, 1–36. [Google Scholar] [CrossRef]
- Buljovcic, Z.; Engels, C. Nitrate uptake ability by maize roots during and after drought stress. Plant Soil 2001, 229, 125–135. [Google Scholar] [CrossRef]
- He, M.Z.; Dijkstra, F.A. Drought effect on plant nitrogen and phosphorus: A metaanalysis. New Phytol. 2014, 4, 924–931. [Google Scholar] [CrossRef]
- Du, Y.; Zhao, Q.; Chen, L.; Yao, X.; Xie, F. Effect of drought stress at reproductive stages on growth and nitrogen metabolism in soybean. Agronomy 2020, 10, 302. [Google Scholar] [CrossRef] [Green Version]
- Xia, H.; Xu, T.; Zhang, J.; Shen, K.; Li, Z.; Liu, J. Drought-Induced Responses of Nitrogen Metabolism in Ipomoea batatas. Plants 2020, 9, 1341. [Google Scholar] [CrossRef] [PubMed]
- Meng, S.; Zhang, C.X.; Su, L.; Li, Y.M.; Zhao, Z. Nitrogen uptake and metabolism of Populus simonii in response to PEG-induced drought stress. Environ. Exp. Bot. 2016, 123, 78–87. [Google Scholar] [CrossRef]
- Huang, L.L.; Li, M.J.; Zhou, K.; Sun, T.T.; Hu, L.Y.; Li, C.Y.; Ma, F.W. Uptake and metabolism of ammonium and nitrate in response to drought stress in Malus prunifolia. Plant Physiol. Biochem. 2018, 127, 185–193. [Google Scholar] [CrossRef]
- Tian, J.; Pang, Y.; Zhao, Z. Drought, Salinity, and Low Nitrogen Differentially Affect the Growth and Nitrogen Metabolism of Sophora japonica (L.) in a Semi-Hydroponic Phenotyping Platform. Front. Plant Sci. 2021, 12, 715456. [Google Scholar] [CrossRef]
- Lee, S.; Marmagne, A.; Park, J.; Fabien, C.; Yim, Y.; Kim, S.-J.; Kim, T.-H.; Lim, P.O.; Masclaux-Daubresse, C.; Nam, H.G. Concurrent activation of OsAMT1;2 and OsGOGAT1 in rice leads to enhanced nitrogen use efficiency under nitrogen limitation. Plant J. 2020, 103, 7–20. [Google Scholar] [CrossRef]
- Ding, L.; Gao, C.; Li, Y.; Zhu, Y.; Xu, G.; Guo, S.W. The enhanced drought tolerance of rice plants under ammonium is related to aquaporin (AQP). Plant Sci. 2015, 234, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Crespo, E.; Gómez-Pastor, R.; Scalschi, L.; Llorens, E.; Camañes, G.; García-Agustín, P. NH4+ induces antioxidant cellular machinery and provides resistance to salt stress in citrus plants. Trees 2014, 28, 1693–1704. [Google Scholar] [CrossRef]
- Yuan, L.X.; Loqué, D.; Kojima, S.; Rauch, S.; Ishiyama, K.; Takahashi, H.; Von Wirén, N. The organization of high-affinity ammonium uptake in Arabidopsis roots depends on the spatial arrangement and biochemical properties of AMT1-type transporters. Plant Cell 2007, 19, 2636–2652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lauter, F.R.; Ninnemann, O.; Bucher, M.; Riesmeier, J.W.; Frommer, W.B. Preferential expression of an ammonium transporter and of two putative nitrate transporters in root hairs of tomato. Proc. Natl. Acad. Sci. USA 1996, 93, 8139–8144. [Google Scholar] [CrossRef] [Green Version]
- Salvemini, F.; Marini, A.M.; Riccio, A.; Patriarca, E.J.; Chiurazzi, M. Functional characterization of an ammonium transporter gene from Lotus japonicus. Gene 2001, 270, 237–243. [Google Scholar] [CrossRef]
- Sonoda, Y.; Ikeda, A.; Saiki, S.; Wirén, N.V.; Yamaya, T.; Yamaguchi, J. Distinct Expression and Function of Three Ammonium Transporter Genes (OsAMT1;1–1;3) in Rice. Plant Cell Physiol. 2003, 44, 726–734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konishi, N.; Ma, J. Three polarly localized ammonium transporter 1 members are cooperatively responsible for ammonium uptake in rice under low ammonium condition. New Phytol. 2021, 232, 1778–1792. [Google Scholar] [CrossRef]
- Kosala, R.; Ashraf, E.K.; Satinder, G.; Bi, Y.M.; Rothstein, S.J. AMT1;1 transgenic rice plants with enhanced NH4 permeability show superior growth and higher yield under optimal and suboptimal NH4 conditions. J. Exp. Bot. 2014, 4, 965–979. [Google Scholar] [CrossRef] [Green Version]
- Li, W.X.; Feng, Z.M.; Zhang, C.X. Ammonium transporter PsAMT1.2 from Populus simonii functions in nitrogen uptake and salt resistance. Tree Physiol. 2021, 41, 2392–2408. [Google Scholar] [CrossRef]
- Bu, Y.; Takano, T.; Liu, S. The role of ammonium transporter (AMT) against salt stress in plants. Plant Signal. Behav. 2019, 14, 8. [Google Scholar] [CrossRef]
- Duan, J.F.; Tian, H.; Gao, Y.J. Expression of nitrogen transporter genes in roots of winter wheat (Triticum aestivum L.) in response to soil drought with contrasting nitrogen supplies. Crop Pasture Sci. 2016, 67, 128–136. [Google Scholar] [CrossRef]
- Huang, L.L.; Li, J.Z.; Zhang, B.; Hao, Y.Y.; Ma, F.W. Genome-Wide Identification and Expression Analysis of AMT Gene Family in Apple (Malus domestica Borkh.). Horticulturae 2022, 8, 457. [Google Scholar] [CrossRef]
- Huang, L.L.; Li, M.J.; Shao, Y.; Sun, T.T.; Li, C.Y.; Ma, F.W. Ammonium uptake increases in response to PEG-induced drought stress in Malus hupehensis Rehd. Environ. Exp. Bot. 2018, 151, 32–42. [Google Scholar] [CrossRef]
- Brautigam, A.; Gagneul, D.; Weber, A.P. High-throughput colorimetric method for the parallel assay of glyoxylic acid and ammonium in a single extract. Anal. Biochem. 2007, 362, 151–153. [Google Scholar] [CrossRef]
- Natali, S.M.; Sanudo-Wilhelmy, S.A.; Lerdau, M.T. Effects of elevated carbon dioxide and nitrogen fertilization on nitrate reductase activity in sweetgum and loblolly pine trees in two temperate forests. Plant Soil 2009, 314, 197–210. [Google Scholar] [CrossRef]
- Yu, X.Z.; Zhang, F.Z. Activities of nitrate reductase and glutamine synthetase in rice seedlings during cyanide metabolism. J. Hazard. Mater. 2012, 225–226, 190–194. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Rennenberg, H.; Loreto, F.; Polle, A.; Brilli, F.; Fares, S.; Beniwal, R.S.; Gessler, A. Physiological responses of forest trees to heat and drought. Plant Biol. 2006, 8, 556–571. [Google Scholar] [CrossRef]
- Bai, J.H.; Jia, J.; Huang, C.; Wang, Q.G.; Wang, W.; Zhang, G.L.; Cui, B.S.; Liu, X.H. Selective uptake of nitrogen by Suaeda salsa under drought and salt stresses and nitrogen fertilization using 15N. Ecol. Eng. 2017, 102, 542–545. [Google Scholar] [CrossRef]
- Fotelli, M.N.; Rennenberg, H.; Geβler, A. Effects of drought on the competitive interference of an early successional species (Rubus fruticosus) on fagus sylvatica l. seedlings: 15N uptake and partitioning, responses of amino acids and other n compounds. Plant Biol. 2002, 4, 311–320. [Google Scholar] [CrossRef]
- Andrews, M.; Lea, P.J.; Raven, J.A.; Azevedo, R.A. Nitrogen use efficiency. 3. nitrogen fixation: Genes and costs. Ann. Appl. Biol. 2009, 155, 1–13. [Google Scholar] [CrossRef]
- Shi, W.M.; Xu, W.F.; Li, S.M.; Zhao, X.Q.; Dong, G.Q. Responses of two rice cultivars differing in seedling-stage nitrogen use efficiency to growth under low-nitrogen conditions. Plant Soil 2010, 326, 291–302. [Google Scholar] [CrossRef]
- Zhang, C.; Meng, S.; Li, M.; Zhong, Z. Transcriptomic insight into nitrogen uptake and metabolism of Populus simonii in response to drought and low nitrogen stresses. Tree Physiol. 2018, 38, 1672–1684. [Google Scholar] [CrossRef]
- Glass, A.; Britto, D.T.; Kaiser, B.N.; Kinghorn, J.R.; Kronzucker, H.J.; Anshuman, K.; Mamoru, O.; Suman, R.; Siddiqi, M.Y.; Unkles, S.E. The regulation of nitrate and ammonium transport systems in plants. J. Exp. Bot. 2002, 53, 855–864. [Google Scholar] [CrossRef] [Green Version]
- Ninnemann, O.; Jauniaux, J.C.; Frommer, W.B. Identification of a high affinity NH4+ transporter from plants. EMBO J. 1994, 13, 3464–3471. [Google Scholar] [CrossRef] [PubMed]
- Loque, D.; Wiren, N.V. Regulatory levels for the transport of ammonium in plant roots. J. Exp. Bot. 2004, 55, 1293–1305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Couturier, J.; Montanini, B.; Martin, F.; Brun, A.; Blaudez, D.; Chalot, M. The expanded family of ammonium transporters in the perennial poplar plant. New Phytol. 2007, 174, 137–150. [Google Scholar] [CrossRef]
- Li, H.; Yang, Q.S.; Liu, W.; Lin, J.; Chang, Y.H. The AMT1 family genes from Malus robusta display differential transcription features and ammonium transport abilities. Mol. Biol. Rep. 2017, 44, 379–390. [Google Scholar] [CrossRef]
- Bu, Y.Y.; Sun, B.; Zhou, A.M.; Zhang, X.X.; Lee, I.; Liu, S.K. Identification and characterization of a PutAMT1;1 gene from puccinellia tenuiflora. PLoS ONE 2013, 8, e83111. [Google Scholar] [CrossRef]
- Gu, R.; Duan, F.; An, X.; Zhang, F.; Von Wirén, N.; Yuan, L. Characterization of AMT-mediated high-affinity ammonium uptake in roots of maize (Zea mays L.). Plant Cell Physiol. 2013, 54, 1515–1524. [Google Scholar] [CrossRef]
- Ludewig, U.; Von Wirén, N.; Frommer, W.B. Uniport of NH4+ by the root hair plasma membrane ammonium transporter LeAMT1;1. J. Biol. Chem. 2002, 277, 13548–13555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, S.Y.; Hao, D.L.; Cong, Y.; Jin, M.; Su, Y.H. The rice OsAMT1;1 is a proton-independent feedback regulated ammonium transporter. Plant Cell Rep. 2015, 34, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Ortiz-Ramirez, C.; Mora, S.I.; Trejo, J.; Pantoja, O. PvAMT1;1, a highly selective ammonium transporter that functions as H+/NH4+ symporter. J. Biol. Chem. 2011, 286, 31113–31122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Von Wirén, N.; Lauter, F.R.; Ninnemann, O.; Gillissen, B.; Walch-Liu, P.; Engels, C.; Jost, W.H.; Frommer, W.B. Differential regulation of three functional ammonium transporter genes by nitrogen in root hairs and by light in leaves of tomato. Plant J. Cell Mol. Biol. 2000, 21, 167–175. [Google Scholar] [CrossRef]
- D’Apuzzo, E.; Rogato, A.; Simon-Rosin, U.; Alaoui, H.E.; Barbulova, A.; Betti, M.; Dimou, M.; Katinakis, P.; Márquez, A.J.; Marini, A.; et al. Characterization of three functional high-affinity ammonium transporters in Lotus japonicus with differential transcriptional regulation and spatial expression. Plant Physiol. 2004, 134, 1763–1774. [Google Scholar] [CrossRef] [Green Version]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Yu, M.; Li, H.; Yang, G.; Huang, L.; Hao, Y. Functional Characterization of Ammonium Transporter MhAMT1;2 in Malus hupehensis. Horticulturae 2023, 9, 434. https://doi.org/10.3390/horticulturae9040434
Li J, Yu M, Li H, Yang G, Huang L, Hao Y. Functional Characterization of Ammonium Transporter MhAMT1;2 in Malus hupehensis. Horticulturae. 2023; 9(4):434. https://doi.org/10.3390/horticulturae9040434
Chicago/Turabian StyleLi, Jiazhen, Muting Yu, Huibin Li, Guangkai Yang, Linlin Huang, and Yanyan Hao. 2023. "Functional Characterization of Ammonium Transporter MhAMT1;2 in Malus hupehensis" Horticulturae 9, no. 4: 434. https://doi.org/10.3390/horticulturae9040434