
Use of Bacteria Isolated from Berry Rhizospheres as Biocontrol Agents for Charcoal Rot and Root-Knot Nematode Strawberry Diseases


Abstract
:1. Introduction
2. Materials and Methods
2.1. In Vitro Experiments for Biocontrol of M. phaseolina
2.1.1. Bacterial Strain Isolation
2.1.2. Bacterial Strain Identification
2.1.3. In Vitro Assays for Plant Growth-Promoting Activities
2.1.4. In Vitro Assays for Biocontrol Enzymatic Activities
2.1.5. In Vitro Assays for Biocontrol Activity against M. phaseolina
2.2. In Planta Experiments for Biocontrol
2.2.1. Experimental Design
2.2.2. Plant Material
2.2.3. Bacterial Inoculation
2.2.4. Fungal Inoculation
2.2.5. Nematode Inoculation
2.2.6. Plant Growing Conditions
2.2.7. Parameters Evaluation for Plant Growth and Disease
2.3. Statistics
3. Results
3.1. Bacterial Identification
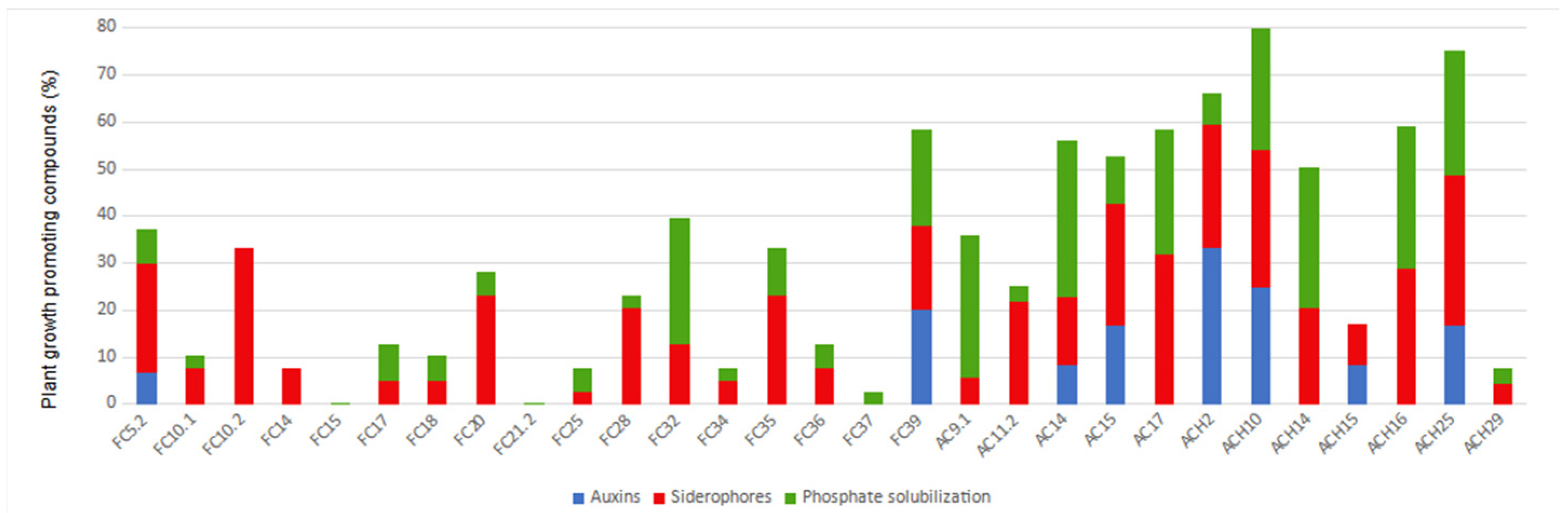
3.2. Plant Growth-Promoting Activities
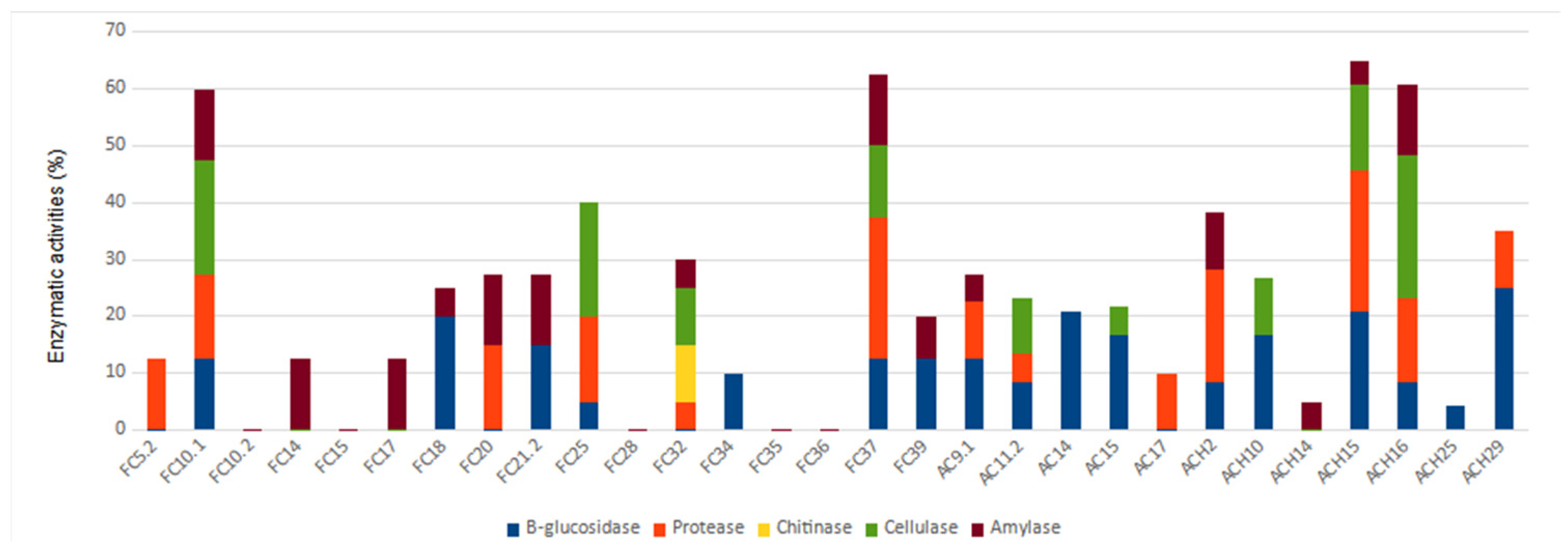
3.3. Biocontrol Enzymatic Activities
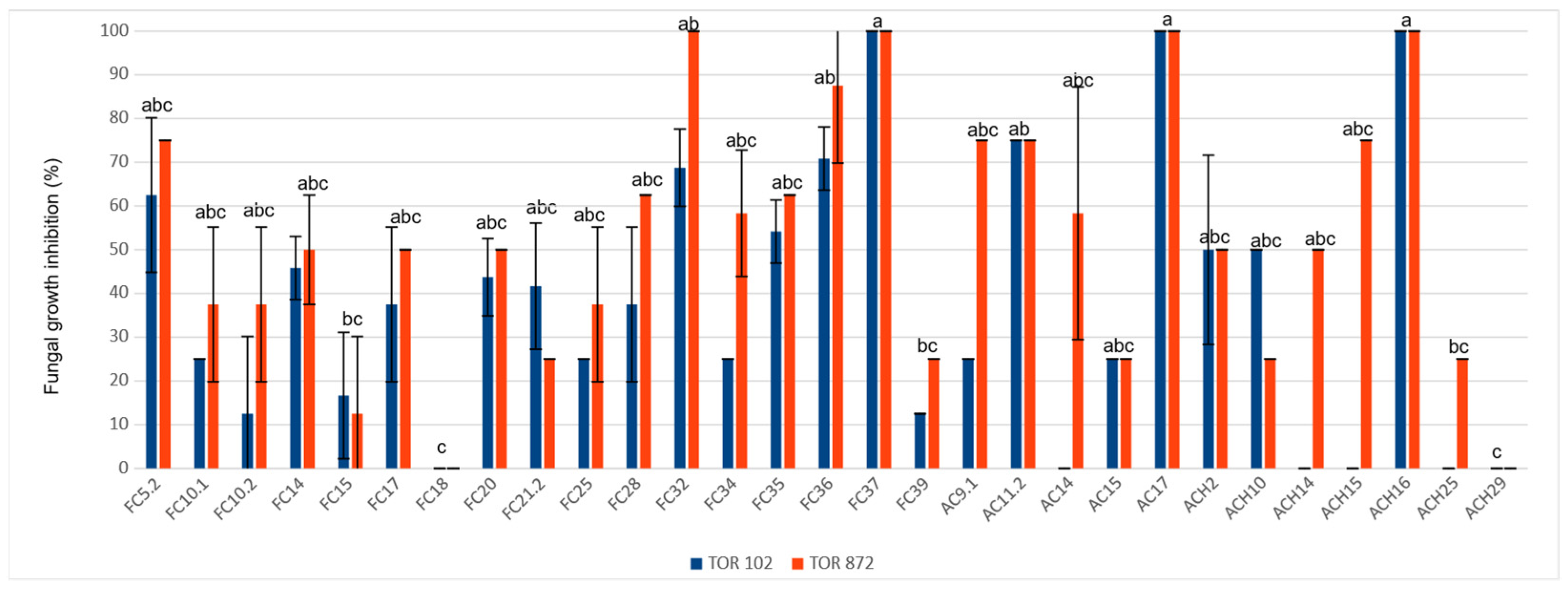
3.4. In Vitro Antifungal Activities against M. phaseolina
3.5. Strawberry Biomass
3.6. M. phaseolina Disease Symptoms
3.7. M. hapla Disease Symptoms
4. Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- MAPAMA. Anuario de Estadística. Avance 2021; Ministerio de Agricultura, Pesca y Alimentación: Madrid, Spain, 2022. [Google Scholar]
- Arroyo, F.T.; Llergo, Y.; Aguado, A.; Romero, F. First report of Fusarium wilt caused by Fusarium oxysporum on strawberry in Spain. Plant Dis. 2009, 93, 323. [Google Scholar] [CrossRef] [PubMed]
- Borrero, C.; Capote, N.; Gallardo, M.A.; Avilés, M. First report of vascular wilt caused by Fusarium proliferatum on strawberry in Spain. Plant Dis. 2019, 103, 581. [Google Scholar] [CrossRef]
- Avilés, M.; Castillo, S.; Bascón, J.; Zea-Bonilla, T.; Martín-Sánchez, P.M.; Pérez-Jiménez, R.M. First report of Macrophomina phaseolina causing crown and root rot of strawberry in Spain. Plant Pathol. 2008, 57, 382. [Google Scholar] [CrossRef]
- Pastrana, A.M.; Capote, N.; de los Santos, B.; Romero, F.; Basallote-Ureba, M.J. First report of Fusarium solani causing crown and root rot on strawberry crops in Southwestern Spain. Plant Dis. 2014, 98, 161. [Google Scholar] [CrossRef]
- Talavera, M.; Miranda, L.; Gómez-Mora, J.A.; Vela, M.D.; Verdejo-Lucas, S. Nematode management in the strawberry fields of Southern Spain. Agronomy 2019, 9, 252. [Google Scholar] [CrossRef] [Green Version]
- Talavera, M.; Gómez-Mora, J.A.; de los Santos, B.; Miranda, L.; Vela, M.D.; Fernández-Plaza, M.; Medina-Mínguez, J.J.; Soria, C.; Verdejo-Lucas, S. Problemática Fitopatológica del cultivo de la fresa en Huelva. Agricultura 2018, 1022, 6–9. [Google Scholar]
- López-Aranda, J.M.; Domínguez, P.; Miranda, L.; de Los Santos, B.; Talavera, M.; Daugovish, O.; Soria, C.; Chamorro, M.; Medina, J.J. Fumigant use for strawberry production in Europe: The current landscape and solutions. Int. J. Fruit Sci. 2016, 16, 1–15. [Google Scholar] [CrossRef]
- Greco, N.; López-Aranda, J.M.; Saporiti, M.; Maccarini, C.; de Tommaso, N.; Myrta, A. Sustainability of European vegetable and strawberry production in relation to fumigation practices in the EU. Acta Hortic. 2020, 1270, 203–210. [Google Scholar] [CrossRef]
- Dimkić, I.; Janakiev, T.; Petrović, M.; Degrassi, G.; Fira, D. Plant-associated Bacillus and Pseudomonas antimicrobial activities in plant disease suppression via biological control mechanisms—A review. Physiol. Mol. Plant Pathol. 2022, 117, 101754. [Google Scholar] [CrossRef]
- Zhai, Y.; Shao, Z.; Cai, M.; Zheng, L.; Li, G.; Huang, D.; Cheng, W.; Thomashow, L.S.; Weller, D.M.; Yu, Z.; et al. Multiple modes of nematode control by volatiles of Pseudomonas putida 1A00316 from Antarctic soil against Meloidogyne incognita. Front. Microbiol. 2018, 9, 253. [Google Scholar] [CrossRef] [Green Version]
- Lalloo, R.; Maharajh, D.; Görgens, J.; Gardiner, N. A downstream process for production of a viable and stable Bacillus cereus aquaculture biological agent. Appl. Microbiol. Biotechnol. 2010, 86, 499–508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, X.-L.; Zhao, X.-M.; Zhao, S.-Y.; Zhao, J.-L.; Mao, Z.-C. The biocontrol functions of Bacillus velezensis strain Bv-25 against Meloidogyne incognita. Front. Microbiol. 2022, 13, 843041. [Google Scholar] [CrossRef] [PubMed]
- Dehghanian, S.Z.; Abdollahi, M.; Charehgani, H.; Niazi, A. Combined of salicylic acid and Pseudomonas fluorescens CHA0 on the expression of PR1 gene and control of Meloidogyne javanica in tomato. Biol. Control 2020, 141, 104134. [Google Scholar] [CrossRef]
- Viejobueno, J.; Rodríguez-Berbel, N.; Miranda, L.; de los Santos, B.; Camacho, M. Potential bacterial antagonists for the control of charcoal rot (Macrophomina phaseolina) in strawberry. Horticulturae 2021, 7, 457. [Google Scholar] [CrossRef]
- De la Lastra, E.; Camacho, M.; Capote, N. Soil bacteria as potential biological control agents of Fusarium species associated with asparagus decline syndrome. Appl. Sci. 2021, 11, 8356. [Google Scholar] [CrossRef]
- Lane, D.J. 16S/23S RRNA sequencing. In Nucleic Acid Techniques in Bacterial Systematic; Stackebrandt, E., Goodfellow, M., Eds.; John Wiley and Sons: New York, NY, USA, 1991; pp. 115–175. [Google Scholar]
- Yoon, S.-H.; Ha, S.-M.; Kwon, S.; Lim, J.; Kim, Y.; Seo, H.; Chun, J. Introducing EzBioCloud: A taxonomically united database of 16s RRNA gene sequences and whole-genome assemblies. Int. J. Syst. Evol. Microbiol. 2017, 67, 1613–1617. [Google Scholar] [CrossRef] [PubMed]
- Benizri, E.; Courtade, A.; Picard, C.; Guckert, A. Role of maize root exudates in the production of auxins by Pseudomonas fluorescens M.3.1. Soil. Biol. Biochem. 1998, 30, 1481–1484. [Google Scholar] [CrossRef]
- Alexander, D.B.; Zuberer, D.A. Use of chrome azurol S reagents to evaluate siderophore production by rhizosphere bacteria. Biol. Fertil. Soils 1991, 12, 39–45. [Google Scholar] [CrossRef]
- De Freitas, J.R.; Banerjee, M.R.; Germida, J.J. Phosphate-solubilizing rhizobacteria enhance the growth and yield but not phosphorus uptake of canola (Brassica napus L.). Biol. Fertil. Soils 1997, 24, 358–364. [Google Scholar] [CrossRef]
- Viejobueno, J.; Albornoz, P.L.; Camacho, M.; de los Santos, B.; Martínez-Zamora, M.G.; Salazar, S.M. Protection of strawberry plants against charcoal rot disease (Macrophomina phaseolina) induced by Azospirillum brasilense. Agronomy 2021, 11, 195. [Google Scholar] [CrossRef]
- Reznikov, S.; Vellicce, G.R.; González, V.; de Lisi, V.; Castagnaro, A.P.; Ploper, L.D. Evaluation of chemical and biological seed treatments to control charcoal rot of soybean. J. General. Plant Pathol. 2016, 82, 273–280. [Google Scholar] [CrossRef] [Green Version]
- Avilés, M.; Castillo, S.; Borrero, C.; Castillo, M.L.; Zea-Bonilla, T.; Pérez-Jiménez, R.M. Response of strawberry cultivars: “Camarosa”, “Candonga” and “Ventana” to inoculation with isolates of Macrophomina phaseolina. Acta Hortic. 2009, 842, 291–294. [Google Scholar] [CrossRef]
- Hussey, R.S.; Barker, K.R. A comparison of methods of collecting inocula of Meloidogyne spp., including a new technique. Plant Dis. Rep. 1973, 57, 1025–1028. [Google Scholar]
- Cesarz, S.; Eva Schulz, A.; Beugnon, R.; Eisenhauer, N. Testing soil nematode extraction efficiency using different variations of the Baermann-Funnel method. Soil Org. 2019, 91, 61–72. [Google Scholar] [CrossRef] [PubMed]
- Koike, S.T.; Arias, R.S.; Hogan, C.S.; Martin, F.N.; Gordon, T.R. Status of Macrophomina phaseolina on strawberry in California and preliminary characterization of the pathogen. Int. J. Fruit Sci. 2016, 16, 148–159. [Google Scholar] [CrossRef]
- Fang, X.L.; Phillips, D.; Li, H.; Sivasithamparam, K.; Barbetti, M.J. Severity of crown and root diseases of strawberry and associated fungal and oomycete pathogens in Western Australia. Australas. Plant Pathol. 2011, 40, 109–119. [Google Scholar] [CrossRef]
- Bridge, J.; Page, S.L.J. Estimation of root knot nematode infestation level on roots using a rating chart. Trop. Pest Manag. 1980, 26, 296–298. [Google Scholar] [CrossRef]
- Coolen, W.A. Methods for the extraction of Meloidogyne spp. and other nematodes from roots and soil. In Root-knot nematodes (Meloidogyne Species)—Systematics, Biology and Control; Lamberti, F., Taylor, C.E., Eds.; Academic Press: London, UK, 1979; pp. 317–329. [Google Scholar]
- Villarreal-Delgado, M.F.; Villa-Rodríguez, E.D.; Cira-Chávez, L.A.; Estrada-Alvarado, M.I.; Parra-Cota, F.I.; de los Santos-Villalobos, S. The genus Bacillus as a biological control agent and its implications in the agricultural biosecurity. Rev. Mex. Fitopatol. 2017, 36, 95–130. [Google Scholar] [CrossRef]
- Anayo, O.F.; Scholastica, E.C.; Peter, O.C.; Nneji, U.G.; Obinna, A.; Mistura, L.O. The beneficial roles of Pseudomonas in Medicine, Industries, and Environment: A Review. In Pseudomonas Aeruginosa—An Armory within; Sriramulu, D., Ed.; IntechOpen: London, UK, 2019. [Google Scholar] [CrossRef] [Green Version]
- Ghazi, A.; Attia, E.A.; Rashed, N.M. Management of charcoal rot (Macrophomina phaseolina) infection in geranium (Pelargonium graveolens L.) using biocontrol agents and essential oils. Environ. Biodivers. Soil Secur. 2018, 2, 131–142. [Google Scholar] [CrossRef]
- Pastrana, A.M.; Basallote-Ureba, M.J.; Aguado, A.; Akdi, K.; Capote, N. Biological control of strawberry soil-borne pathogens Macrophomina phaseolina and Fusarium solani, using Trichoderma asperellum and Bacillus spp. Phytopathol. Mediterr. 2016, 55, 109–120. [Google Scholar] [CrossRef]
- Abd-El-Kareem, F.; Elshahawy, I.E.; Abd-Elgawad, M.M.M. Native bacteria for field biocontrol of black root rot in Egyptian strawberry. Bull. Natl. Res. Cent. 2022, 46, 82. [Google Scholar] [CrossRef]
- Lovaisa, N.C.; Guerrero, M.F.; Delaporte, P.G.A.; Salazar, S.M. Response of strawberry plants inoculated with Azospirillum and Burkholderia at field conditions. Rev. Agron. Noroeste Argent 2015, 35, 33–36. [Google Scholar]
- Guerrero-Molina, M.F.; Lovaisa, N.C.; Salazar, S.M.; Martínez-Zamora, M.G.; Díaz-Ricci, J.C.; Pedraza, R.O. Physiological, structural and molecular traits activated in strawberry plants after inoculation with the plant growth-promoting bacterium Azospirillum brasilense REC3. Plant Biol. 2014, 17, 766–773. [Google Scholar] [CrossRef] [PubMed]
- Askary, T.H.; Pala-Martinelli, P.R.; Askary, T.H. Biocontrol Agents of Phytonematodes; CAB International: Wallingford, UK; Oxford, UK, 2015; ISBN 9788578110796. [Google Scholar]
- Flor-Peregrín, E.; Azcón, R.; Salmerón, T.; Talavera, M. Biological protection conferred by Glomus spp. and Bacillus megaterium against Meloidogyne incognita in tomato and pepper. IOBC/WPRS Bull. 2012, 83, 215–218. [Google Scholar]
- Turatto, M.F.; Dourado, F.d.S.; Zilli, J.E.; Botelho, G.R. Control potential of Meloidogyne javanica and Ditylenchus spp. using fluorescent Pseudomonas and Bacillus spp. Braz. J. Microbiol. 2018, 49, 54–58. [Google Scholar] [CrossRef]
- Singh, N.; Kumar, S.; Bajpai, V.K.; Dubey, R.C.; Maheshwari, D.K.; Kang, S.C. Biological control of Macrophomina phaseolina by chemotactic fluorescent Pseudomonas aeruginosa PN1 and its plant growth promotory activity in Chir-pine. Crop. Prot. 2010, 29, 1142–1147. [Google Scholar] [CrossRef]
- Yin, N.; Zhao, J.-L.; Liu, R.; Li, Y.; Ling, J.; Yang, Y.-H.; Xie, B.-Y.; Mao, Z.-C. Biocontrol Efficacy of Bacillus cereus Strain Bc-cm103 Against Meloidogyne incognita. Plant Dis. 2021, 105, 2061–2070. [Google Scholar] [CrossRef]
- Egan, A.; Kakouli-Duarte, T. Observations on the interaction between plant growth-promoting bacteria and the root-knot nematode Meloidogyne hapla. MicrobiologyOpen 2022, 11, e1319. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Treatment Code | BCO | SBP |
|---|---|---|
| Ø-Ø | Ø | Ø |
| Ø-FC37 | B. velezensis FC37 | Ø |
| Ø-Hvs8 | B. frigoritolerans Hvs8 | Ø |
| Ø-AC17 | P. aeruginosa AC17 | Ø |
| MH-Ø | Ø | M. hapla |
| MH-FC37 | B. velezensis FC37 | M. hapla |
| MH-Hvs8 | B. frigoritolerans Hvs8 | M. hapla |
| MH-AC17 | P. aeruginosa AC17 | M. hapla |
| MP-Ø | Ø | M. phaseolina |
| MP-FC37 | B. velezensis FC37 | M. phaseolina |
| MP-HVS8 | B. frigoritolerans Hvs8 | M. phaseolina |
| MP-AC17 | P. aeruginosa AC17 | M. phaseolina |
| Strain Code | Related Species (blast/ncbl) | Accession Number | Similitude % |
|---|---|---|---|
| FC5.2 | Pseudomonas kribbensis | OQ255878 | 99.59 |
| FC10.1 | Xanthomonas translucens | OQ255863 | 99.73 |
| FC10.2 | Cupriavidus metalliduras | OQ255864 | 100 |
| FC14 | Cytobacillus firmus | OQ255865 | 99.52 |
| FC15 | Comamonas testosteroni | OQ255866 | 100 |
| FC17 | Arthrobacter pascens | OQ255867 | 99.59 |
| FC18 | Pararhizobium herbae | OQ255868 | 99.7 |
| FC20 | Pseudomonas brassicacearum subsp. neoaurantiaca | OQ255869 | 99.93 |
| FC21.2 | Agrobacterium arsenijevicii | OQ255870 | 100 |
| FC25 | Xanthomonas translucens | OQ255871 | 99.73 |
| FC28 | Pseudomonas flavescens | OQ255872 | 98.83 |
| FC32 | Bacillus albus | OQ255873 | 100 |
| FC34 | Arthrobacter humicola | OQ255874 | 99.31 |
| FC35 | Pseudomonas nitritireducens | OQ255875 | 99.86 |
| FC36 | Arthrobacter pascens | OQ255876 | 99.45 |
| FC37 | Bacillus velezensis | OU487633 | * |
| FC39 | Raoultella ornithinolytica | OQ255877 | 99.86 |
| AC9.1 | Priestia aryabhattai | OQ255855 | 100 |
| AC11.2 | Bacillus altitudinis | OQ255851 | 100 |
| AC14 | Mixta calida | OQ255852 | 100 |
| AC15 | Enterobacter quasiroggenkampii | OQ255853 | 99.86 |
| AC17 | Pseudomonas aeruginosa | OQ255854 | 100 |
| ACH2 | Chryseobacterium cucumelis | OQ255860 | 99.93 |
| ACH10 | Klebsiella variicola subsp. variicola | OQ255856 | 99.73 |
| ACH14 | Achromobacter veterisilvaer | OQ255857 | 99.93 |
| ACH15 | Microbacterium arborescens | OQ255858 | 99.93 |
| ACH16 | Bacillus velezensis | OQ255859 | 99.93 |
| ACH25 | Enterobacter kobei | OQ255861 | 99.93 |
| ACH29 | Curtobacterium citreum | OQ255862 | 99.72 |
| Hvs8 | Brevibacterium frigoritolerans | OU487634 | * |
| Treatment Code | Dead Plants(%) | Plant Severity Index | Crown Incidence (%) | Crown Severity Index | Petiole Colonization (%) | Root Colonization (%) |
|---|---|---|---|---|---|---|
| MP-Ø | 12.5 ± 8.5 a | 2.9 ± 0.2 a | 75.0 ± 11.2 a | 1.6 ± 0.3 a | 75.0 ± 11.2 a | 75.0 ± 11.2 a |
| MP-Hvs8 | 18.8 ± 10.0 a | 2.0 ± 0.3 b | 18.8 ± 10.0 b | 0.8 ± 0.4 ab | 12.5 ± 8.5 b | 50.0 ± 12.9 a |
| MP-AC17 | 0.0 ± 0.0 b | 1.5 ± 0.1 b | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 53.3 ± 13.3 a |
| MP-FC37 | 6.3 ± 6.1 a | 1.9 ± 0.2 b | 31.3 ± 11.9 b | 0.6 ± 0.3 b | 31.3 ± 11.9 b | 43.8 ± 12.8 a |
| Treatment Code | Galling Index | RKN Per Pot (750 mL) |
|---|---|---|
| MH-Ø | 2.4 ± 0.1 a | 3033 ± 172 a |
| MH-Hvs8 | 2.2 ± 0.1 a | 2819 ± 214 a |
| MH-AC17 | 0.9 ± 0.9 b | 1518 ± 130 b |
| MH-FC37 | 2.3 ± 0.2 b | 2708 ± 121 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Camacho, M.; de los Santos, B.; Vela, M.D.; Talavera, M. Use of Bacteria Isolated from Berry Rhizospheres as Biocontrol Agents for Charcoal Rot and Root-Knot Nematode Strawberry Diseases. Horticulturae 2023, 9, 346. https://doi.org/10.3390/horticulturae9030346
Camacho M, de los Santos B, Vela MD, Talavera M. Use of Bacteria Isolated from Berry Rhizospheres as Biocontrol Agents for Charcoal Rot and Root-Knot Nematode Strawberry Diseases. Horticulturae. 2023; 9(3):346. https://doi.org/10.3390/horticulturae9030346
Chicago/Turabian StyleCamacho, María, Berta de los Santos, María Dolores Vela, and Miguel Talavera. 2023. "Use of Bacteria Isolated from Berry Rhizospheres as Biocontrol Agents for Charcoal Rot and Root-Knot Nematode Strawberry Diseases" Horticulturae 9, no. 3: 346. https://doi.org/10.3390/horticulturae9030346