
Changes in Carbohydrates, Organic Acids, and Minerals at Different Development Stages of Hexachlamys edulis Fruit, a Wild South American Species with Horticultural Potential
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growing Conditions
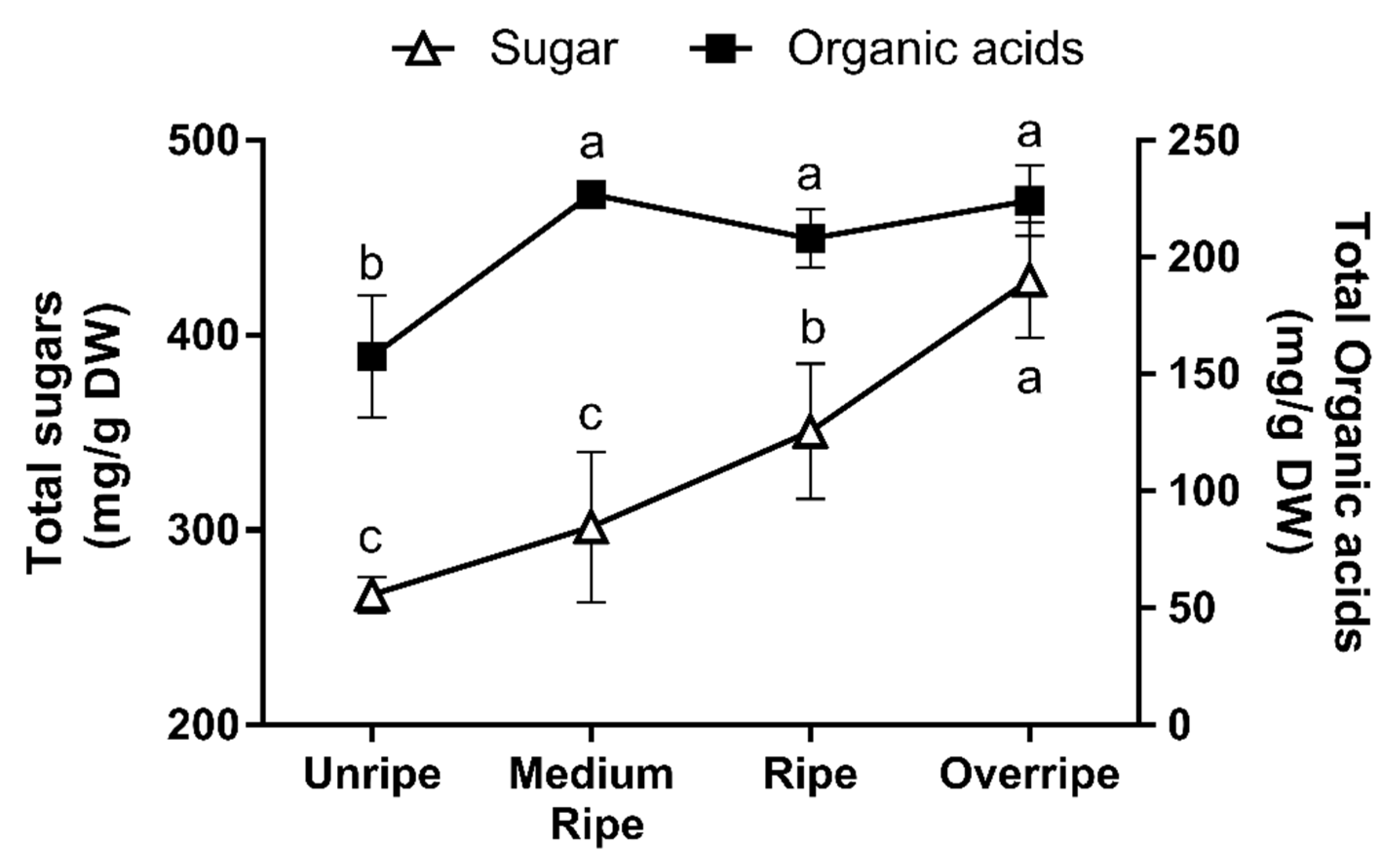
2.2. Analysis of Carbohydrates and Organic Acids
2.3. Analysis of Minerals
2.4. Statistical Analysis
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hossain, M.M.; Rahim, M.A.; Haque, M.R. Biochemical properties of some important underutilized minor fruits. J. Agric. Food Res. 2021, 5, 100148. [Google Scholar] [CrossRef]
- Murthy, H.N.; Bapat, V.A. Importance of Underutilized Fruits and Nuts. In Bioactive Compounds in Underutilized Fruits and Nuts; Murthy, H., Bapat, V., Eds.; Reference Series in Phytochemistry; Springer: Cham, Switzerland, 2020. [Google Scholar] [CrossRef]
- Lamani, S.; Anu-Appaiah, K.A.; Murthy, H.N.; Dewir, Y.H.; Rikisahedew, J.J. Analysis of Free Sugars, Organic Acids, and Fatty Acids of Wood Apple (Limonia acidissima L.) Fruit Pulp. Horticulturae 2022, 8, 67. [Google Scholar] [CrossRef]
- Gordon, A. Bioactive Compounds in Underutilized Tropical Fruits from Latin America. Ph.D. Dissertation, Universitäts-und Landesbibliothek, Bonn, Germany, 2012. Available online: https://nbn-resolving.org/urn:nbn:de:hbz:5n-29864 (accessed on 2 December 2022).
- Nandal, U.; Bhardwaj, R.L. The role of underutilized fruits in nutritional and economic security of tribals: A review. Crit. Rev. Food Sci. Nutr. 2014, 54, 880–890. [Google Scholar] [CrossRef] [PubMed]
- Barua, U.; Das, R.P.; Gogoi, B.; Baruah, S.R. Underutilized fruits of Assam for livelihood and nutritional security. Agric. Rev. 2019, 40, 175–184. [Google Scholar] [CrossRef] [Green Version]
- Povilonis, I.S.; Arena, M.E.; Radice, S. Hexachlamys edulis (Berg) Kausel & Legrand, “ubajay”, a native fruit species from South America. AHS 2021, 35, 389–397. [Google Scholar] [CrossRef]
- Arena, M.E.; Povilonis, I.S.; Borroni, V.; Constenla, D.; Radice, S. Changes in physicochemical properties at different development stages of Hexachlamys edulis fruit, an underutilized South American species. Heliyon 2021, 7, e08323. [Google Scholar] [CrossRef]
- Britton, G.; Khachik, F. Carotenoids in Food. In Carotenoids: Nutrition and Health; Britton, G., Pfander, H., Liaaen-Jensen, S., Eds.; Birkhäuser: Basel, Switzerland, 2009; Volume 5, pp. 45–66. ISBN 978-3-7643-7501-0. [Google Scholar] [CrossRef]
- Povilonis, I.S.; Arena, M.E.; Radice, S. Caracterización de la variabilidad fenotípica de los frutos de ubajay (Hexachlamys edulis). In Proceedings of the 41° Congreso Argentino de Horticultura, La Plata, Argentina, 5–8 October 2021; Available online: http://sedici.unlp.edu.ar/bitstream/handle/10915/128494/Documento_completo.pdf-PDFA.pdf?sequence=1&isAllowed=y (accessed on 15 November 2022).
- Radice, S.; Povilonis, I.S.; Arena, M.E. Flower and fruit formation of Hexachlamys edulis in Buenos Aires, Argentina. JAEID 2023, in press. [Google Scholar]
- Povilonis, I.S.; Arena, M.E.; Radice, S. Efecto de la temperatura de conservación sobre propiedades químicas de los frutos de Hexachlamys edulis en Concordia, Entre Ríos. In Proceedings of the VIII Congreso Internacional de Ciencia y Tecnología de Alimentos (CICYTAC), Cordoba, Argentina, 4–6 October 2022. [Google Scholar]
- Batista-Silva, W.; Nascimento, V.L.; Medeiros, D.B.; Nunes-Nesi, A.; Ribeiro, D.M.; Zsögön, A.; Araújo, W.L. Modifications in Organic Acid Profiles during Fruit Development and Ripening: Correlation or Causation? Front. Plant Sci. 2018, 9, 1689. [Google Scholar] [CrossRef]
- Shi, Y.; Li, B.J.; Su, G.; Zhang, M.; Grierson, D.; Chen, K.S. Transcriptional regulation of fleshy fruit texture. J. Integr. Plant Biol. 2022, 64, 1649–1672. [Google Scholar] [CrossRef]
- Xie, F.; Chen, C.; Chen, J.; Yuan, Y.; Hua, Q.; Zhang, Z.; Zhao, J.; Hu, G.; Chen, J.; Qin, Y. Metabolic Profiling of Sugars and Organic Acids, and Expression Analyses of Metabolism-Associated Genes in Two Yellow-Peel Pitaya Species. Plants 2022, 11, 694. [Google Scholar] [CrossRef]
- Zhi, W.S.; Li, Y.F.; Zi, S.W.; Xue, X.B. Sugar transport, metabolism, accumulation and their regulation in fruits. J. Plant Physiol. Mol. Biol. 2004, 30, 1–10. [Google Scholar] [PubMed]
- Vicente, A.R.; Manganaris, G.A.; Sozzi, G.O.; Crisosto, C.H. Nutritional quality of fruits and vegetables. In Postharvest Handling: A Systems Approach, 2nd ed.; Florkowski, W.J., Shewfelt, R.L., Brueckner, B., Prussia, S.E., Eds.; 2009; pp. 57–106. ISBN 978-0-12-374112-7. [Google Scholar]
- Ahmad, A.; Khalid, N. Chapter 5—Dietary Fibers in Modern Food Production: A Special Perspective with β-Glucans. In Biopolymers for Food Design; Grumezescu, A.M., Holban, A.M., Eds.; Academic Press: Philadelphia, PA, USA, 2018; pp. 125–156. ISBN 978-0-12-811449-0. [Google Scholar] [CrossRef]
- EI-Zoghbi, M. Biochemical changes in some tropical fruits during ripening. Food Chem. 1994, 49, 33–37. [Google Scholar] [CrossRef]
- Walker, R.P.; Famiani, F. Organic Acids in Fruits: Metabolism, Functions and Contents. In Horticultural Review, 1st ed.; Warrington, I., Ed.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2018; pp. 371–430. ISBN 978-1-119-43107-7. [Google Scholar]
- Paul, V.; Pandey, R.; Ramesh, K.V.; Singh, A. Role of mineral nutrients in physiology, ripening and storability of fruits. In Advances in Plant Physiology; Hemantaranjan, A., Ed.; An International Treatise Series: Nutriophysiological and Molecular Interventions for crop improvement under changing climate; Scientific Publishers: Jodhpur, India, 2012; Volume 13, pp. 56–96. [Google Scholar]
- Rahman, M.M.; Roy, M.; Sajib, M.A.M.; Sarkar, A.; Hussain, M.S. Radiation effects on essential minerals content of cucumber (Cucumis sativus). Amer. J. Food Nutr. 2015, 3, 69–74. [Google Scholar]
- Tyagi, S.; Sahay, S.; Imran, M.; Rashmi, K.; Mahesh, S.S. Pre-harvest factors influencing the postharvest quality of fruits: A review. Curr. J. Appl. Sci. Technol. 2017, 23, 12. [Google Scholar] [CrossRef]
- Taghavi, T.; Siddiqui, R.; Rutto, L.K. The effect of preharvest factors on fruit and nutritional quality in strawberry. In Strawberry Pre-and Post-harvest Management Techniques for Higher Fruit Quality; IntechOpen: London, UK, 2019. [Google Scholar]
- Colaric, M.; Stampar, F.; Solar, A.; Hudina, M. Influence of branch bending on sugar, organic acid and phenolic content in fruits of ‘Williams’ pears (Pyrus communis L.). J. Sci. Food Agric. 2006, 86, 2463–2467. [Google Scholar] [CrossRef]
- Lage-Yusty, M.; Simal-Lozano, J.; Gómez, G.S. Determinación de almidón en alimentos. Trab. Compostel. Biol. 1983, 10, 15–25. [Google Scholar]
- Megazyme. Total Dietary Fiber. Assay Procedure. 2017. Available online: https://www.megazyme.com/documents/Assay_Protocol/K-TDFR-200A_DATA.pdf (accessed on 10 June 2022).
- González-Agüero, M.; Tejerina Pardo, L.; Zamudio, M.S.; Contreras, C.; Undurraga, P.; Defilippi, B.G. The Unusual Acid-Accumulating Behavior during Ripening of Cherimoya (Annona cherimola Mill.) is Linked to Changes in Transcription and Enzyme Activity Related to Citric and Malic Acid Metabolism. Molecules 2016, 21, 398. [Google Scholar] [CrossRef] [Green Version]
- Carnelossi, M.A.G.; Costa de Sena, H.; Narain, N.; Yaguiu, P.; da Silva, G.F. Physico-Chemical Quality Changes in Mangaba (Hancornia speciosa gomes) Fruit Stored at Different Temperatures. Braz. Arch. Biol. Technol. 2009, 52, 985–990. [Google Scholar] [CrossRef] [Green Version]
- Rogiers, S.Y.; Knowles, N.R. Physical and chemical changes during growth, maturation, and ripening of Saskatoon (Amelanchier alnifolia) fruit. Can. J. Bot. 1997, 75, 1215–1225. [Google Scholar] [CrossRef]
- Pareek, S. Chapter 17—Nutritional and Biochemical Composition of Lychee (Litchi chinensis Sonn.) Cultivars. In Nutritional Composition of Fruit Cultivars; Simmonds, M.S.J., Preedy, V.R., Eds.; Academic Press: Cambridge, MA, USA, 2016; pp. 395–418. ISBN 9780124081178. [Google Scholar] [CrossRef]
- Zhao, A.-L.; Xue, X.-F.; Wang, Y.-K.; Sui, C.-L.; Ren, H.-Y.; Li, D.-K. The Sugars and Organic Acids Composition in Fruits of Different Chinese Jujube Cultivars of Different Development Stages. Acta Hortic. Sin. 2016, 43, 1175–1185. [Google Scholar] [CrossRef]
- Albertini, M.-V.; Carcouet, E.; Pailly, O.; Gambotti, C.; Luro, F.; Berti, L. Changes in organic acids and sugars during early stages of development of acidic and acidless citrus fruit. J. Agric. Food Chem. 2006, 54, 8335–8339. [Google Scholar] [CrossRef]
- Bae, H.; Yun, S.K.; Jun, J.H.; Yoon, I.K.; Nam, E.Y.; Kwon, J.H. Assessment of organic acid and sugar composition in apricot, plumcot, plum, and peach during fruit development. J. Appl. Bot. Food Qual. 2014, 87, 24–29. [Google Scholar] [CrossRef]
- Mao, Y.; Tian, S.; Qin, Y.; Cheng, S. An optimized organic acid human sensory sourness analysis method. J. Sci. Food Agric. 2021, 101, 5880–5887. [Google Scholar] [CrossRef]
- Aslam, M.M.; Deng, L.; Wang, X.; Wang, Y.; Pan, L.; Liu, H.; Niu, L.; Lu, Z.; Cui, G.; Zeng, W.; et al. Expression patterns of genes involved in sugar metabolism and accumulation during peach fruit development and ripening. Sci. Hortic. 2019, 257, 108633. [Google Scholar] [CrossRef]
- Zhang, W.; Lunn, J.E.; Feil, R.; Wang, Y.; Zhao, J.; Tao, H.; Guo, Y.; Zhao, Z. Trehalose 6-phosphate signal is closely related to sorbitol in apple (Malus domestica Borkh. cv. Gala). Biol. Open 2017, 6, 260–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Wang, J.; Mujumdar, A.S.; Jin, X.; Liu, Z.L.; Zhang, Y.; Xiao, H.W. Effects of postharvest ripening on physicochemical properties, microstructure, cell wall polysaccharides contents (pectin, hemicellulose, cellulose) and nanostructure of kiwifruit (Actinidia deliciosa). Food Hydrocoll. 2021, 118, 106808. [Google Scholar] [CrossRef]
- Desnoues, E.; Gibon, Y.; Baldazzi, V.; Signoret, V.; Génard, M.; Quilot-Turion, B. Profiling sugar metabolism during fruit development in a peach progeny with different fructose-to-glucose ratios. BMC Plant Biol. 2014, 14, 336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trad, M.; Boge, M.; Hamda, H.B.; Renard, C.M.G.C.; Harbi, M. The Glucose-Fructose ratio of wild Tunisian grapes. Cogent Food Agric. 2017, 3, 1374156. [Google Scholar] [CrossRef]
- Song, J.; Bi, J.; Chen, Q.; Wu, X.; Lyu, Y.; Meng, X. Assessment of sugar content, fatty acids, free amino acids, and volatile profiles in jujube fruits at different ripening stages. Food Chem. 2019, 270, 344–352. [Google Scholar] [CrossRef]
- Osorio, S.; Scossa, F.; Fernie, A.R. Molecular regulation of fruit ripening. Front. Plant Sci. 2013, 14, 198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chervin, C. Should Starch Metabolism Be a Key Point of the Climacteric vs. Non-climacteric Fruit Definition? Front. Plant Sci. 2020, 11, 609189. [Google Scholar] [CrossRef]
- Ikegaya, A.; Toyoizumi, T.; Ohba, S.; Nakajima, T.; Kawata, T.; Ito, S.; Arai, E. Effects of distribution of sugars and organic acids on the taste of strawberries. Food Sci. Nutr. 2019, 7, 2419–2426. [Google Scholar] [CrossRef]
- Kader, A.A. Flavor quality of fruits and vegetables. J. Sci. Food Agric. 2008, 88, 1863–1868. [Google Scholar] [CrossRef]
- Baccichet, I.; Chiozzotto, R.; Bassi, D.; Gardana, C.; Cirilli, M.; Spinardi, A. Characterization of fruit quality traits for organic acids content and profile in a large peach germplasm collection. Sci. Hortic. 2021, 278, 109865. [Google Scholar] [CrossRef]
- Nadeem, M.; Qureshi, T.M.; Ugulu, I.; Riaz, M.N.; An, Q.U.; Khan, Z.I.; Dogan, Y. Mineral, vitamin and phenolic contents and sugar profiles of some prominent date palm (Phoenix dactylifera) varieties of Pakistan. Pak. J. Bot. 2019, 51, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Czech, A.; Zarycka, E.; Yanovych, D.; Zasadna, Z.; Grzegorczyk, I.; Kłys, S. Mineral content of the pulp and peel of various citrus fruit cultivars. Biol. Trace Elem. Res. 2020, 193, 555–563. [Google Scholar] [CrossRef] [Green Version]
- Aburto, N.J.; Hanson, S.; Gutierrez, H.; Hooper, L.; Elliott, P.; Cappuccio, F.P. Effect of increased potassium intake on cardiovascular risk factors and disease: Systematic review and meta-analyses. BMJ 2013, 346, f1378. [Google Scholar] [CrossRef] [Green Version]
- DiNicolantonio, J.J.; Liu, J.; O’Keefe, J.H. Magnesium for the prevention and treatment of cardiovascular disease. Open Heart 2018, 5, e000775. [Google Scholar] [CrossRef] [Green Version]
- WHO. Potassium Intake for Adults and Children; WHO: Geneva, Switzerland, 2012; pp. 1–52. ISBN 978-92-4-150482-9. Available online: https://www.who.int/publications/i/item/9789241504829 (accessed on 20 January 2023).
- World Health Organization. Vitamin and Mineral Requirements in Human Nutrition; World Health Organization: Geneva, Switzerland, 2004; Available online: https://www.fao.org/3/y2809e/y2809e00.htm (accessed on 20 January 2023).



{kind=link}
{kind=link}
| Factor | TAR (mg/g) | MAL (mg/g) | QUI (mg/g) | SUC (mg/g) |
|---|---|---|---|---|
| Stages | ||||
| Unripe | 0.00 ± 0.10 b | 44.90 ± 8.33 c | 0.37 ± 0.24 b | 112.33 ± 3.31 a |
| Medium Ripe | 0.00 ± 0.10 b | 90.19 ± 2.85 b | 25.25 ± 1.80 a | 92.48 ± 3.31 b |
| Ripe | 0.00 ± 0.10 b | 105.14 ± 1.95 a | 22.62 ± 1.09 a | 99.43 ± 3.31 b |
| Overripe | 0.94 ± 0.10 a | 84.79 ± 2.34 b | 22.85 ± 1.78 a | 115.65 ± 3.31 a |
| F | 26.039 | 77.887 | 262.200 | 12.398 |
| p | <0.001 | <0.001 | <0.001 | <0.001 |
| Factor | SUC (mg/g) | GLU (mg/g) | FRU (mg/g) | GLU/ FRU | TS/TA | STR (mg/g) |
|---|---|---|---|---|---|---|
| Stages | ||||||
| Unripe | 21.20 ± 4.80 b | 95.59 ± 2.42 b | 150.08 ± 6.50 bc | 0.63 ± 0.01 b | 1.72 ± 0.09 ab | 171.39 ± 1.93 a |
| Medium Ripe | 77.79 ± 4.80 a | 89.49 ± 5.90 b | 134.40 ± 6.50 c | 0.66 ± 0.00 b | 1.44 ± 0.09 b | 161.19 ± 1.93 a |
| Ripe | 82.53 ± 4.80 a | 105.00 ± 77.59 b | 163.08 ± 6.50 b | 0.64 ± 0.00 b | 1.54 ± 0.09 ab | 40.45 ± 1.93 b |
| Overripe | 59.38 ± 4.80 a | 163.13 ± 6.57 a | 205.85 ± 6.50 a | 0.79 ± 0.00 a | 1.90 ± 0.09 a | 65.96 ± 1.93 b |
| F | 33.672 | 42.000 | 33.660 | 118.100 | 4.972 | 11.705 |
| p | <0.001 | <0.001 | <0.001 | <0.001 | 0.045 | 0.003 |
| Factor | IDF (g/100 g) | SDF (g/100 g) | TDF (g/100 g) |
|---|---|---|---|
| Stages | |||
| Unripe | 26.71 ± 1.68 a | 9.03 ± 1.35 | 35.74 ± 2.22 ab |
| Medium Ripe | 27.13 ± 1.93 a | 9.67 ± 1.35 | 36.80 ± 2.22 a |
| Ripe | 15.81 ± 1.93 b | 11.26 ± 1.35 | 27.07 ± 2.22 ab |
| Overripe | 15.51 ± 1.93 b | 10.44 ± 1.35 | 25.94 ± 2.22 b |
| F | 14.895 | 0.505 | 6.511 |
| P | 0.001 | 0.689 | 0.015 |
| Factor | Mg (mg/g) | K (mg/g) | Mn (mg/g) | TC (mg/g) |
|---|---|---|---|---|
| Stages | ||||
| Unripe | 0.33 ± 0.10 | 20.63 ± 7.63 | 0.00 ± 0.00 | 20.97 ± 7.74 |
| Medium Ripe | 0.34 ± 0.03 | 20.69 ± 2.55 | 0.01 ± 0.01 | 21.05 ± 2.57 |
| Ripe | 0.40 ± 0.02 | 22.85 ± 2.73 | 0.00 ± 0.00 | 23.27 ± 2.76 |
| Overripe | 0.24 ± 0.04 | 17.57 ± 3.14 | 0.00 ± 0.00 | 17.82 ± 3.17 |
| F | 3.559 | 0.761 | 2.815 | 0.787 |
| p | 0.087 | 0.556 | 0.130 | 0.543 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arena, M.E.; Povilonis, I.S.; Borroni, V.; Pérez, E.; Pellegrino, N.; Cacciatore, C.; Radice, S. Changes in Carbohydrates, Organic Acids, and Minerals at Different Development Stages of Hexachlamys edulis Fruit, a Wild South American Species with Horticultural Potential. Horticulturae 2023, 9, 314. https://doi.org/10.3390/horticulturae9030314
Arena ME, Povilonis IS, Borroni V, Pérez E, Pellegrino N, Cacciatore C, Radice S. Changes in Carbohydrates, Organic Acids, and Minerals at Different Development Stages of Hexachlamys edulis Fruit, a Wild South American Species with Horticultural Potential. Horticulturae. 2023; 9(3):314. https://doi.org/10.3390/horticulturae9030314
Chicago/Turabian StyleArena, Miriam Elisabet, Ignacio Sebastián Povilonis, Virginia Borroni, Ethel Pérez, Néstor Pellegrino, Claudio Cacciatore, and Silvia Radice. 2023. "Changes in Carbohydrates, Organic Acids, and Minerals at Different Development Stages of Hexachlamys edulis Fruit, a Wild South American Species with Horticultural Potential" Horticulturae 9, no. 3: 314. https://doi.org/10.3390/horticulturae9030314