
Large-Scale In Vitro Propagation and Ex Vitro Adaptation of the Endangered Medicinal Plant Eryngium maritimum L.

1
Institute for Environmental Solutions, “Lidlauks”, Priekuļi Parish, LV-4126 Cēsis County, Latvia
2
The National Botanic Garden of Latvia, Miera Iela 1, LV-2169 Salaspils, Latvia
3
Field and Forest, SIA, 2 Izstades Str., Priekuli Parish, LV-4126 Cēsis County, Latvia
*
Author to whom correspondence should be addressed.
Horticulturae 2023, 9(2), 271; https://doi.org/10.3390/horticulturae9020271
Submission received: 12 January 2023
/
Revised: 13 February 2023
/
Accepted: 14 February 2023
/
Published: 17 February 2023
(This article belongs to the Special Issue A New Decade in the Propagation of Horticultural and Medicinal Plants)
Abstract
:The endangered medicinal plant Eryngium maritimum L. faces significant natural and anthropogenic threats. Therefore, in vitro propagation is recommended for both conservation and commercial purposes. The aim of the study was to develop a series of protocols for seed disinfection, in vitro multiplication and rooting, and ex vitro and field adaptation. For explant disinfection, the length and temperature of three consecutive disinfectants were investigated. Macrosalt modifications of MS medium and plant growth regulator addition to media effect on axillary bud propagation rate and rooting was studied. Survival and leaf growth during ex vitro and field adaptation in response to potting media, pot cell diameter, and light spectrum were tested. Seeds treated with diluted detergent at 40 °C for 180 min, followed by 0.01% KMnO4 for 3 h and commercial bleach for 11 min, achieved a lower rate of contamination and high germination rate. Axillary bud proliferation and rooting were enhanced by reducing nitrogen content in media and adding plant growth regulators. Potting media and pot size affect survival and growth ex vitro. Timely transplantation to field conditions before overwintering increases plantlet survival. In the present work, a suitable foundation is laid to scaleup the production of E. maritimum by micropropagation.
1. Introduction
Sea holly (Eryngium maritimum L.) belongs to the genus Eryngium, which unites approximately 230 different species widely distributed throughout the world, especially in Asia and Africa. Sea holly inhabits coastal areas along the Black Sea, the Mediterranean Sea, the North Sea, and the Baltic Sea, as well as the coasts of the Atlantic Ocean in North Africa and Europe [1]. E. maritimum is used in European medicine as a cough suppressant, aphrodisiac, and diuretic, as well as in cases of liver and kidney disease [2]. It is a source of various secondary metabolites, such as essential oils [2,3,4,5,6], triterpenoid saponins [7], phenolic acids, and flavonoids [8].
Conservation and reproduction in vitro are especially important for protected and endangered species that might not be sufficiently conserved in their natural habitats or in collections [9]. Plant species are preferably conserved in situ by protecting habitats and applying species-specific management practices in the wild. If in situ conservation strategies are not sufficient, alternative ex situ strategies may be applied, e.g., propagation in vitro, cryopreservation, or seed banking [10,11,12]. Even though seed banking is suggested as the most suitable method for long-term seed conservation, it is not recommended for species with highly dormant seeds [11]. As in many species of the Apiaceae family, E. maritimum has poor and non-uniform germination [13]; thus, in vitro propagation is a good alternative for conservation. With this technology, a large number of genetically homogeneous plants can be rapidly produced [14], and plants with commercially important traits, e.g., a desired chemical profile, can be selected and propagated [15]. In vitro technologies have been proven efficient for multiplying plant species of medicinal value, including those of the Eryngium genus [7,10,16,17,18].
The first step is plant tissue culture for viable explant establishment in aseptic conditions. At this stage, explants must be disinfected to ensure aseptic growth conditions, and, at the same time, the growth of the explant must be achieved [10]. Seeds, excised embryos; apical, axillary, and lateral buds; young shoots, and young leaves have been used as explants in the Eryngium genus [7,18,19,20,21]. Initiation of plant tissue cultures from seeds is recommended to maintain high genetic diversity within populations. Genetic diversity is essential to establishing viable populations for conservation purposes and reintroduction [22,23], and the most suitable individuals for desired commercial purposes are to be selected [24]. Previous attempts [7] have failed to introduce E. maritimum to in vitro conditions through seed explants, as only 5% of seeds germinated, and the emerged seedlings did not develop into healthy plants. Furthermore, severe contamination of seeds has been reported in the Eryngium genus (60–100%) [10], and only the implementation of protocols with multiple steps of the disinfectant application has succeeded in eliminating infection [10,18]. The disinfection must eliminate microorganisms on the seed surface while maintaining seed vitality. The compounds, their concentrations, and their application length must be adapted for each species based on seed morphology [25], the microorganisms present [26], as well as the mode of action of the disinfectant [27]. Various disinfectant agents are commonly used for seed decontamination, such as sodium and calcium hypochlorite (NaOCl, Ca(OCl)2) [28,29], ethanol [27], mercuric chloride (HgCl2) [30], hydrogen peroxide (H2O2) [31], silver nitrate (AgNO3) [29,32], and different detergents [18,25,30]. Less-known disinfection methods with both a stimulating effect on germination and a deleterious effect on microflora include seed treatment with warm water (up to 65 °C) [20] and seed immersion in potassium permanganate [33,34].
Plant tissue culture provides plants with all necessary conditions for growth, including optimal light and temperature, as well as a basal medium containing all necessary macronutrients, micronutrients, and water [35,36]. Murashige–Skoog [37] (MS) medium is the most commonly used basal medium. It has high ammonium and nitrate levels and provides an optimal nutrient composition for most plant tissue culture applications. When lower nitrogen levels need to be provided, macrosalt and microsalt levels in MS media are often reduced [36]. For example, in the rooting stage, a reduced salt concentration may contribute to better root formation [38]. This approach has been applied in the propagation stage of Eryngium shoot cultures [7,17,21].
Basal media can be supplemented with plant growth regulators (PGR) to manipulate plant development [7,17,21]. In plant tissue cultures, auxins and cytokinins are the most commonly used PGRs to promote shoot proliferation [39]. Cytokinins promote mitotic cell division in the shoots and regulate the morphogenesis of new shoot apical meristems [39]; therefore, they are widely used to enhance shoot proliferation [39,40]. The rate at which new shoots form depends on the type and amount of cytokinin used. It is species-specific, and concentrations below or above the optimal result in a lower shoot formation rate [41]. Auxins, on the other hand, stimulate root induction and increase the quality of roots [38,42]. Naphthaleneacetic acid (NAA), indole-3-acetic acid (IAA), and indole-3-butyric acid (IBA) are the most widely used auxins. The plant response to each of the auxins and their concentrations is species-dependent [42]. In E. maritimum, E. planum, E. campestre, and E. alpinum, the addition of the cytokinine 6-benzylaminopurine (BAP) alone or in addition to the auxins IAA or IBA have been demonstrated to induce high axillary shoot proliferation [7,10,16,17,18]. Reduction of the salt concentration in MS media and the addition of auxins has been proven efficient in the Eryngium genus, e.g., NAA addition while rooting for E. alpinum and E. planum [18,43], and IAA for E. maritimum and E. campestre [7,17].
In vitro culture provides plants with a stable temperature, optimal light conditions, and all necessary nutrients and sugars via the medium, thereby reducing the need for photosynthesis. The transition to the ex vitro environment requires suitable microenvironmental factors in order to develop photoautotrophic plantlets [14]. Monitoring the quantity, quality, and direction of light is of uttermost importance for the survival of photoautotrophic organisms. Several photoreceptors belonging to three principal families—phytochromes, cryptochromes, and phototropins—collect light stimuli from the broad light spectrum (280–750 nm) [44]. The ability to sense light signals allows plants to regulate responses in the complex phenomenon of photomorphogenesis (including in vitro and ex vitro growth) [45]. The effect of the artificial light spectrum on plant biomass accumulation and morphogenesis has been widely researched [46,47,48].
The spectral quality of light influences plant development in vitro and ex vitro [49]. Initial ex vitro stages under controlled conditions under artificial light can positively impact plant growth when compared to natural lighting conditions in a greenhouse [47]. Various artificial light sources, such as fluorescent lamps [49], high-pressure sodium lamps [50], and light-emitting diodes (LEDs), can be used for plant cultivation indoors [51]. Each type of lamp has deviations in light intensity and quality from natural light. The differences are more pronounced in the far-red range [46]. Blue and red LEDs are primarily used for plant cultivation as their spectral power distribution is optimal for photosynthesis [52]. The addition of far-red and green lights to blue and red ones can increase biomass accumulation and modify morphogenesis during the ex vitro adaptation stage [53].
The composition of a horticultural substrate replacing soil can affect plant growth and morphological development [47,54]. Substrates from multiple ingredients are prepared for ex vitro acclimatization to ensure the best conditions for gradual root maturation [55]. In most cases, substrates are prepared from peat, vermiculite, perlite, and sand [55,56]. The addition of a nutrient source, e.g., mineral fertilizers or compost, improves plant development during ex vitro adaptation [47,57]. The addition of cow manure to substrate mixtures is an option to provide a nutrient source, stabilize the substrate pH, and suppress pathogen development [56]. However, no data is available on the effect of cow manure addition to substrate mixtures on Eryngium ex vitro acclimatization success.
In large-scale propagation, both optimal growing conditions and economic profitability are critical. By lowering maintenance costs per unit of area, reduced container size provides a significant economic benefit. However, by decreasing container size, root development and photosynthetic activity can be reduced, thus affecting plantlet growth [58]. Containers with cell volumes of less than 100 mL are typically used for medicinal plant seedling production [59]. The experiments on Eryngium [7,10] acclimatization have been carried out in pots over 200 mL. Therefore, studies on acclimatization based on industry requirements are needed.
Even though in vitro propagated plantlets can be directly planted in agricultural fields, gradual adaptation to field conditions is preferable to increase survival [47]. Initial plantlet vigor, planting date, shading, soil, and air temperature can influence plantlet survival and development in field conditions [60]. Determining the optimal planting date is critical [61], especially in the condition of Northern Europe, because plants need to develop a robust root system and accumulate a sufficient amount of nutrients for successful overwintering.
The present study was undertaken to identify an efficient protocol for in vitro propagation and ex vitro adaptation of E. maritimum starting from in vitro germinated seed. The effects of prolonged cold storage on seed germination were tested. Additionally, the impact of multistep seed pretreatments on the infection rate was determined. In this study, we expanded the number of previously tested cytokinins added to E. maritimum micropropagation media and tested three different media compositions with varying macrosalt concentrations on in vitro shoot multiplication. We further expanded previous experiments with various IAA concentrations and MS media modifications to determine their effect on in vitro rooting. Moreover, we tested commercially available LED lights with varying light quality to determine if they affect the survival of plantlets within 21 days after transfer ex vitro. We also tested how two potting media and two container sizes affect plantlet survival and leaf development during the ex vitro adaptation stage. Furthermore, we investigated the effects of planting time (June vs. July) and mericlone on plantlet survival after transfer from ex vitro to field conditions.
2. Materials and Methods
2.1. Plant Material
Seeds were either sourced from the wild in Latvia and Estonia, acquired from the University of Pisa botanical garden, or sourced from commercial seed companies. Collection of seeds in the wild took place during the seed ripening stage at the end of August and in September according to permits issued by the Nature Conservation Agency, Republic of Latvia (decision no. 3.6/470/2020-N5 issued on 13.08.2020), and the Environmental Board, Republic of Estonia (decision no. 1-3/20/756 issued at 11.08.2020). In total, seeds from eight different accessions were obtained and subjected to the initiation of plant tissue culture (Table 1).
2.2. In Vitro Germination
Dry seeds were mechanically cleaned, and mature seeds were selected. In total, 252 seeds were tested. Each experimental variant had four replications. Five experimental variants for seed disinfection were tested with varying lengths of detergent, 0.01% KMnO4 solution, bleach treatments, and differing temperatures of detergent application (Table 2) [33,62]. Seeds were submerged in either a 20 °C or 40 °C dilution of the commercial surfactant “Fairy” for 15–180 min and then rinsed for 5 min in water. Then, the seeds were immersed in 0.01% KMnO4 solution [34] for 1–3 h and washed. Subsequently, in sterile conditions, seeds were immersed in commercial sodium hypochlorite solution (“ACE” < 5% sodium hypochlorite) and rinsed in autoclaved distilled water.
Following pericarp removal [18], seeds were placed in a 12 × 110 mm-sized test tube on MS medium (Table S1) with half the nitrate composition (ABMS). Seeds were incubated in a growing chamber for one month at 22 °C with a room moisture level of 70%, a photoperiod of 16/8 h, and a light intensity of 150–200 mol/(m2s). Seeds were stratified at 5 °C for 6 months. After stratification, germination was initiated at 22 °C. Infected seeds were counted and removed on a daily basis, and germinated seeds were counted on days 5, 7, 9, 14, and 20 after the initiation of germination in warm temperatures. Seeds with emerged radicles were considered to have germinated.
2.3. Micropropagation
An axillary branching method was used for shoot multiplication [63]. Each experimental variant consisted of 15 shoots in three replicates. Once the plantlets reached 3–4 cm in length, they were transferred to the micropropagation media. Several experimental variants of micropropagation media were prepared with various amounts and concentrations of plant growth regulators (PGR) and basal media macrosalt. Five different cytokinins (6-benzylaminopurine—BAP, meta-tooling—mT, N6-(2-isopentyl)adenine—2iP, zeatin—Z, kinetin—Kin) were tested at concentrations of 0.0, 0.5, and 1.0 mg L−1. Murashige—Skoog (MS) basal medium [37] and two of its modifications—AMS (MS medium with NH4NO3 concentration reduced in half) and ABMS (MS medium with both NH4NO3 and KNO3 reduced in half)—were utilized. All propagation media contained auxin IAA 0.1 mg L−1 and sucrose 20 g L−1. At the final passage before rooting, an experiment was carried out to determine the quality of the shoots formed in terms of shoot size. Shoots of 10–15 mm in length were considered too small to be subjected to rooting; thus, shoots > 16 mm long were considered to have sufficient quality. The number of experimental variants was reduced by dropping those with the lowest shoot proliferation rate in the previous experiment. Thus, small and optimal axillary shoot formation in response to the addition of BAP and mT at 0.5 and 1.0 mg L−1 to MS, AMS, and AMBS was tested.
2.4. In Vitro Rooting
Healthy shoots at least 3 cm long were selected for rooting. Each experimental variant consisted of 40 shoots in three replicates. Two microsalt compositions were tested: MS media with ammonium nitrate (NH4NO3) concentration reduced by half (AMS) and MS media with both NH4NO3 and potassium nitrate (KNO3)reduced by half (ABMS). The plant growth regulator IAA was added to the media at concentrations of either 0.1 or 0.5 mg L−1. Media with no added PGR was used as a control. Three mericlones of the population collected in Užava, Latvia, were used for testing: M1, M2, and M3.
2.5. Ex Vitro Adaptation
Ex vitro adaptation trials were carried out to test if mericlone, planting substrate, and artificial light quality affect plantlet survival and growth. Two trials were carried out. Each trial consisted of 15 plantlets in three replications. The first trial examined three factors: mericlone, substrate, and light quality. Three mericlones from a single accession from Užava, Latvia, were tested (M1, M2, and M3). Two types of substrates were tested: soil, sand, vermiculite, and perlite at a ratio of 3:1:1:1 (B1); and soil, cow manure, vermiculite, and perlite at a ratio of 3:1:1:1 (B2). Three lamp types were tested: two universal lamps with a full spectrum from 380 to 800 nm (Surya VEG, 30 W [*2], spectra Sun Sky and Heliospectra, 300 W, white spectrum 5700 K), and a lamp with both red (630 nm) and blue (460 nm) light in a ratio of 3:1. Plastic pots sized 6 × 6 × 6 cm were used for plantlet cultivation.
The second trial was based on the results of the first one, with the aim of testing how the diameter of individual cells in the multiple-compartment cell propagation trays affects plantlet survival. Two factors were tested: mericlone (M1 and M2 derived from the accession collected at Užava, Latvia) and pot cell size. Two propagation trays were used: one had a 2.8 cm diameter, 5.0 cm height, and 30 mL volume, and the other had a 3.7 cm diameter, 5.0 cm height, and 54 mL volume. The substrate consisted of a peat-based substrate (Laflora KKS), sand, vermiculite and perlite (3:1:1:1 v/v/v/v). Plantlets were grown under the Heliospectra 300 W white spectrum 5700 K lamp with a 16/8 h photoperiod.
Rooted in vitro plantlets were planted in a soil substrate and sprayed with demineralized water in both trials. The trays were covered with transparent lids and placed in a growth chamber with a temperature of 22 ± 1 °C and a photoperiod of 16/8 h. The relative humidity inside the propagation trays was maintained at 100% for the first five days; then, it was reduced by 10% every five days until it reached 70%. The plastic lid was removed after three weeks of acclimatization, and thereafter, the humidity in the climatic chamber was kept at 70%. Each ex vitro adaptation experiment consisted of 15 plantlets replicated three times.
Two factors were evaluated in the field trials: mericlone (three mericlones—M1, M2, and M3—from the accession collected in Užava, Latvia) and two different planting dates: 11 June 2021 and 20 July 2021. The mericlones M1 and M3 were planted on 11 June 2021. Mericlone M2 was planted on 11 June 2021 and 20 July 2021.
Field trials to assess E. maritimum adaptability to field conditions were carried out in the growing seasons of 2021 and 2022. Trials were set up in an organically certified experimental field at Priekuļi parish, Cēsis county, Latvia (latitude 57°19′11.7″ N, longitude 25°19′18.8″ E, altitude 115 m). A completely randomized block design with three replications and 40 plants per replication was used. Plants were spaced 15 cm apart in 3 × 1.5 m plots, with 75 cm wide inter-rows. The physicochemical properties of the soil and the cultivation practices were the same as in our previous study [64]. After planting, the plants were watered to 100% of the soil’s capacity; no additional watering was applied.
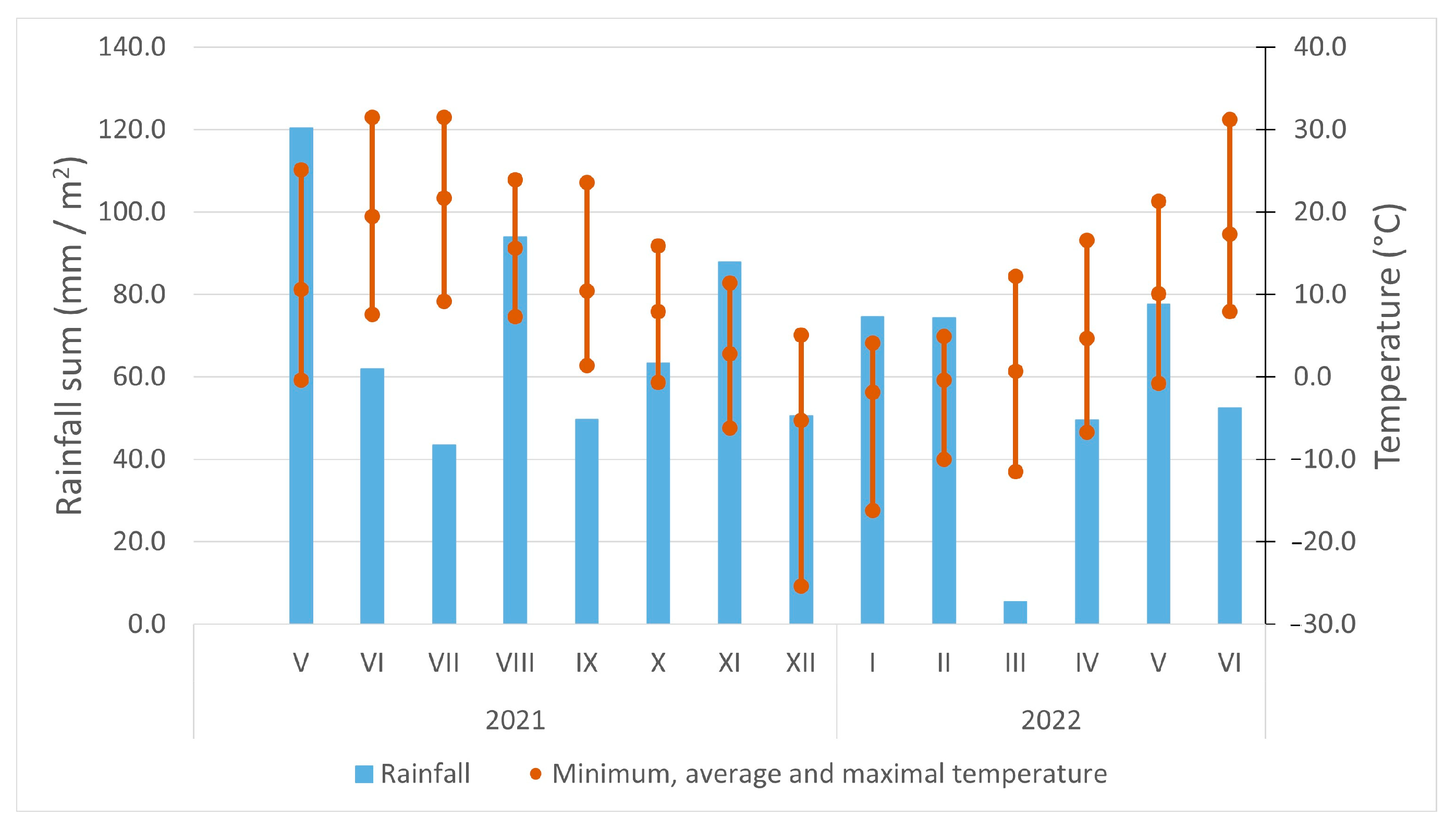
The growing season of 2021 was distinguished by optimum temperature and moisture regimes in June, August, and September, as well as a very hot and dry period in July (Figure 1). During the winter season of 2021–2022, frosts alternated with periods of thawing, making such conditions generally unfavorable for plant overwintering. The regrowth of plants was delayed by the long, dry, and cool April of 2022.
2.6. Morphological Characterisation
The number of shoots formed was counted, and callus size was assessed in scores 1–3, depending on their diameter (0.5–1 cm; 1.1–2.0 cm, >2 cm, respectively), after 30 days of micropropagation. In the final passage before rooting, an experiment was carried out to determine the quality of the shoots in terms of shoot size. Shoots sized 10–15 mm were considered too small to be subjected to rooting. Larger shoots were considered optimal for transfer to rooting media.
Before transfer to ex vitro conditions, the number of roots per plantlet was counted, and root development was visually scored from 1 to 3 (1 for 1–2 cm long roots; 2 for 2–4 cm long roots; and 3 for >4 cm long roots) (Figure 2).
The survival rate (%) was calculated by dividing the number of alive plants by the total number of plants and multiplying by 100. The number of mature leaves on each plantlet was counted 21 days after transfer ex vitro.
The plantlet survival rate (%) in field conditions was calculated by dividing the number of alive plantlets after full vegetation recovery (June 2022) by the number of alive plantlets in the autumn of the planting year (October 2021) and multiplied by 100.
2.7. Statistical Analysis
The data were subjected to one-way, two-way, and three-way analysis of variance (ANOVA), followed by Tukey’s test (p < 0.05).
One-way ANOVA was used to analyze:
- -
- the impact of the disinfection on the seed infection rate;
- -
- the impact of the seed origin on the seed germination rate;
- -
- the impact of mericlone on the root development of in vitro plantlets;
- -
- impact of mericlone, substrate, or artificial light quality on the survival and number of leaves of E. maritimum ex vitro plantlets 21 days after ex vitro transplantation.
Two-way ANOVA was used to assess:
- -
- The impact of in vitro media macrosalt content and concentration of indole-3-acetic on the number of roots formed;
- -
- the impact of pot size on the survival and number of leaves of E. maritimum ex vitro plantlets 21 days after transplantation;
- -
- the impact of mericlone and cultivation length on the field survival rate;
- -
- the impact of planting date and cultivation length on the field survival rate.
Three-way ANOVA was used to determine:
- -
- The impact of basal media composition and supplementation of media with various cytokinins and the concentration of the cytokinins on the number of regenerated axillary shoots, the number of optimal sizes (>16 mm) and small (10–15 mm) axillary shoots, and shoot basal callus size;
All analyses were conducted using R version 4.0.4.1.
3. Results
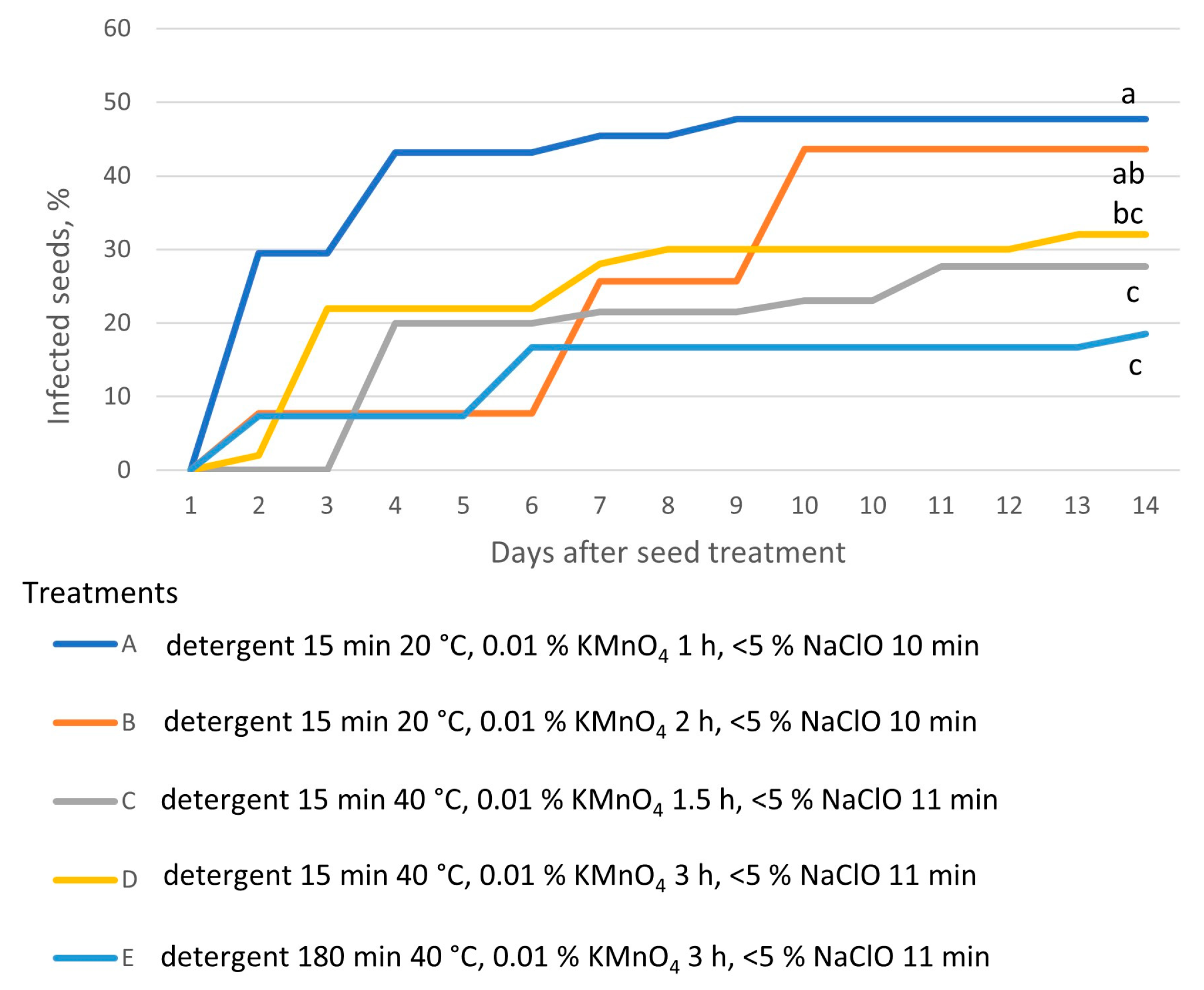
All of the multistep disinfection methods that were tested, which included putting the seeds in diluted detergent, a KMnO4 solution, and commercial bleach, eliminated the infection (Figure 3). The treatment protocol had a significant effect on the final percentage of infected seeds (Table S2). An iterative approach to the experimental design was taken. In each iteration, the length of immersion at one or two compounds was increased, and additionally, in iterations C–D (Table 2), the temperature of household detergent was increased from 20 °C to 40 °C. The first two iterations used the same amount of detergent and bleach but differed in the amount of KMnO4 solution applied (1 and 2 h, respectively). Both treatments resulted in insufficient elimination of contamination (>40% infected seeds). In the next iteration, the temperature of the detergent solution was raised by 20 °C, and bleach was applied for one extra minute. The combined effect of those two changes significantly reduced the infection rate (<30%), according to Tukey’s test (Figure 4). The last iteration, which combined prolonged application of KMnO4 and bleach, and a warm detergent solution, resulted in the largest proportion of contamination-free explants (81.5%). Contamination of explants appeared up to 14 days after the initiation of plant tissue culture (Figure 4).
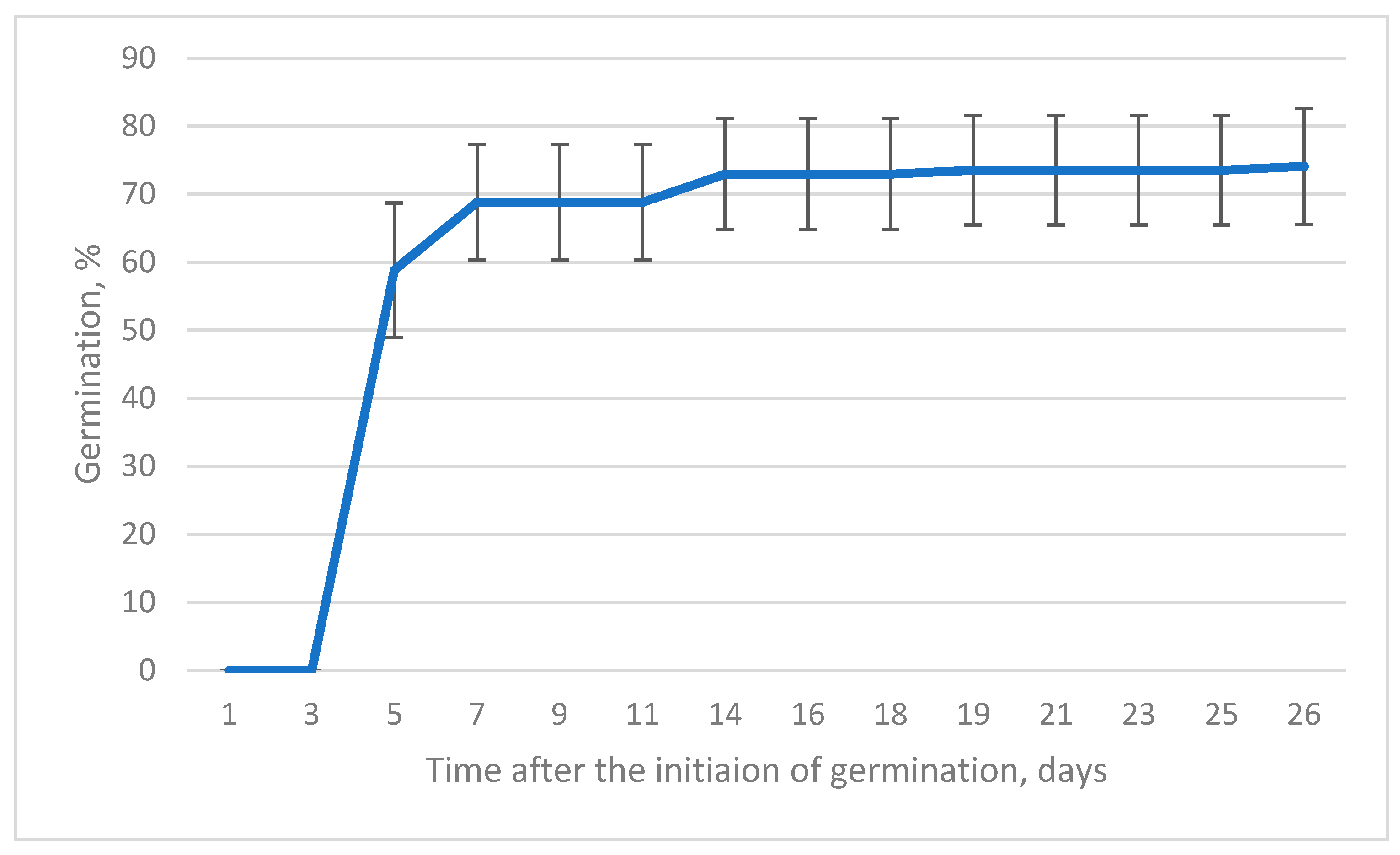
Seeds completed germination within 26 days after transfer from the cold stratification (6 months) to 22 °C in the growing chamber. After such a prolonged stratification, germination was very vigorous, and most of the sprouting seeds (79%) completed germination within the first five days (Figure 4).
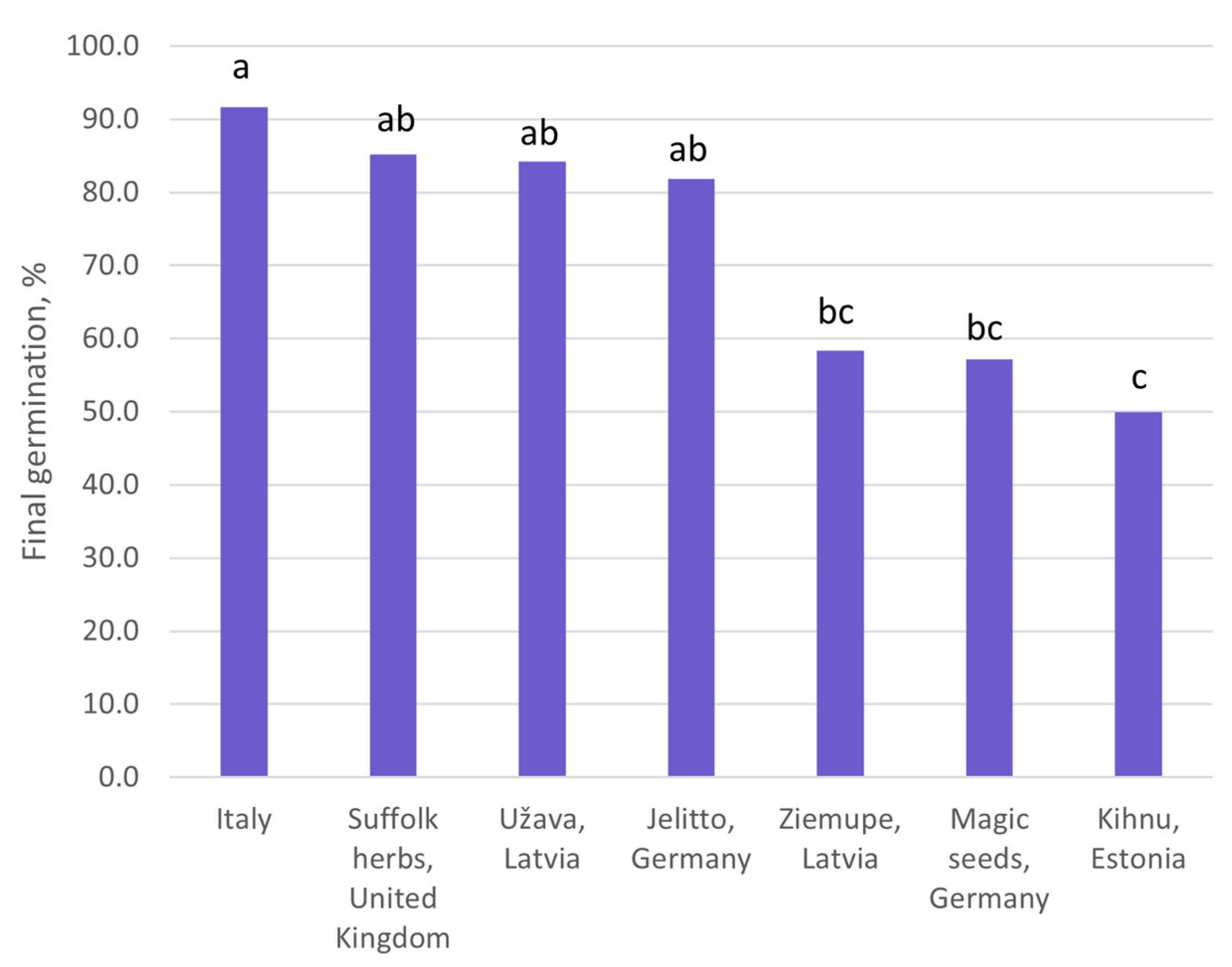
The final germination rate was significantly affected by seed origin (Table S3). The highest germination rate was observed in the population from the Mediterranean region, Italy (97.1%). The germination rate of Atlantic populations did not correspond to the longitudinal gradient of origin. Three Atlantic populations had germination rates statistically indifferent from the Mediterranean one (>81.8%) (Figure 5), but three populations had significantly lower germination rates (<60%) according to Tukey’s test. All the seeds continued to grow as vital, healthy plantlets.
3.1. Micropropagation
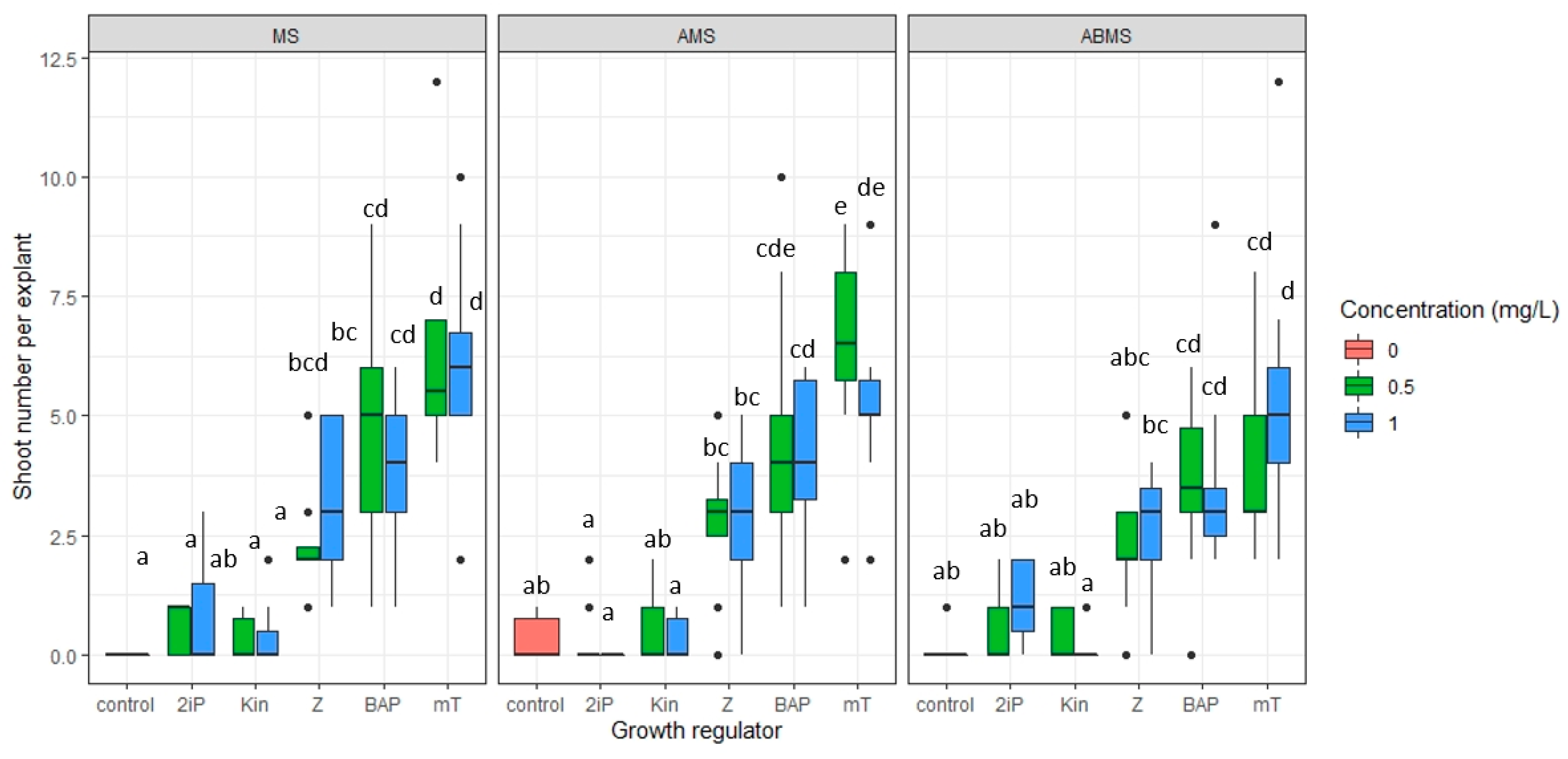
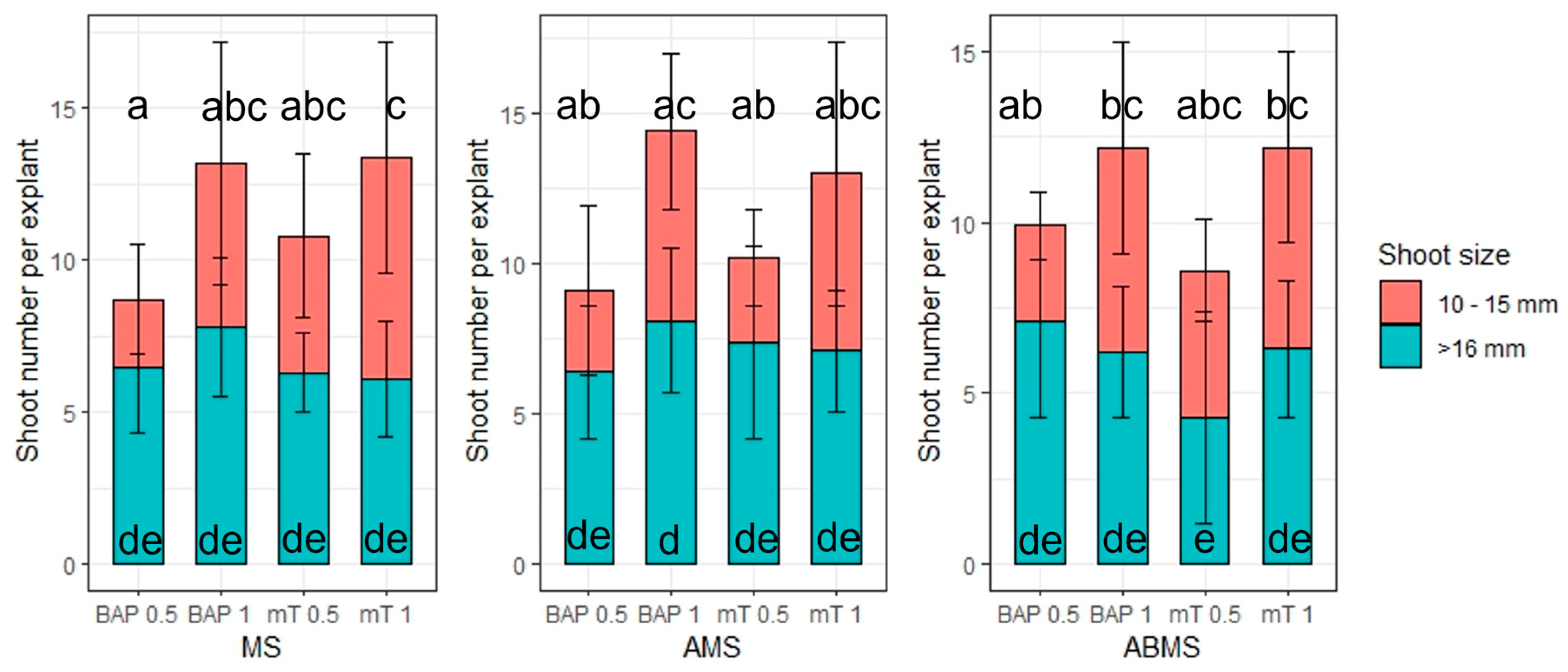
The number of shoots formed at the first passage of multiplication was significantly influenced by the type of plant growth regulator (PGR) added to the media (p ≤ 0.001) and the types of media used (p ≤ 0.05) (Table S4). The addition of 2 iP and Kin up to 1 mg L−1 did not significantly increase the number of shoots per explant (Figure 6). Supplementation of media with Z, BAP and mT stimulated axillary bud proliferation equally well at both 0.5 and 1.0 mg L−1. The largest number of shoots formed per explant (≥6.1) was observed on MS media with mT at 0.5 and 1.0 mg L−1 and AMS with mT at 0.5 mg L−1 (Figure 6). Even though the average number of shoots was smaller on BAP-supplemented media compared to mT, the differences were not statistically significant. Therefore, the effect of supplementing media with BAP or mT on shoot quality was determined in terms of the number of small (10–15 mm) and optimal (>16 mm) shoots that formed (Figure 7). The formation of sufficiently large shoots is crucial at the rooting stage. Medium composition (MS, AMS, ABMS) and supplemented cytokinin (mT, BAP) affected the number of optimal shoots formed (Table S5). A post hoc Tukey test revealed that the average number of optimal-size axillary shoots is statistically indifferent among most of the treatments. The only significant differences were between the two variants (AMS with BAP at 1 mg L−1 and AMBS with mT at 0.5 mg L−1). The type of cytokinin used and its concentration had a significant effect (p ≤ 0.05) on the number of small shoots that formed (Table S6). At lower cytokinin concentration (0.5 mg L−1), fewer small shoots were formed compared to 1.0 mg L−1.
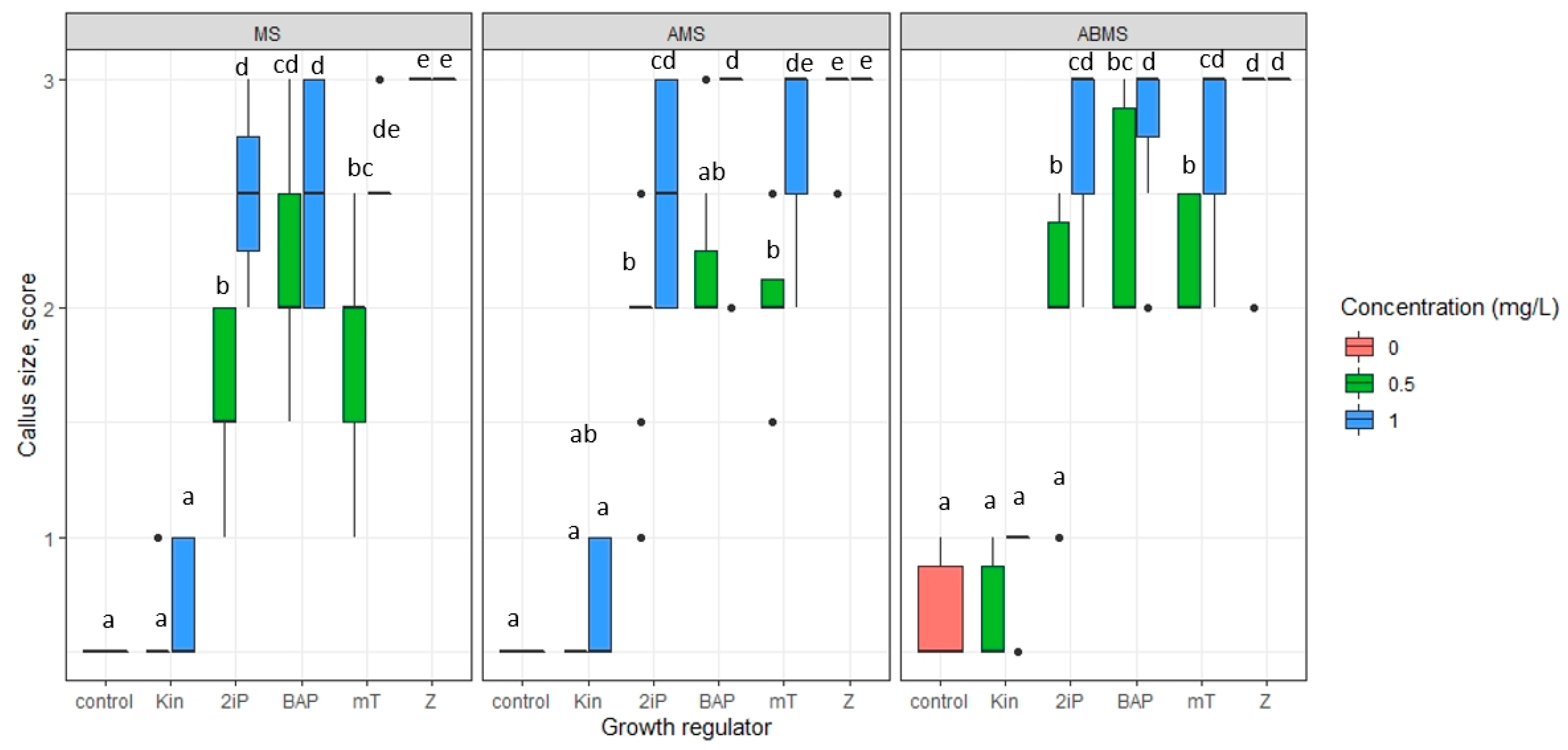
All regenerants developed a callus at the base of their shoots. The size of the callus was significantly (p ≤ 0.001) (Table S7) affected by the applied PGR (BAP mT, 2iP, Z, Kin), their concentration, the medium, and the interaction between the PGR and the medium (Figure 8). the increase of 2iP, BAT and mT concentration from 0.5 to 1.0 mg L−1 resulted in a larger average callus ball size. The application of Z yielded the largest callus balls (average score, 3) at both applied concentrations, but the application of Kin produced small calli (Figure 8). Among the tested variants, MS medium supplemented with 0.5 mg L−1 mT was the most successful for micropropagation of E. maritimum through axillary bud cultures, as the propagation rate was high and the size of the calli was smaller than that in other high-propagation-rate variants.
3.2. Rooting
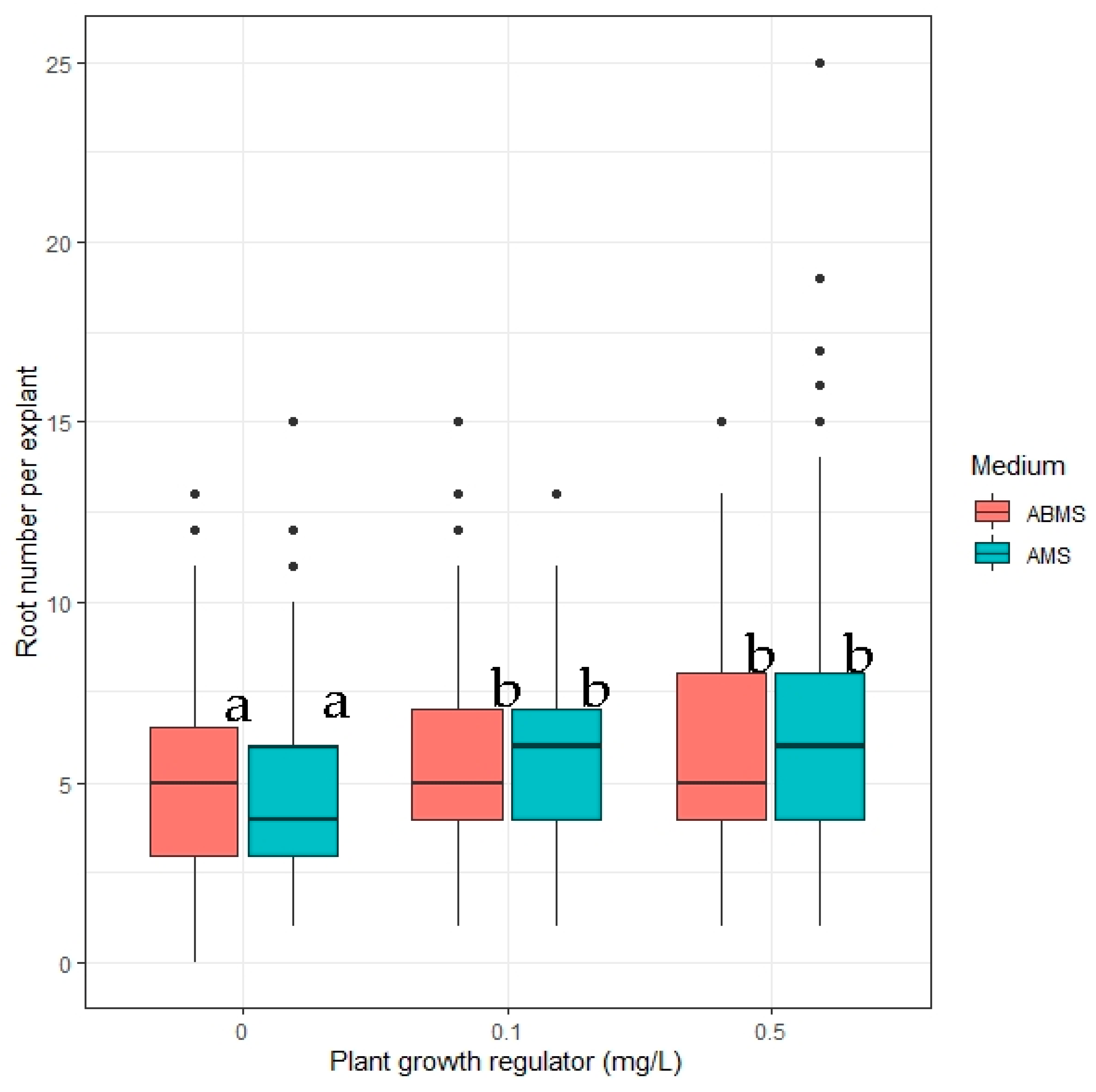
Spontaneous rooting was observed on all propagation media. The experimental media for rooting initiation were AMS and ABMS with three different IAA concentrations: 0, 0.1, and 0.5 mg L−1. All tested media resulted in 100% rooting. The addition of plant growth regulator significantly (p ≤ 0.05) increased the number of roots formed (Table S8), but there were no significant differences (p > 0.05) between the two media compositions or the concentration of IAA (0.1 versus 0.5 mg L−1) (Figure 9).
3.3. Ex Vitro Adaptation and Transfer to Field Conditions
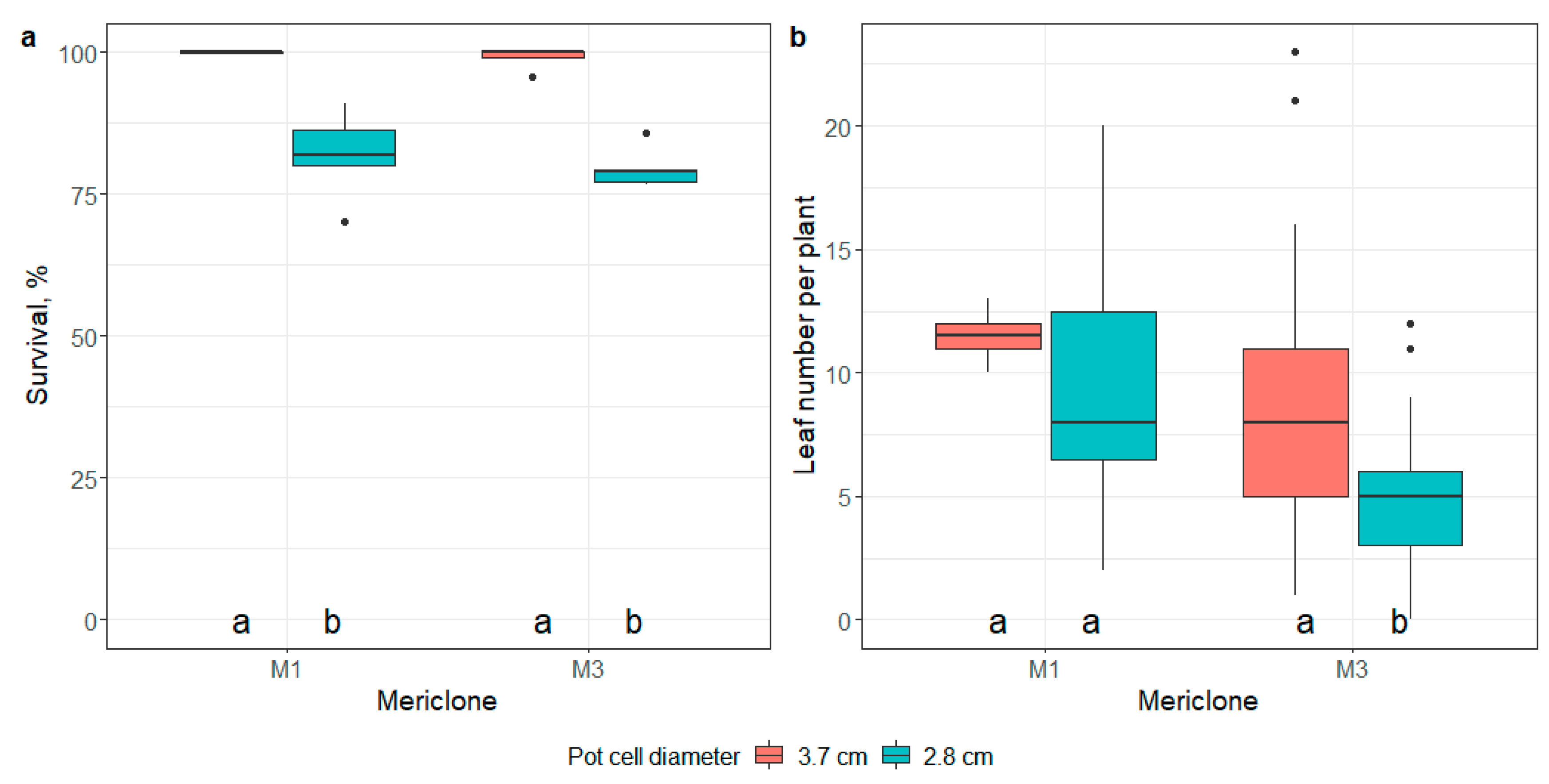
There were significant differences in root development (p < 0.05) among the mericlones of the accession collected at Užava, Latvia, at the ex vitro transplantation stage (Table S9). The root development of the M3 mericlone scored 1.8 on average on a scale of 1–3, but the M1 and M2 mericlones scored 1.3 (Table 3). Ex vitro adaptation resulted in a survival rate of over 99% for all tested mericlones (Table 3 and Table S10).
The average number of leaves per plantlet (5.7 to 6.5) 21 days after transplantation did not significantly differ among the mericlones (Table 3 and Table S11).
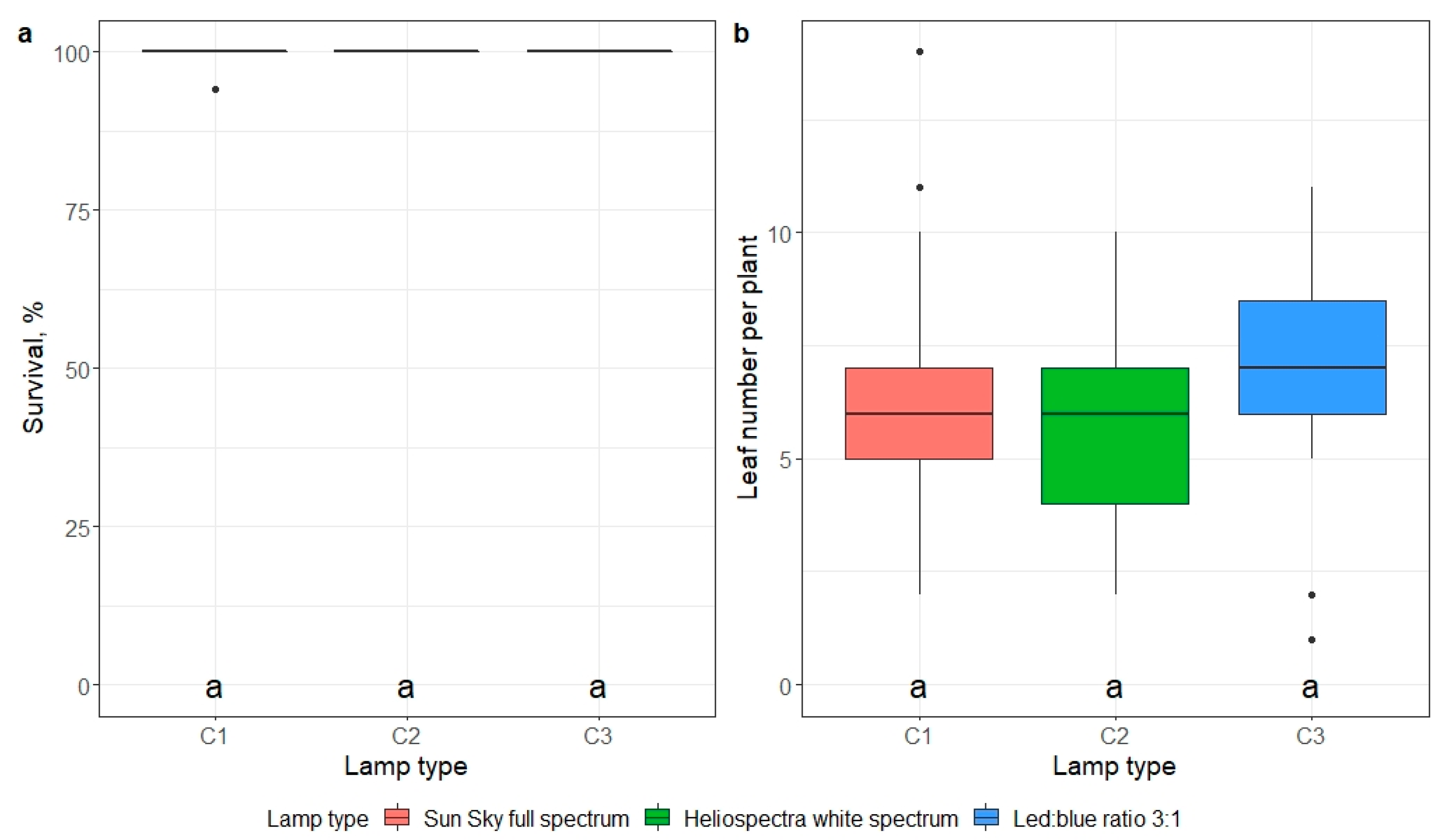
Artificial light quality (full spectrum LED lights versus blue and red emitting LEDs) did not affect plantlet survival or the number of leaves per plantlet during 21 days of cultivation after transfer ex vitro (Tables S12 and S13). Plant survival exceeded 99% under all tested LED lights (Figure 10a). Under two different full-spectrum LEDs, the average number of leaves was 6.0 and 5.6, respectively, and 6.6 under blue and red LEDs (Figure 10b).
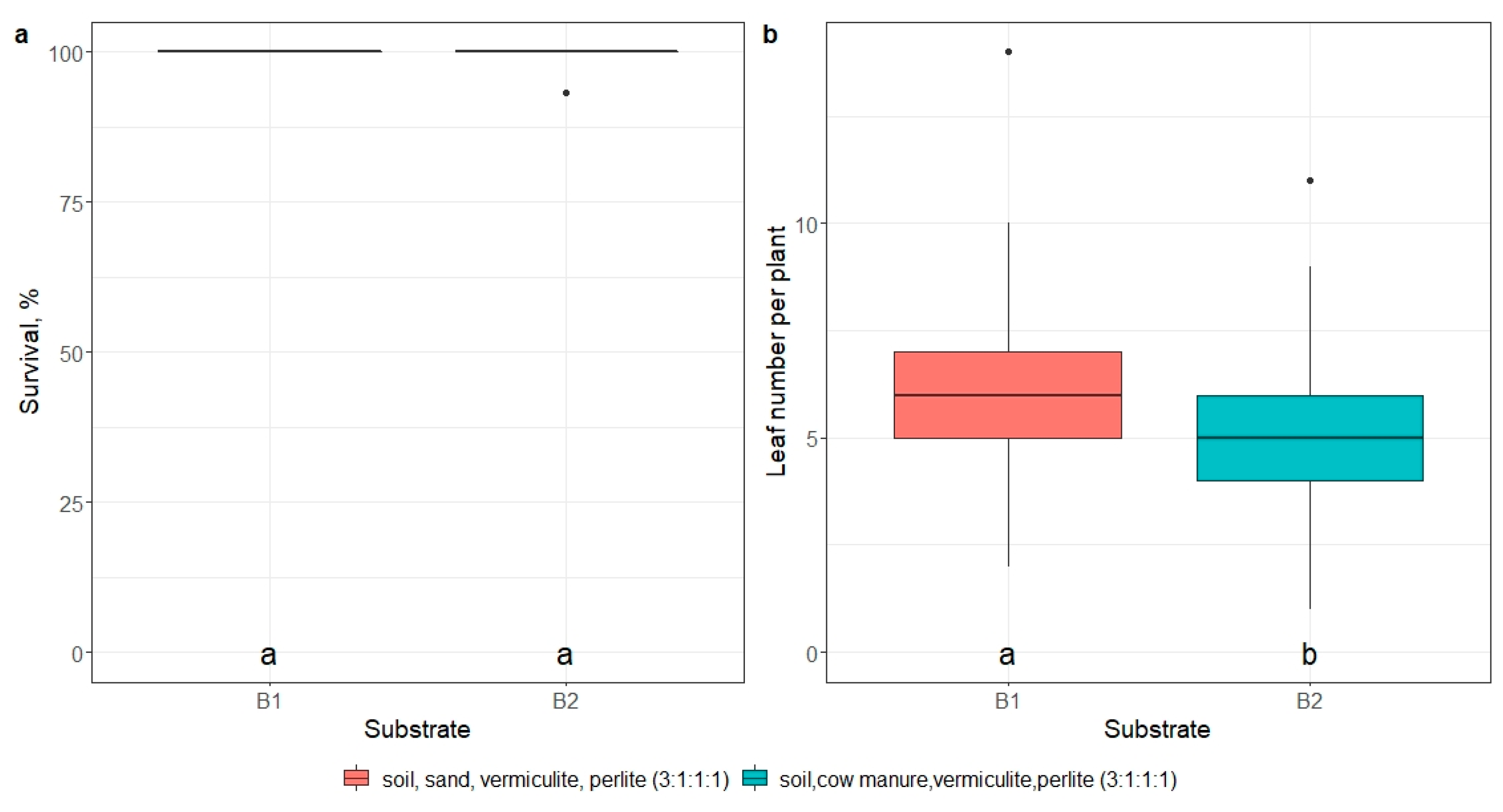
The potting mixture composition affected plantlet growth. The addition of cow manure instead of sand in the potting mixture significantly (p < 0.01) reduced the number of leaves (5.0 and 5.9 leaves per plantlet, respectively) but did not affect plantlet survival (p > 0.05) (Figure 11, Tables S14 and S15).
The survival of plantlets and the average number of leaves per plant significantly increased (p < 0.01) when the cell diameter of the multicell propagation trays was increased from 2.8 to 3.7 cm (Figure 12, Tables S16 and S17).
Plantlet survival in the open field decreased significantly with the later planting date (p < 0.05), without notable differences between tested mericlones (p > 0.05) (Tables S18 and S19). More than 86% of plants planted in June survived to the autumn, while for the plants planted in July, the number decreased to 55% (Table 4). Survival decreased by an additional 20–30% after the winter for trials planted in June and July, respectively.
4. Discussion
While the wild populations of many medicinal plants face extinction due to habitat destruction and overexploitation, the demand for medicinal plants is ever-increasing [65]. In general, propagation from seed is the most convenient and cost-effective method for cultivation, but this is not true for E. maritimum due to the legal prohibition against seed collection in the wild [15], seed dormancy due to undeveloped embryos, and erratic germination [13]. In vitro propagation allows a large number of plantlets to be obtained from a small quantity of source material, thus providing the opportunity to create large ex situ collections or cultivate plants on a commercial scale. The present study focuses on the development of a protocol for micropropagation starting from in vitro-germinated seeds and for ex vitro and field adaptation of E. maritimum plantlets.
A previous study on E. maritimum established in vitro cultures from apical and axillary buds but did not succeed in initiating in vitro cultures from seeds, despite many attempts [7]. In this study, we developed a method for plant tissue culture establishment from seeds that proved successful for seeds of different origins. The high infection rate of in vitro germinated seeds is one of the main problems with establishing plant tissue culture in the genus Eryngium. Because of this, we tried a method that has been used successfully with E. viviparum [10] and E. planum [18]. It involves the application of several disinfectants in consecutive order. The best protocol resulted in 81.5% infection-free explants. It consisted of seed incubation for 180 min in diluted detergent at 40 °C, followed by 3 h incubation in 0.01% KMnO4 and 11 min of sterilization in commercial bleach. The different agents have complementary modes of action. Detergents promote even distribution of disinfectants on tissues [30], and the strong oxidants with broad-spectrum antibacterial and antifungal effects [28,29] proved to be successful at eliminating contamination. In addition, seed treatment with warm water and KMnO4 has been demonstrated to enhance germination [26,34]. For the abovementioned reasons, seed treatment with warm water has increasingly been used in organic agriculture [26]. The temperature also affects the effectiveness of the active substance [66], in this case, the detergent.
E. maritimum seeds are not completely developed at the end of the fruiting period and exhibit morphophysiological dormancy [13]. Cold stratification and the use of GA3 have both been shown to break dormancy [13]. Significant differences in stratification response, viability, and germination have been observed between populations at locations with different climatic conditions; seeds from northern populations of the Atlantic coast of Europe require a longer cold stratification period compared to seeds from the Mediterranean region [67]. In the current study, the cold stratification period (30 weeks) was longer than in other studies, which applied a maximum of 14 weeks for Mediterranean populations and 20 weeks for Atlantic populations [67]. Cortés-Fernández et al. [67] pointed out that seeds of populations from higher latitudes need longer cold stratification periods to release them from dormancy. In this study, prolonged cold stratification was applied to populations both from the Mediterranean and Atlantic regions. If the lower germination rates in Atlantic populations were due to more pronounced dormancy, the longer pretreatment should balance out the germination outcomes. The remaining difference in germination confirms the previously published hypothesis that a necessity for prolonged stratification is not the only factor influencing germination and that other factors, such as poor pollination or genetic inbreeding, play a role [67]. Additionally, the possibility to successfully ripen seeds differs in Mediterranean and Atlantic regions; e.g., in Spain, fruits ripen in August [68], while in the Baltic Sea region, fruit ripening takes place until October [69], when frost and snow cover are not unusual [69], and the possibility for seeds to complete maturation is inhibited.
The micropropagation protocol presented in this study can be implemented for commercial and conservational purposes of E. maritimum. A hormone-free medium can be used for seed germination but not for efficient propagation. Cytokinin application and ammonium nitrate reduction in the media helped to achieve a high propagation rate. The application of mT and IAA resulted in a larger propagation rate than previously reported in E. maritimum on media supplemented with BAP and IAA [7]. In contrast to this study, no calluses formed during the micropropagation in the previous report [7]. Calli challenge micropropagation and shoot regeneration from calli induce somaclonal variation [70]. In this study, the calli could have formed because of genetic differences between plant populations or because different PGRs or their rates were used compared to the previous study [7].
The ability to root in vitro propagated shoots is critical for acclimating plants to ex vitro and field conditions. It is well known that reduced salt concentrations and the addition of auxins to the basal medium result in better root formation [38]. Basal media with reduced nitrogen content supported good root formation in this study. Modified media are often used at the rooting stage in plant tissue cultures as decreased salt concentration decreases osmotic pressure, which in turn stimulates rooting [71]. Rooting was observed on PGR—free media, indicating the presence of endogenous auxins. Exogenous auxin application increased adventitious rooting. The results are in agreement with Thiem’s [18] finding in E. planum that auxin application increases root number. Further experiments determining the combined effect of different auxins are recommended, as they have been proven to induce a larger number of roots than single auxins in E. alpinum [43]. The quality of the root system of in vitro plantlets influences ex vitro acclimatization. This study confirmed that in vitro plantlets with a good root system have a higher survival rate and faster growth during the ex vitro adaptation stage.
Acclimatization and transplantation of plantlets from in vitro to ex vitro conditions have been identified as the most challenging stage of in vitro propagation [14]. Many factors affect survival, e.g., air and soil moisture and temperature, photoperiod, light intensity and spectrum, and physiochemical properties of the potting substrate. Pot type and size can influence the growth and survival of plantlets [72]. In commercial nurseries, pot size must be chosen based on a compromise between production costs and plant quality. Expenses can be reduced by using small pots, thus growing more plants per 1 m2. Larger volume pots encourage the growth of shoots and roots [73]. We observed that the reduction of pot cell diameter resulted in a decrease in the plantlet survival rate and the number of leaves per plant.
Different soil substrate mixtures can change the conditions provided for plantlets’ adaptation to ex vitro conditions. Locally available compost based on cow manure can be used as a nutrient-rich [56] peat substitute in substrate mixtures. The addition of nutrient sources to potting mix can increase ex vitro adaptation success [47,57]. In this study, adding cow manure to the potting mixture did not change survival but reduced leaf growth. Sea holly has adapted to grow in dunes with limited nutrient and water availability [74]. The lower availability of nutrients in substrate mixtures without cow manure probably promotes better root development because less nutrient-rich substrates have smaller osmotic pressure, which stimulates rhizogenesis [71]. Even though nitrogen-rich substrates promote plant aboveground growth, root growth is stimulated by less nutritional soils [75]. As roots formed during growth in plant tissue cultures are underdeveloped [76], conditions promoting rhizogenesis are preferable to sustain plant growth.
Optimal lighting is critical during the ex vitro adaptation stage [53]. Two commercially available full spectrum LEDs and one blue/red LED ensured equally high survival rates and equally good leaf growth within 21 days of ex vitro transfer. Cabral et al. [72] also found no influence of full spectrum versus blue/red LEDs on the survival of ex vitro plantlets. However, LED spectral quality during ex vitro adaptation in Rubus idaeus [53] and Epidendrum denticulatum [72] influenced morphophysiological parameters. The current study focused only on plant leaf growth as an easily measurable indicator of plant growth; therefore, the results must be considered with caution.
In field conditions, crops typically face various environmental stress factors. Plant adaptation to field conditions, and thus productivity, is strongly influenced by soil characteristics, air and soil temperature, and soil moisture. Planting time must be adjusted based on the local climatic conditions and the species of interest [77]. The growing conditions in Northern Europe are characterized by a short growing season and cold winters [78]; therefore, good plantlet development prior to overwintering is critical. For E. maritimum, the first planting date in June ensured a good survival rate (at the end of the vegetation season and after overwintering. The higher plantlet survival observed with an early planting date in June is most likely due to higher soil moisture, optimal temperature conditions, and a longer period for plant development until the end of vegetation. To the best of our knowledge, this is the first report on the planting time of sea holly in Northern European conditions, and the findings show that planting at least four months before the end of the vegetation season is required for effective plantlet adaptation.
5. Conclusions
In conclusion, sea holly seed disinfection results in a low contamination rate if a series of disinfectants with a long application time is used. Prolonged cold storage of seeds in sterile culture results in a high germination rate and speed, as well as vigorous seedlings. The micropropagation and rooting efficiency can be enhanced by adding plant growth regulators and reducing the nitrogen content in the media. A substrate with low nitrogen content and a larger pot size stimulates growth and increases plantlet survival during the ex vitro adaptation stage. Planting for at least four months before the end of vegetation season in the field conditions in Northern Europe is necessary to achieve a high survival rate during overwintering.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/horticulturae9020271/s1, Table S1: Composition of Murashige—Skoog’s medium (MS), and its modifications AMS and ABMS media; Table S2: Analysis of variance of the seed infection rate during in vitro establishment, as influenced by disinfection treatment; Table S3: Analysis of variance of the origin on Eryngium maritimum in vitro germination rate; Table S4: Analysis of variance of the impact of basal media composition and supplementation of media with various cytokinins and the concentration of the cytokinins on the number of regenerated axillary shoots; Table S5: Analysis of variance of the impact of basal media composition and supplementation of media with various cytokinins and the concentration of the cytokinins on the number of >16 mm long regenerated axillary shoots; Table S6: Analysis of variance of the impact of basal media composition and supplementation of media with various cytokinins and the concentration of the cytokinins on the number of 10–15 mm long regenerated axillary shoots; Table S7: Analysis of variance of the impact of basal media composition and supplementation of media with various cytokinins and the concentration of the cytokinins on the shoot basal callus size; Table S8: Analysis of variance of the impact of media macrosalt content and concentration of indole-3-acetic on number of roots formed; Table S9: Analysis of variance of the impact of mericlone on the root development of E. maritimum in vitro plantlets; Table S10: Analysis of variance of the impact of mericlone on the survival of E. maritimum in vitro plantlets 21 days after ex vitro transplantation; Table S11: Analysis of variance of the impact of mericlone on the number of leaves of E. maritimum in vitro plantlets 21 days after ex vitro transplantation; Table S12: Analysis of variance of the impact of artificial light quality on the number of leaves of E. maritimum ex vitro plantlets 21 days after ex vitro transplantation; Table S13: Analysis of variance of the impact of artificial light quality on the survival of E. maritimum ex vitro plantlets 21 days after ex vitro transplantation; Table S14: Analysis of variance of the impact of substrate on the number of leaves of the mericlone M2 plantlets 21 days after ex vitro transplantation; Table S15: Analysis of variance of the impact of substrate on the plantlet survival of the mericlone M2 21 days after ex vitro transplantation; Table S16: Analysis of variance of the impact of pot cell diameter on the survival of E. maritimum ex vitro plantlets 21 days after transplantation; Table S17: Analysis of variance of the impact of pot cell diameter on the number of leaves of E. maritimum ex vitro plantlets 21 days after transplantation; Table S18: Analysis of variance of the impact of planting date and cultivation length on field survival rate; Table S19: Analysis of variance of the impact of mericlone and cultivation length on field survival rate.
Author Contributions
Conceptualization, I.M.; methodology, I.M., D.K. and A.K.; investigation, L.K. and D.K.; data curation, L.K.; writing—original draft preparation, I.M.; writing—review and editing, D.K., L.K. and A.K.; visualization, I.M. and A.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the European Regional Development Fund, grant number 1.1.1.1/19/A/083, “Research on plant tissue culture application for commercial propagation of endangered medicinal plants”.
Data Availability Statement
I.M. is responsible for data-keeping. Data are available upon request.
Acknowledgments
The authors thank Dott. Marco D’Antraccol from the University of Pisa Botanical Garden for seed material.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Clausing, G.; Vickers, K.; Kadereit, J.W. Historical Biogeography in a Linear System: Genetic Variation of Sea Rocket (Cakile Maritima) and Sea Holly (Eryngium Maritimum) along European Coasts. Mol. Ecol. 2000, 9, 1823–1833. [Google Scholar] [CrossRef] [PubMed]
- Ben Lajnef, H.; Ferioli, F.; Pasini, F.; Politowicz, J.; Khaldi, A.; Filippo D’Antuono, L.; Caboni, M.F.; Nasri, N. Chemical Composition and Antioxidant Activity of the Volatile Fraction Extracted from Air-Dried Fruits of Tunisian Eryngium Maritimum L. Ecotypes. J. Sci. Food Agric. 2018, 98, 635–643. [Google Scholar] [CrossRef] [PubMed]
- Darriet, F.; Andreani, S.; De Cian, M.C.; Costa, J.; Muselli, A. Chemical Variability and Antioxidant Activity of Eryngium Maritimum L. Essential Oils from Corsica and Sardinia. Flavour Fragr. J. 2014, 29, 3–13. [Google Scholar] [CrossRef]
- Darriet, F.; Znini, M.; Majidi, L.; Muselli, A.; Hammouti, B.; Bouyanzer, A.; Costa, J. Evaluation of Eryngium Maritimum Essential Oil as Environmentally Friendly Corrosion Inhibitor for Mild Steel in Hydrochloric Acid Solution. Int. J. Electrochem. Sci 2013, 8, 4328–4345. [Google Scholar]
- Kikowska, M.; Kalemba, D.; Dlugaszewska, J.; Thiem, B. Chemical Composition of Essential Oils from Rare and Endangered Species—Eryngium maritimum L. and E. alpinum L. Plants 2020, 9, 417. [Google Scholar] [CrossRef] [Green Version]
- Maggio, A.; Bruno, M.; Formisano, C.; Rigano, D.; Senatore, F. Chemical Composition of the Essential Oils of Three Species of Apiaceae Growing Wild in Sicily: Bonannia Graeca, Eryngium Maritimum and Opopanax Chironium. Nat. Prod. Commun. 2013, 8, 1934578X1300800640. [Google Scholar] [CrossRef] [Green Version]
- Kikowska, M.; Thiem, B.; Sliwinska, E.; Rewers, M.; Kowalczyk, M.; Stochmal, A.; Oleszek, W. The Effect of Nutritional Factors and Plant Growth Regulators on Micropropagation and Production of Phenolic Acids and Saponins from Plantlets and Adventitious Root Cultures of Eryngium maritimum L. J. Plant Growth Regul. 2014, 33, 809–819. [Google Scholar] [CrossRef] [Green Version]
- Kikowska, M.; Chanaj-Kaczmarek, J.; Derda, M.; Budzianowska, A.; Thiem, B.; Ekiert, H.; Szopa, A. The Evaluation of Phenolic Acids and Flavonoids Content and Antiprotozoal Activity of Eryngium Species Biomass Produced by Biotechnological Methods. Molecules 2022, 27, 363. [Google Scholar] [CrossRef]
- Sultana, K.W.; Das, S.; Chandra, I.; Roy, A. Efficient Micropropagation of Thunbergia Coccinea Wall. and Genetic Homogeneity Assessment through RAPD and ISSR Markers. Sci. Rep. 2022, 12, 1683. [Google Scholar] [CrossRef]
- Ayuso, M.; García-Pérez, P.; Ramil-Rego, P.; Gallego, P.P.; Barreal, M.E. In Vitro Culture of the Endangered Plant Eryngium Viviparum as Dual Strategy for Its Ex Situ Conservation and Source of Bioactive Compounds. Plant Cell Tissue Organ Cult. 2019, 138, 427–435. [Google Scholar] [CrossRef]
- Pence, V.C.; Meyer, A.; Linsky, J.; Gratzfeld, J.; Pritchard, H.W.; Westwood, M.; Bruns, E.B. Defining Exceptional Species—A Conceptual Framework to Expand and Advance Ex Situ Conservation of Plant Diversity beyond Conventional Seed Banking. Biol. Conserv. 2022, 266, 109440. [Google Scholar] [CrossRef]
- Pence, V.C.; Ballesteros, D.; Walters, C.; Reed, B.M.; Philpott, M.; Dixon, K.W.; Pritchard, H.W.; Culley, T.M.; Vanhove, A.-C. Cryobiotechnologies: Tools for Expanding Long-Term Ex Situ Conservation to All Plant Species. Biol. Conserv. 2020, 250, 108736. [Google Scholar] [CrossRef]
- Necajeva, J.; Ievinsh, G. Seed Dormancy and Germination of an Endangered Coastal Plant Eryngium Maritimum (Apiaceae). Est. J. Ecol. 2013, 62, 150–161. [Google Scholar] [CrossRef] [Green Version]
- Bączek, K.; Pawełczak, A.; Przybył, J.L.; Kosakowska, O.; Węglarz, Z. Secondary Metabolites of Various Eleuthero (Eleutherococcus Senticosus/Rupr. et Maxim./Maxim) Organs Derived from Plants Obtained by Somatic Embryogenesis. In Plant Cell and Tissue Differentiation and Secondary Metabolites: Fundamentals and Applications; Ramawat, K.G., Ekiert, H.M., Goyal, S., Eds.; Springer International Publishing: Cham, Switzerland, 2021; pp. 433–466. ISBN 978-3-030-30185-9. [Google Scholar]
- Kikowska, M.; Thiem, B. In Vitro Systems of Selected Eryngium Species (E. Planum, E. Campestre, E. Maritimum, and E. Alpinum) for Studying Production of Desired Secondary Metabolites (Phenolic Acids, Flavonoids, Triterpenoid Saponins, and Essential Oil). In Plant Cell and Tissue Differentiation and Secondary Metabolites: Fundamentals and Applications; Ramawat, K.G., Ekiert, H.M., Goyal, S., Eds.; Springer International Publishing: Cham, Switzerland, 2021; pp. 869–901. ISBN 978-3-030-30185-9. [Google Scholar]
- Kikowska, M.; Kruszka, D.; Derda, M.; Hadaś, E.; Thiem, B. Phytochemical Screening and Acanthamoebic Activity of Shoots from in Vitro Cultures and in Vivo Plants of Eryngium Alpinum L.—The Endangered and Protected Species. Molecules 2020, 25, 1416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kikowska, M.; Thiem, B.; Sliwinska, E.; Rewers, M.; Kowalczyk, M.; Stochmal, A.; Długaszewska, J. Micropropagation of Eryngium Campestre L. Via Shoot Culture Provides Valuable Uniform Plant Material with Enhanced Content of Phenolic Acids and Antimicrobial Activity. Acta Biol. Cracoviensia Ser. Bot. 2016, 58, 43–56. [Google Scholar] [CrossRef]
- Thiem, B.; Kikowska, M.; Krawczyk, A.; Więckowska, B.; Sliwinska, E. Phenolic Acid and DNA Contents of Micropropagated Eryngium planum L. Plant Cell, Tissue Organ Cult. 2013, 114, 197–206. [Google Scholar] [CrossRef] [Green Version]
- Kļaviņa, D.; Gailīte, A.; Ievinsh, G. Initial Responses of Explants from Rare and Endangered Coastal Plant Species during Initiation of Tissue Culture. Acta Univ. Latv. 2006, 710, 81–91. [Google Scholar]
- Nagananda, G.S.; Rajath, S.; Mathew, R.K.; Rajan, S.S. Effect of Adjuvants and Nitrogen Sources on in Vitro Shoot Regeneration and Clonal Propagation of Medicinally Important Plant Eryngium foetidum L. Res. Biotechnol. 2012, 3, 21–25. [Google Scholar]
- Kikowska, M.; Thiem, B.; Szopa, A.; Klimek-Szczykutowicz, M.; Rewers, M.; Sliwinska, E.; Ekiert, H. Comparative Analysis of Phenolic Acids and Flavonoids in Shoot Cultures of Eryngium Alpinum L.: An Endangered and Protected Species with Medicinal Value. Plant Cell, Tissue Organ Cult. 2019, 139, 167–175. [Google Scholar] [CrossRef] [Green Version]
- Fay, M.F. Conservation of Rare and Endangered Plants Using in Vitro Methods. Vitr. Cell. Dev. Biol. Plant 1992, 28, 1–4. [Google Scholar] [CrossRef]
- Benson, E.E.; Danaher, J.E.; Pimbley, I.M.; Anderson, C.T.; Wake, J.E.; Daley, S.; Adams, L.K. In Vitro Micropropagation of Primula Scotica: A Rare Scottish Plant. Biodivers. Conserv. 2000, 9, 711–726. [Google Scholar] [CrossRef]
- Bertsouklis, K.; Theodorou, P.; Aretaki, P.-E. In Vitro Propagation of the Mount Parnitha Endangered Species Sideritis Raeseri Subsp. Attica. Horticulturae 2022, 8, 1114. [Google Scholar] [CrossRef]
- Ezz, A.L.; Dura, S.; Daradkeh, N. Propagation Physiology of Juniperus Phoenicea L. from Jordan Using Seeds and in Vitro Culture Techniques: Baseline Information for a Conservation Perspective. African J. Biotechnol. 2012, 11, 7684–7692. [Google Scholar]
- Kim, M.; Shim, C.; Lee, J.; Wangchuk, C. Hot Water Treatment as Seed Disinfection Techniques for Organic and Eco-Friendly Environmental Agricultural Crop Cultivation. Agriculture 2022, 12, 1081. [Google Scholar] [CrossRef]
- Sen, M.K.; Jamal, M.; Nasrin, S. Sterilization Factors Affect Seed Germination and Proliferation of Achyranthes Aspera Cultured in Vitro. Environ. Exp. Biol. 2013, 11, 119–123. [Google Scholar]
- Gammoudi, N.; Nagaz, K.; Ferchichi, A. Establishment of Optimized in Vitro Disinfection Protocol of Pistacia Vera L. Explants Mediated a Computational Approach: Multilayer Perceptron–Multi−objective Genetic Algorithm. BMC Plant Biol. 2022, 22, 324. [Google Scholar] [CrossRef]
- Yildiz, M.; Fatih Ozcan, S.; T Kahramanogullari, C.; Tuna, E. The Effect of Sodium Hypochlorite Solutions on the Viability and in Vitro Regeneration Capacity of the Tissue. Nat. Prod. J. 2012, 2, 328–331. [Google Scholar] [CrossRef]
- Sahu, P.K.; Tilgam, J.; Mishra, S.; Hamid, S.; Gupta, A.; Verma, S.K.; Kharwar, R.N. Surface Sterilization for Isolation of Endophytes: Ensuring What (Not) to Grow. J. Basic Microbiol. 2022, 62, 647–668. [Google Scholar] [CrossRef]
- Chokheli, V.A.; Dmitriev, P.A.; Rajput, V.D.; Bakulin, S.D.; Azarov, A.S.; Varduni, T.V.; Stepanenko, V.V.; Tarigholizadeh, S.; Singh, R.K.; Verma, K.K.; et al. Recent Development in Micropropagation Techniques for Rare Plant Species. Plants 2020, 9, 1733. [Google Scholar] [CrossRef]
- Tiwari, A.K.; Tripathi, S.; Lal, M.; Mishra, S. Screening of Some Chemical Disinfectants for Media Sterilization During In Vitro Micropropagation of Sugarcane. Sugar Tech 2012, 14, 364–369. [Google Scholar] [CrossRef]
- Subramanya, S.H.; Pai, V.; Bairy, I.; Nayak, N.; Gokhale, S.; Sathian, B. Potassium Permanganate Cleansing Is an Effective Sanitary Method for the Reduction of Bacterial Bioload on Raw Coriandrum Sativum. BMC Res. Notes 2018, 11, 124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grauda, D.; Lapiņa, L.; Jansone, B.; Jansons, A.; Rashal, I. Recovering Genetic Resources of Some Legume Species of Latvian Origin by Plant Tissue Culture. Proc. Latv. Acad. Sci. Sect. B Nat. Exact Appl. Sci. 2013, 67, 224–228. [Google Scholar] [CrossRef] [Green Version]
- Chokheli, V.A.; Bakulin, S.D.; Ermolaeva, O.Y.; Kozlovsky, B.L.; Dmitriev, P.A.; Stepanenko, V.V.; Kornienko, I.V.; Bushkova, A.A.; Rajput, V.D.; Varduny, T.V. Investigation of Growth Factors and Mathematical Modeling of Nutrient Media for the Shoots Multiplication In Vitro of Rare Plants of the Rostov Region. Horticulturae 2023, 9, 60. [Google Scholar] [CrossRef]
- Phillips, G.C.; Garda, M. Plant Tissue Culture Media and Practices: An Overview. Vitr. Cell. Dev. Biol. Plant 2019, 55, 242–257. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Kadapatti, S.S.; Murthy, H.N. In Vitro Micropropagation of Andrographis Macrobotrys. J. Herbs. Spices Med. Plants 2022, 28, 89–98. [Google Scholar] [CrossRef]
- Asensio, E.; de Medinacelli Juan-Méndez, R.; Juan-Vicedo, J. In Vitro Propagation and Phytochemistry of Thymol-Producing Plants from a Horticultural Form of Thymus × Josephi-Angeli Mansanet & Aguil. (Lamiaceae). Horticulturae 2022, 8, 1188. [Google Scholar]
- Schaller, G.E.; Street, I.H.; Kieber, J.J. Cytokinin and the Cell Cycle. Curr. Opin. Plant Biol. 2014, 21, 7–15. [Google Scholar] [CrossRef]
- Müller, D.; Leyser, O. Auxin, Cytokinin and the Control of Shoot Branching. Ann. Bot. 2011, 107, 1203–1212. [Google Scholar] [CrossRef] [Green Version]
- Abdalla, N.; El-Ramady, H.; Seliem, M.K.; El-Mahrouk, M.E.; Taha, N.; Bayoumi, Y.; Shalaby, T.A.; Dobránszki, J. An Academic and Technical Overview on Plant Micropropagation Challenges. Horticulturae 2022, 8, 677. [Google Scholar] [CrossRef]
- Kikowska, M.; Sliwinska, E.; Thiem, B. Micropropagation and Production of Somatic Seeds for Short-Term Storage of the Endangered Species Eryngium alpinum L. Plants 2020, 9, 498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beatrice, P.; Chiatante, D.; Scippa, G.S.; Montagnoli, A. Photoreceptors’ Gene Expression of Arabidopsis Thaliana Grown with Biophilic LED-Sourced Lighting Systems. PLoS ONE 2022, 17, e0269868. [Google Scholar] [CrossRef] [PubMed]
- Paradiso, R.; Proietti, S. Light-Quality Manipulation to Control Plant Growth and Photomorphogenesis in Greenhouse Horticulture: The State of the Art and the Opportunities of Modern LED Systems. J. Plant Growth Regul. 2022, 41, 742–780. [Google Scholar] [CrossRef]
- Hwang, H.; An, S.; Lee, B.; Chun, C. Improvement of Growth and Morphology of Vegetable Seedlings with Supplemental Far-Red Enriched LED Lights in a Plant Factory. Horticulturae 2020, 6, 109. [Google Scholar] [CrossRef]
- Akimova, S.; Radzhabov, A.; Esaulko, A.; Samoshenkov, E.; Nechiporenko, I.; Kazakov, P.; Voskoboinikov, Y.; Matsneva, A.; Zubkov, A.; Aisanov, T. Improvement of Ex Vitro Growing Completion of Highbush Blueberry (Vaccinium Corymbosum L.) in Containers. Forests 2022, 13, 1550. [Google Scholar] [CrossRef]
- Chen, L.; Yang, Y.; Chen, F.; Zhao, J.; Wang, X.; Khan, A.T.; Yuegao, X. Effects of Red and Blue LEDs on in Vitro Growth and Microtuberization of Potato Single-Node Cuttings. Front. Agric. Sci. Eng. 2018, 5, 197–205. [Google Scholar]
- Dutta Gupta, S.; Jatothu, B. Fundamentals and Applications of Light-Emitting Diodes (LEDs) in in Vitro Plant Growth and Morphogenesis. Plant Biotechnol. Rep. 2013, 7, 211–220. [Google Scholar] [CrossRef]
- Randall, W.C.; Lopez, R.G. Comparison of Supplemental Lighting from High-Pressure Sodium Lamps and Light-Emitting Diodes during Bedding Plant Seedling Production. HortScience 2014, 49, 589–595. [Google Scholar] [CrossRef]
- Hung, C.D.; Hong, C.-H.; Kim, S.-K.; Lee, K.-H.; Park, J.-Y.; Nam, M.-W.; Choi, D.-H.; Lee, H.-I. LED Light for in Vitro and Ex Vitro Efficient Growth of Economically Important Highbush Blueberry (Vaccinium Corymbosum L.). Acta Physiol. Plant. 2016, 38, 152. [Google Scholar] [CrossRef]
- Kobori, M.M.R.G.; da Costa Mello, S.; de Freitas, I.S.; Silveira, F.F.; Alves, M.C.; Azevedo, R.A. Supplemental Light with Different Blue and Red Ratios in the Physiology, Yield and Quality of Impatiens. Sci. Hortic. 2022, 306, 111424. [Google Scholar] [CrossRef]
- Tarakanov, I.G.; Kosobryukhov, A.A.; Tovstyko, D.A.; Anisimov, A.A.; Shulgina, A.A.; Sleptsov, N.N.; Kalashnikova, E.A.; Vassilev, A.V.; Kirakosyan, R.N. Effects of Light Spectral Quality on the Micropropagated Raspberry Plants during Ex Vitro Adaptation. Plants 2021, 10, 2071. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.K.; Shawon, M.; Ahmed, R.; An, J.H.; Yun, Y.J.; Park, S.J.; Na, J.K.; Choi, K.Y. Influence of Substrate Composition and Container Size on the Growth of Tissue Culture Propagated Apple Rootstock Plants. Agronomy 2021, 11, 2450. [Google Scholar] [CrossRef]
- Mohammed, M.; Munir, M.; Ghazzawy, H.S. Design and Evaluation of a Smart Ex Vitro Acclimatization System for Tissue Culture Plantlets. Agronomy 2023, 13, 78. [Google Scholar] [CrossRef]
- Pascual, J.A.; Ceglie, F.; Tuzel, Y.; Koller, M.; Koren, A.; Hitchings, R.; Tittarelli, F. Organic Substrate for Transplant Production in Organic Nurseries. A Review. Agron. Sustain. Dev. 2018, 38, 35. [Google Scholar] [CrossRef] [Green Version]
- Neri, J.C.; Meléndez-Mori, J.B.; Tejada-Alvarado, J.J.; Vilca-Valqui, N.C.; Huaman-Huaman, E.; Oliva, M.; Goñas, M. An Optimized Protocol for Micropropagation and Acclimatization of Strawberry (Fragaria× Ananassa Duch.) Variety ‘Aroma.’. Agronomy 2022, 12, 968. [Google Scholar] [CrossRef]
- Bouzo, C.A.; Favaro, J.C. Container Size Effect on the Plant Production and Precocity in Tomato (Solanum Lycopersicum L.). Bulg. J. Agric. Sci. 2015, 21, 325–332. [Google Scholar]
- Oh, H.J.; Park, Y.G.; Park, J.E.; Jeong, B.R. Effect of Cell Size on Growth and Development of Plug Seedlings of Three Indigenous Medicinal Plants. J. Bio-Environ. Control 2014, 23, 71–76. [Google Scholar] [CrossRef]
- Kumar, R.R.; Purohit, V.K.; Prasad, P.; Nautiyal, A.R. Efficient In Vitro Propagation Protocol of Swertia Chirayita (Roxb. Ex Fleming) Karsten: A Critically Endangered Medicinal Plant. Natl. Acad. Sci. Lett. 2018, 41, 123–127. [Google Scholar] [CrossRef] [Green Version]
- Modgil, M.; Sharma, T.; Thakur, M. Commercially Feasible Protocol for Rooting and Accimatization of Micropropagated Apple Rootstocks. In Proceedings of the Acta Horticulturae; International Society for Horticultural Science (ISHS): Leuven, Belgium, 2009; pp. 209–214. [Google Scholar]
- Smith, R.H. Chapter 5-Contamination, 3rd ed.; Academic Press: San Diego, CA, USA, 2013; pp. 53–62. ISBN 978-0-12-415920-4. [Google Scholar]
- Ngezahayo, F.; Liu, B. Axillary Bud Proliferation Approach for Plant Biodiversity Conservation and Restoration. Int. J. Biodivers. 2014, 2014, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Mežaka, I.; Kronberga, A.; Nakurte, I.; Taškova, I.; Jakovels, D.; Primavera, A. Genetic, Chemical and Morphological Variability of Chamomile (Chamomilla Recutita L.) Populations of Latvia. Ind. Crops Prod. 2020, 154, 112614. [Google Scholar] [CrossRef]
- Olaitan, M.M.; Mangse, G.; Ogbaga, C.C.; Uthman, T.O. Gibberellic Acid Influences Growth Indices and Biochemical Parameters in Micropropagated Ocimum Gratissimum L. Explants. J. Med. Plants Econ. Dev. 2022, 6, 7. [Google Scholar] [CrossRef]
- Pepe, M.; Hesami, M.; Jones, A.M. Machine Learning-Mediated Development and Optimization of Disinfection Protocol and Scarification Method for Improved In Vitro Germination of Cannabis Seeds. Plants 2021, 10, 2397. [Google Scholar] [CrossRef] [PubMed]
- Cortés-Fernández, I.; Cerrato, M.D.; Ribas-Serra, A.; Cardona, C.; González, C.; Gil, L. Evidence of Interpopulation Variation in the Germination of Eryngium Maritimum L.(Apiaceae). Plant Ecol. 2021, 222, 1101–1112. [Google Scholar] [CrossRef]
- Cortés-Fernández, I.; Cerrato, M.D.; Ribas-Serra, A.; Gil Vives, L. Floral Traits and Reproductive Success Variation among Inflorescence Orders in Eryngium Maritimum. Plant Biol. 2022, 24, 249–258. [Google Scholar] [CrossRef] [PubMed]
- Aviziene, D.; Pakalnis, R.; Sendzikaite, J. Status of Red-Listed Species Eryngium Maritimum L. on the Lithuanian Coastal Dunes. In Proceedings of the 7th International Conference Environmental Engineering, Vilnius, Lithuania, 22–23 May 2008; pp. 22–28. [Google Scholar]
- Raturi, M.K.; Thakur, A. Silver Nitrate and Silver-Thiosulphate Mitigates Callus and Leaf Abscission during Shisham Clonal Micro-Propagation. J. Plant Biotechnol. 2021, 48, 173–178. [Google Scholar] [CrossRef]
- Arab, M.M.; Yadollahi, A.; Eftekhari, M.; Ahmadi, H.; Akbari, M.; Khorami, S.S. Modeling and Optimizing a New Culture Medium for In Vitro Rooting of G×N15 Prunus Rootstock Using Artificial Neural Network-Genetic Algorithm. Sci. Rep. 2018, 8, 9977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabral, N.N.; Pescador, R.; Pinheiro, M.V.M.; Ornellas, T.S.; Rizzolo, R.G.; Bordallo, S.U.; Guterres, S.M.; Gris, T.; Schvambach, M.I.; de Souza, P.F. Different Spectral Qualities Do Not Influence the in Vitro and Ex Vitro Survival of Epidendrum Denticulatum Barb. Rod.: A Brazilian Orchid. Vegetos 2022, 1–15. [Google Scholar] [CrossRef]
- Poorter, H.; Bühler, J.; van Dusschoten, D.; Climent, J.; Postma, J.A. Pot Size Matters: A Meta-Analysis of the Effects of Rooting Volume on Plant Growth. Funct. Plant Biol. 2012, 39, 839–850. [Google Scholar] [CrossRef] [Green Version]
- Łabuz, T.A. Evaluation of Past and Present Sea Holly (Eryngium Maritimum) Habitats on Polish Coastal Dunes. Acta Univ. Latv. 2007, 723, 114. [Google Scholar]
- Rizwan, H.M.; Irshad, M.; He, B.; Liu, S.; Lu, X.; Sun, Y.; Qiu, D. Role of Reduced Nitrogen for Induction of Embryogenic Callus Induction and Regeneration of Plantlets in Abelmoschus esculentus L. South African J. Bot. 2020, 130, 300–307. [Google Scholar] [CrossRef]
- Shekhawat, M.S.; Mehta, S.R.; Manokari, M.; Priyadharshini, S.; Badhepuri, M.K.; Jogam, P.; Dey, A.; Rajput, B.S. Morpho-Anatomical and Physiological Changes of Indian Sandalwood (Santalum Album L.) Plantlets in Ex Vitro Conditions to Support Successful Acclimatization for Plant Mass Production. Plant Cell, Tissue Organ Cult. 2021, 147, 423–435. [Google Scholar] [CrossRef]
- Singh, V.P.; Singh, M.; Singh, D. V Growth, Yield and Quality of Peppermint (Mentha x Piperita L.) as Influenced by Planting Time. J. Herbs. Spices Med. Plants 1998, 5, 33–39. [Google Scholar] [CrossRef]
- Olesen, J.E.; Trnka, M.; Kersebaum, K.C.; Skjelvåg, A.O.; Seguin, B.; Peltonen-Sainio, P.; Rossi, F.; Kozyra, J.; Micale, F. Impacts and Adaptation of European Crop Production Systems to Climate Change. Eur. J. Agron. 2011, 34, 96–112. [Google Scholar] [CrossRef]
Figure 1.
Weather conditions during field trials in 2021 and 2022. Bars represent the monthly sum of precipitation; whiskers represent the minimum and maximum temperatures registered within a month (data source: Priekuli meteorological station, Latvia).
Figure 1.
Weather conditions during field trials in 2021 and 2022. Bars represent the monthly sum of precipitation; whiskers represent the minimum and maximum temperatures registered within a month (data source: Priekuli meteorological station, Latvia).

Figure 2.
The scale for visual assessment of root development (scores from left: 1 for 1–2 cm long roots; 2 for 2–4 cm long roots; 3 for >4 cm long roots).
Figure 2.
The scale for visual assessment of root development (scores from left: 1 for 1–2 cm long roots; 2 for 2–4 cm long roots; 3 for >4 cm long roots).

Figure 3.
Development of microbial infection after in vitro initiation of E. maritimum seeds pre-treated with multistep sterilization methods (capital letters A, B, C, D, and E refer to the treatments listed in Table 2). The infection rate 14 days after treatment, marked with the same lowercase letters (a–c), did not significantly differ according to Tukey’s test (>0.05). (n = 4).
Figure 3.
Development of microbial infection after in vitro initiation of E. maritimum seeds pre-treated with multistep sterilization methods (capital letters A, B, C, D, and E refer to the treatments listed in Table 2). The infection rate 14 days after treatment, marked with the same lowercase letters (a–c), did not significantly differ according to Tukey’s test (>0.05). (n = 4).

Figure 4.
Cumulative germination percentage of pooled seeds from different origins during the 26-day period. Whiskers indicate the standard deviation. (n = 5).
Figure 4.
Cumulative germination percentage of pooled seeds from different origins during the 26-day period. Whiskers indicate the standard deviation. (n = 5).

Figure 5.
Final germination of Eryngium maritimum seeds of various origins. Final germination rates marked with the same lowercase letter (a–c) do not significantly differ according to Tukey’s test (>0.05). (n = 4).
Figure 5.
Final germination of Eryngium maritimum seeds of various origins. Final germination rates marked with the same lowercase letter (a–c) do not significantly differ according to Tukey’s test (>0.05). (n = 4).

Figure 6.
The effect of selected plant growth regulators (6-benzylaminopurine—BAP, meta-Topolin—mT, N6-(2-isopentyl)adenine—2iP, Zeatin—Z, Kinetin—Kin) and MS media modifications (MS media, MS media with NH4NO3 concentration reduced in half (AMS) and MS media with both NH4NO3 and KNO3 reduced in half (ABMS)) on shoot multiplication. Median values are represented by horizontal lines within boxplots. Whiskers indicate the 95% confidence interval of the median. Dots represent the outliers. Treatments tested on the same macrosalt media composition indicated with the same letter do not differ according to Tuckey’s test (>0.05). (n = 3).
Figure 6.
The effect of selected plant growth regulators (6-benzylaminopurine—BAP, meta-Topolin—mT, N6-(2-isopentyl)adenine—2iP, Zeatin—Z, Kinetin—Kin) and MS media modifications (MS media, MS media with NH4NO3 concentration reduced in half (AMS) and MS media with both NH4NO3 and KNO3 reduced in half (ABMS)) on shoot multiplication. Median values are represented by horizontal lines within boxplots. Whiskers indicate the 95% confidence interval of the median. Dots represent the outliers. Treatments tested on the same macrosalt media composition indicated with the same letter do not differ according to Tuckey’s test (>0.05). (n = 3).

Figure 7.
The effect of selected plant growth regulators (6-benzylaminopurine—BAP, meta-Topolin—mT) and MS media modifications (MS media, MS media with NH4NO3 concentration reduced in half (AMS) and MS media with both NH4NO3 and KNO3 reduced in half (ABMS)) on amount of regenerated small (10–15 mm) and optimal (>16 mm) length shoots. Median values are represented by horizontal lines within boxplots. Whiskers indicate the 95% confidence interval of the median. Treatments tested on the same macrosalt media composition indicated with the same letter do not differ according to Tuckey’s test (>0.05). (n = 3).
Figure 7.
The effect of selected plant growth regulators (6-benzylaminopurine—BAP, meta-Topolin—mT) and MS media modifications (MS media, MS media with NH4NO3 concentration reduced in half (AMS) and MS media with both NH4NO3 and KNO3 reduced in half (ABMS)) on amount of regenerated small (10–15 mm) and optimal (>16 mm) length shoots. Median values are represented by horizontal lines within boxplots. Whiskers indicate the 95% confidence interval of the median. Treatments tested on the same macrosalt media composition indicated with the same letter do not differ according to Tuckey’s test (>0.05). (n = 3).

Figure 8.
The effect of selected plant growth regulators (6-benzylaminopurine—BAP, meta-topolin—topolin—mT, N6-(2-isopentyl)adenine——2iP, zeatin—zeatin—Z, kinetin—kinetin—Kin) and MS media modifications (MS media, MS media with NH4NO3 concentration reduced by half (AMS), and MS media with both NH4NO3 and KNO3 reduced by half (ABMS)) on callus ball size. Median values are represented by horizontal lines within boxplots. Whiskers indicate the 95% confidence interval of the median. Dots represent the outliers. Treatments tested on the same macrosalt media composition indicated with the same letter do not differ according to Tukey’s test (>0.05). (n = 3).
Figure 8.
The effect of selected plant growth regulators (6-benzylaminopurine—BAP, meta-topolin—topolin—mT, N6-(2-isopentyl)adenine——2iP, zeatin—zeatin—Z, kinetin—kinetin—Kin) and MS media modifications (MS media, MS media with NH4NO3 concentration reduced by half (AMS), and MS media with both NH4NO3 and KNO3 reduced by half (ABMS)) on callus ball size. Median values are represented by horizontal lines within boxplots. Whiskers indicate the 95% confidence interval of the median. Dots represent the outliers. Treatments tested on the same macrosalt media composition indicated with the same letter do not differ according to Tukey’s test (>0.05). (n = 3).

Figure 9.
The effect of concentration of indole-3-acetic acid (IAA) added to two MS media with NH4NO3 concentration reduced in half (AMS) and MS media with both NH4NO3 and KNO3 reduced in half (ABMS) on the number of roots per explant. Median values are represented by horizontal lines within boxplots. Whiskers indicate the 95% confidence interval of the median. Dots represent the outliers. Treatments tested on the same macrosalt media composition indicated with the same letter do not differ according to Tukey’s test (>0.05). (n = 3).
Figure 9.
The effect of concentration of indole-3-acetic acid (IAA) added to two MS media with NH4NO3 concentration reduced in half (AMS) and MS media with both NH4NO3 and KNO3 reduced in half (ABMS) on the number of roots per explant. Median values are represented by horizontal lines within boxplots. Whiskers indicate the 95% confidence interval of the median. Dots represent the outliers. Treatments tested on the same macrosalt media composition indicated with the same letter do not differ according to Tukey’s test (>0.05). (n = 3).

Figure 10.
The effect of lamp type on plantlet survival (a) and the number of developed leaves per plant (b) 21 days after ex vitro transfer. Median values are represented by horizontal lines within boxplots. Whiskers indicate the 95% confidence interval of the median. Dots represent the outliers. Mericlones and pot cell diameters indicated with the same letter do not differ according to Tukey’s test (>0.05). (n = 3).
Figure 10.
The effect of lamp type on plantlet survival (a) and the number of developed leaves per plant (b) 21 days after ex vitro transfer. Median values are represented by horizontal lines within boxplots. Whiskers indicate the 95% confidence interval of the median. Dots represent the outliers. Mericlones and pot cell diameters indicated with the same letter do not differ according to Tukey’s test (>0.05). (n = 3).

Figure 11.
The effect of substrate (soil, sand, vermiculite, and perlite) at a ratio of 3:1:1:1 (B1) and soil, cow manure, vermiculite, and perlite at a ratio of 3:1:1:1 (B2) on mericlone M2 plantlet survival (a) and the number of developed leaves per plant (b) 21 days after transfer to ex vitro. Median values are represented by horizontal lines within boxplots. Whiskers indicate the 95% confidence interval of the median. Dots represent the outliers. Mericlones and pot cell diameters indicated with the same letter do not differ according to Tukey’s test (>0.05). (n = 3).
Figure 11.
The effect of substrate (soil, sand, vermiculite, and perlite) at a ratio of 3:1:1:1 (B1) and soil, cow manure, vermiculite, and perlite at a ratio of 3:1:1:1 (B2) on mericlone M2 plantlet survival (a) and the number of developed leaves per plant (b) 21 days after transfer to ex vitro. Median values are represented by horizontal lines within boxplots. Whiskers indicate the 95% confidence interval of the median. Dots represent the outliers. Mericlones and pot cell diameters indicated with the same letter do not differ according to Tukey’s test (>0.05). (n = 3).

Figure 12.
The effect of pot cell diameter (3.7 cm and 2.8 cm) on plantlet survival (a) and the number of developed leaves per plant (b) 21 days after transfer to ex vitro. Median values are represented by horizontal lines within boxplots. Whiskers indicate the 95% confidence interval of the median. Dots represent the outliers. Mericlones and pot cell diameters indicated with the same letter do not differ according to Tukey’s test (>0.05). (n = 3).
Figure 12.
The effect of pot cell diameter (3.7 cm and 2.8 cm) on plantlet survival (a) and the number of developed leaves per plant (b) 21 days after transfer to ex vitro. Median values are represented by horizontal lines within boxplots. Whiskers indicate the 95% confidence interval of the median. Dots represent the outliers. Mericlones and pot cell diameters indicated with the same letter do not differ according to Tukey’s test (>0.05). (n = 3).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Seed sources of the studied Eryngium maritimum accessions.
| Origin | Source | Coordinates |
|---|---|---|
| Jellito, Germany | Commercial | |
| Magic seeds, Germany | Commercial | |
| Suffolk herbs, United Kingdom | Commercial | |
| Kihnu, Estonia | Wild | N58°7′49″ E23°57′32″ |
| Saaremaa, Estonia | Wild | N58°28′36″ E21°54′59″ |
| Ziemupe, Latvia | Wild | N56°48′4″ E21°4′4″ |
| Užava, Latvia | Wild | N57°14′49″ E21°25′52″ |
| Italy | University of Pisa botanical garden collection |
Table 2.
Experimental variants for Eryngium maritimum seed disinfection.
| Experimental Variant | Diluted Commercial Detergent “Fairy” | KMnO4 (0.01%) | Commercial Bleach “ACE” |
|---|---|---|---|
| A | 15 min 20 °C | 1 h | 10 min |
| B | 15 min 20 °C | 2 h | 10 min |
| C | 15 min 40 °C | 1.5 h | 11 min |
| D | 15 min 40 °C | 3 h | 11 min |
| E | 180 min 40 °C | 3 h | 11 min |
Table 3.
Differences in root development before ex vitro adaptation and survival, and the number of developed leaves 21 days after ex vitro adaptation, between tested mericlones.
Table 3.
Differences in root development before ex vitro adaptation and survival, and the number of developed leaves 21 days after ex vitro adaptation, between tested mericlones.
| Survival, % | Root Development (Score, 1–3) | Survival, % | Number of Developed Leaves per Plant |
|---|---|---|---|
| M1 | 1.3a 1 | 99.0a | 5.7a |
| M2 | 1.3a | 99.4a | 6.2a |
| M3 | 1.8a | 99.0a | 6.5a |
1 Means tracked by the same letters are not significantly different according to Tukey’s test.
Table 4.
Survival (%) of plantlets of three mericlones in field conditions (autumn 2021 and spring 2022) based on planting date (n = 40, three replications).
Table 4.
Survival (%) of plantlets of three mericlones in field conditions (autumn 2021 and spring 2022) based on planting date (n = 40, three replications).
| Planting Date | June 2021 | July 2021 | ||
|---|---|---|---|---|
| Survival, % | Autumn 2021 | Spring 2022 | Autumn 2021 | Spring 2022 |
| M1 | 90.8ab 1 | 72.5b | - | - |
| M2 | 99.2a | 77.5b | 55.8c | 22.5d |
| M3 | 86.7ab | 72.5b | - | - |
1 Means tracked by the same letters are not significantly different according to Tukey’s test.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Mežaka, I.; Kļaviņa, D.; Kaļāne, L.; Kronberga, A. Large-Scale In Vitro Propagation and Ex Vitro Adaptation of the Endangered Medicinal Plant Eryngium maritimum L. Horticulturae 2023, 9, 271. https://doi.org/10.3390/horticulturae9020271
AMA Style
Mežaka I, Kļaviņa D, Kaļāne L, Kronberga A. Large-Scale In Vitro Propagation and Ex Vitro Adaptation of the Endangered Medicinal Plant Eryngium maritimum L. Horticulturae. 2023; 9(2):271. https://doi.org/10.3390/horticulturae9020271
Chicago/Turabian StyleMežaka, Ieva, Dace Kļaviņa, Laura Kaļāne, and Arta Kronberga. 2023. "Large-Scale In Vitro Propagation and Ex Vitro Adaptation of the Endangered Medicinal Plant Eryngium maritimum L." Horticulturae 9, no. 2: 271. https://doi.org/10.3390/horticulturae9020271
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.