

Standardization of an In Vitro Seed Germination Protocol Compared to Acid Scarification and Cold Stratification Methods for Different Raspberry Genotypes

 ,
,  and
and 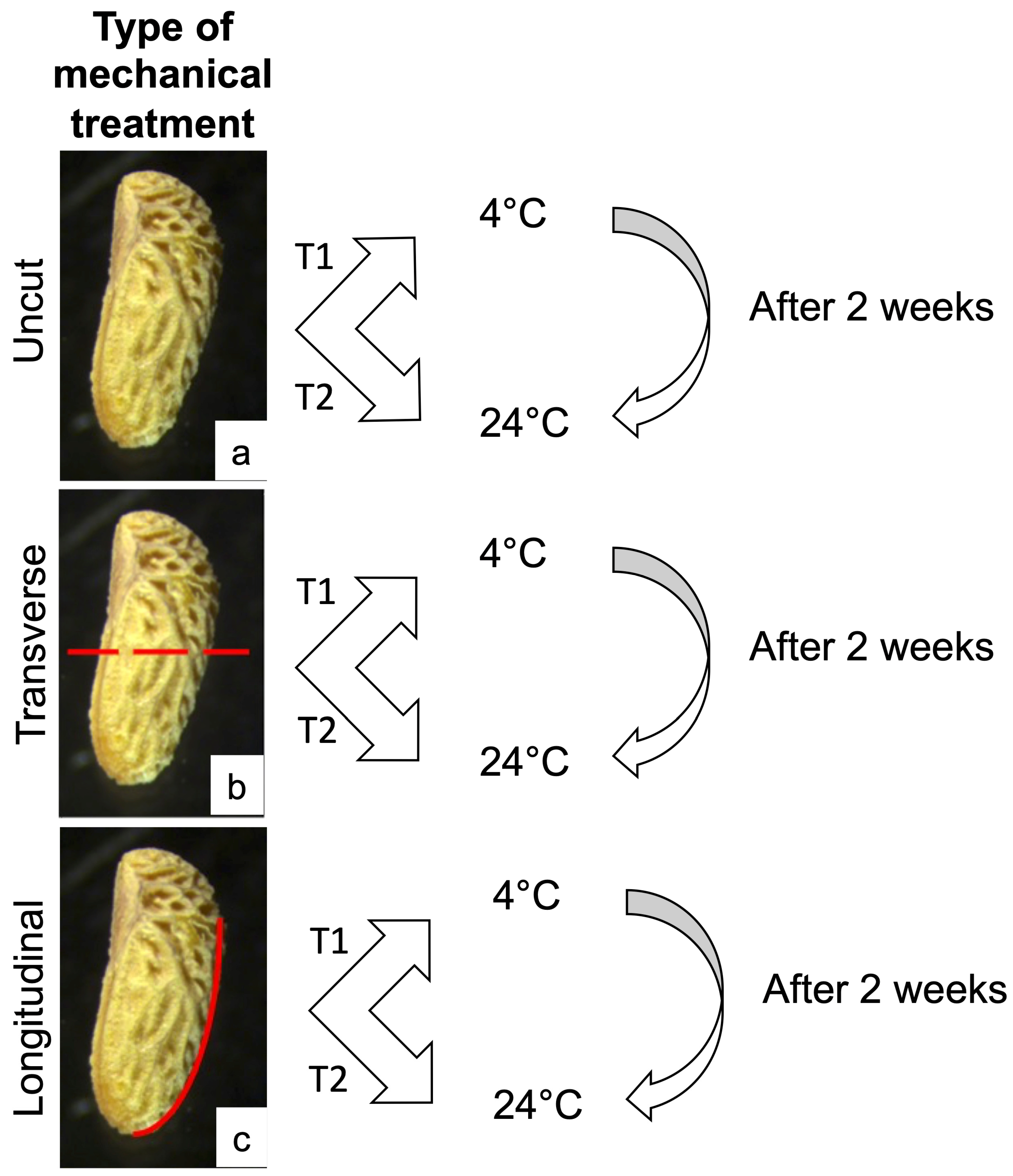{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Seed Source
2.2. Seed Germination through Scarification and Stratification
2.2.1. Seed Scarification
2.2.2. Seed Cold Stratification and Sowing
2.3. In Vitro Seed Germination
2.4. Seedlings In Vitro Rooting and Acclimatization
2.5. Application of the Optimized In Vitro Seed Germination Protocol to Combination Crosses
2.6. Statistical Analysis
3. Results
3.1. Seed Germination after Scarification and Stratification
3.2. In Vitro Seed Germination Experiments
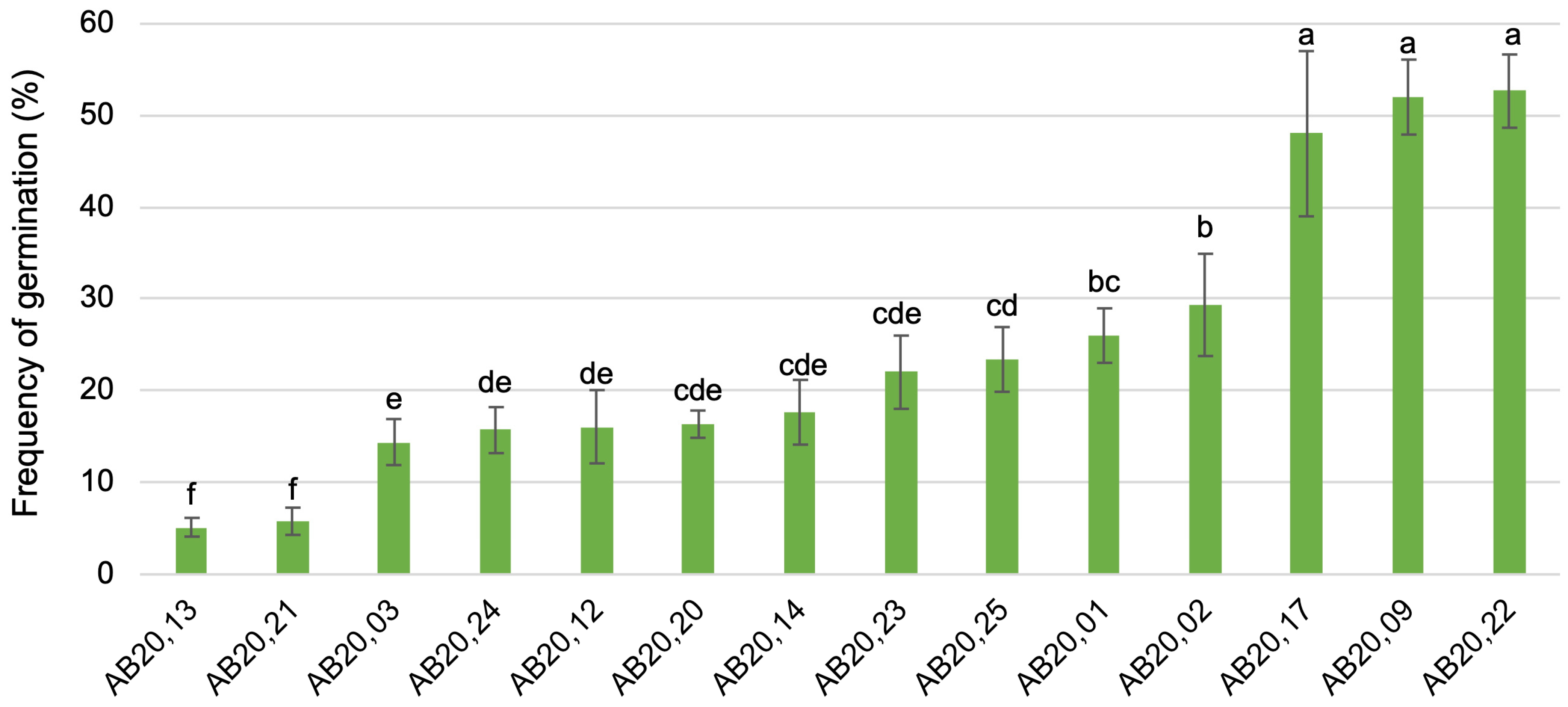
3.3. Application of the Optimized In Vitro Seed Germination Protocol to Combination Crosses
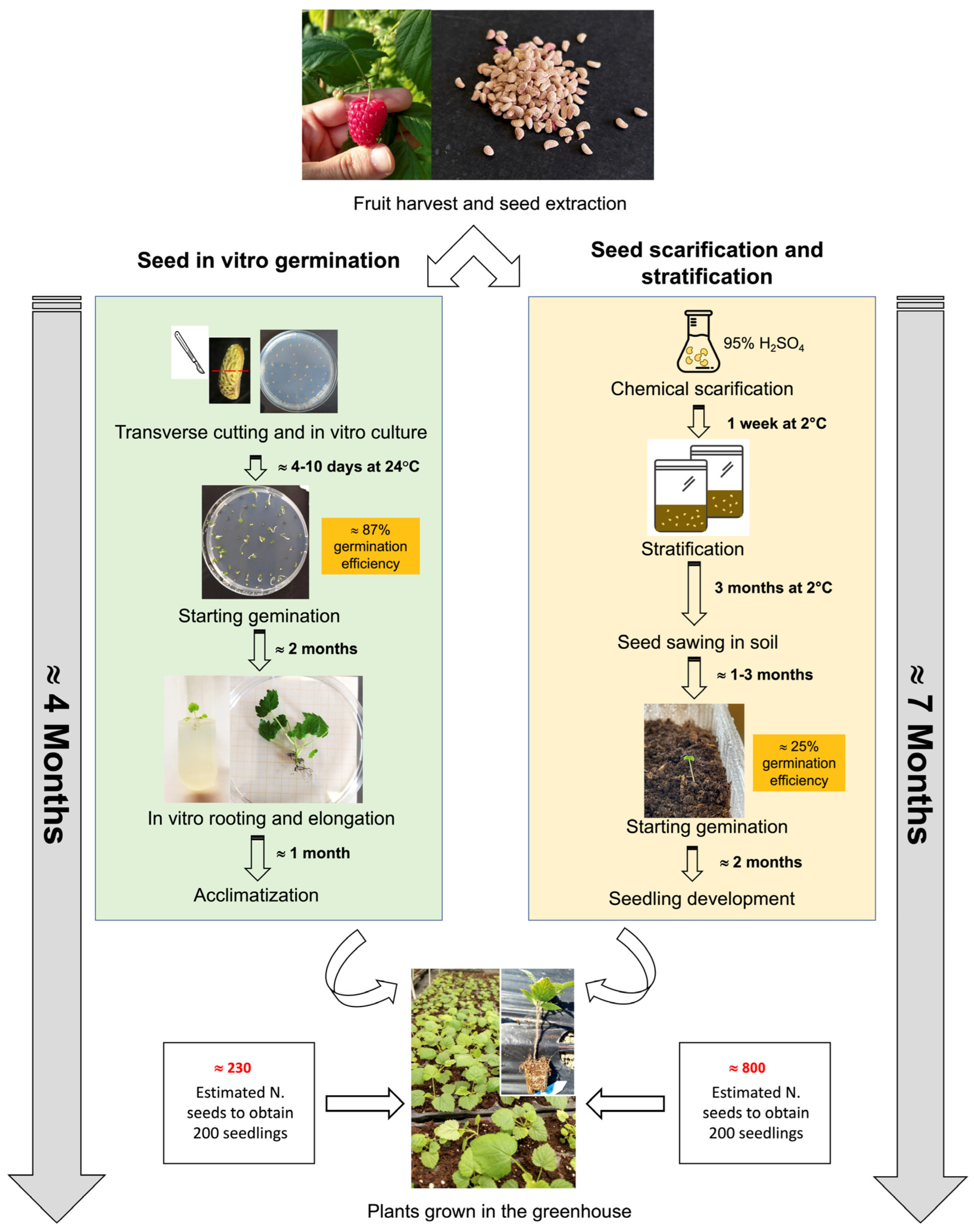
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hall, D.; Shepherd, T.; Fountain, M.; Vétek, G.; Birch, N.; Jorna, C.; Farman, D.; Cross, J. Investigation of attraction of raspberry cane midge, Resseliella theobaldi, to volatiles from wounded raspberry primocanes. WPRS Bull. 2011, 70, 1–9. [Google Scholar]
- Jennings, D.L. Raspberries and Blackberries: Their Breeding, Diseases and Growth; Academic Press: London, UK, 1988; p. 230. [Google Scholar]
- Graham, J.; Smith, K.; MacKenzie, K.; Jorgenson, L.; Hackett, C.; Powell, W. The construction of a genetic linkage map of red raspberry (Rubus idaeus subsp. idaeus) based on AFLPs, genomic-SSR and EST-SSR markers. Theor. Appl. Genet. 2004, 109, 740–749. [Google Scholar] [PubMed]
- Faostat. Available online: http:/FAOSTAT.FAO.ORG (accessed on 5 December 2022).
- Sabbadini, S.; Capocasa, F.; Battino, M.; Mazzoni, L.; Mezzetti, B. Improved nutritional quality in fruit tree species through traditional and biotechnological approaches. Trends Food Sci. Technol. 2021, 117, 125–138. [Google Scholar] [CrossRef]
- Palonen, P.; Weber, C. Fruit Color Stability, Anthocyanin Content, and Shelf Life Were Not Correlated with Ethylene Production Rate in Five Primocane Raspberry Genotypes. Sci. Hortic. 2019, 247, 9–16. [Google Scholar] [CrossRef]
- Graham, J.; Jennings, N. Raspberry Breeding. In Breeding Plantation Tree Crops: Temperate Species; Springer: New York, NY, USA, 2009; pp. 233–248. [Google Scholar]
- Contreras, E.; Grez, J.; Gambardella, M. Scarification and Stratification Protocols for Raspberry (Rubus idaeus L.) Seed Germination. Acta Hortic. 2016, 1133, 153–157. [Google Scholar] [CrossRef]
- Zurawicz, E.; Masny, A.; Kubik, J.; Lewandowski, M. Germination of Red Raspberry Seeds as Affected by Origin and Chemical Scarification. Hortic. Sci. 2017, 44, 133–140. [Google Scholar] [CrossRef] [Green Version]
- Tiwari, A.K.; Tiwari, T.N.; Prasad, S.R. Seed dormancy in ornamental plants: A review. Indian J. Agric. Sci. 2016, 86, 580–592. [Google Scholar]
- Wada, S.; Reed, B.M. Standardizing Germination Protocols for Diverse Raspberry and Blackberry Species. Sci. Hortic. 2011, 132, 42–49. [Google Scholar] [CrossRef]
- Wada, S.; Kennedy, J.A.; Reed, B.M. Seed-coat anatomy and proanthocyanidins contribute to the dormancy of Rubus seed. Sci. Hortic. 2011, 130, 762–768. [Google Scholar] [CrossRef]
- Warr, H.J.; Savory, D.R.; Bal, A.K. Germination studies of bakeapple (cloud-berry) seeds. Can. J. Plant Sci. 1979, 59, 69–74. [Google Scholar] [CrossRef]
- Taylor, K. Biological Flora of the British Isles: Rubus Vestitus Weihe. J. Ecol. 2005, 93, 1249–1262. [Google Scholar] [CrossRef]
- Moore, J.N.; Brown, G.R.; Lundergan, C. Effect of Duration of Acid Scarification on Endocarp Thickness and Seedling Emergence of Blackberries1. HortScience 1974, 9, 204–205. [Google Scholar] [CrossRef]
- Nesme, X. Respective effects of endocarp, testa and endosperm, and embryo on the germination of raspberry (Rubus idaeus L.) seeds. Can. J. Plant Sci. 1985, 65, 125–130. [Google Scholar] [CrossRef] [Green Version]
- Peacock, D.N.; Hummer, K.E. Pregermination Studies with Liquid Nitrogen and Sulfuric Acid on Several Rubus Species. HortScience 1996, 31, 238–239. [Google Scholar] [CrossRef] [Green Version]
- Grzesik, M.; Romanowska-Duda, Z. Quality of Seeds as a Key to Commodity Cultivation of Lavatera Thuringiaca L.—Plants with High Potential for Multi Directional Use. Acta Innov. 2015, 15, 5–12. [Google Scholar]
- Ke, S.; Skirvin, R.M.; McPheeters, K.D.; Otterbacher, A.G.; Galletta, G. In vitro germination and growth of Rubus seeds and embryos. HortScience 1985, 20, 1047–1049. [Google Scholar] [CrossRef]
- Masny, A.; Kubik, J.; Żurawicz, E. Seed Germination of Raspberry (Rubus Idaeus L.) Depending on the Age of Seeds and Hybridization Partners. J. Hortic. Res. 2022, 3, 61–66. [Google Scholar] [CrossRef]
- Koornneef, M.; Bentsink, L.; Hilhorst, H. Seed dormancy and germination. Curr. Opin. Plant Biol. 2002, 5, 33–36. [Google Scholar] [CrossRef] [Green Version]
- Finkelstein, R.; Reeves, W.; Ariizumi, T.; Steber, C. Molecular aspects of seed dormancy. Annu. Rev. Plant Biol. 2008, 59, 387–415. [Google Scholar] [CrossRef] [Green Version]
- Rehman, S.; Cho, H.R.; Jamil, M.; Yun, S.J. Effect of GA and ABA on germination behavior of Black Raspberry (Rubus coreanus Miquel) Seeds. Pak. J. Bot. 2011, 43, 2811–2816. [Google Scholar]
- Batak, I.; Dević, M.; Gibal, Z.; Grubišić, D.; Poff, K.L.; Konjević, R. The effects of potassium nitrate and NO-donors on phytochrome A-and phytochrome B-specific induced germination of Arabidopsis thaliana seeds. Seed Sci. Res. 2002, 12, 253–259. [Google Scholar] [CrossRef]
- Nelson, D.C.; Riseborough, J.A.; Flematti, G.R.; Stevens, J.; Ghisalberti, E.L.; Dixon, K.W.; Smith, S.M. Karrikins discovered in smoke trigger Arabidopsis seed germination by a mechanism requiring gibberellic acid synthesis and light. Plant Physiol. 2009, 149, 863–873. [Google Scholar] [CrossRef] [Green Version]
- Sharma, D.R.; Kaur, R.; Kumar, K. Embryo rescue in plants—A review. Euphytica 1996, 89, 325–337. [Google Scholar] [CrossRef]
- Mroginski, L.; Dolce, N.; Sansberro, P.; Luna, C.; Gonzalez, A.; Rey, H. Cryopreservation of Ilex Immature Zygotic Embryos. In Plant Embryo Culture; Thorpe, T., Yeung, E., Eds.; Humana Press: Totowa, NJ, USA, 2011; pp. 215–225. [Google Scholar]
- Dolce, N.R.; Mroginski, L.A.; Rey, H.Y. Enhanced Seed Germination of Ilex Dumosa R. (Aquifoliaceae) through in Vitro Culture of Cut Pyrenes. HortScience 2011, 46, 278–281. [Google Scholar] [CrossRef]
- Fay, M.F. Conservation of rare and endangered plants using in vitro methods. Vitr. Cell. Dev. Biol. Plant 1992, 28, 1–4. [Google Scholar] [CrossRef]
- Andrianjaka-Camps, Z.N.; Baumgartner, D.; Camps, C.; Guyer, E.; Arrigoni, E.; Carlen, C. Prediction of raspberries puree quality traits by Fourier transform infrared spectroscopy. LWT Food Sci. Technol. 2015, 6, 1056–1062. [Google Scholar] [CrossRef]
- Maksimović, J.J.D.; Milivojević, J.M.; Poledica, M.M.; Nikolić, M.D.; Maksimović, V.M. Profiling antioxidant activity of two primocane fruiting red raspberry cultivars (Autumn bliss and Polka). J. Food Compos. Anal. 2013, 31, 173–179. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Jennings, D.L.; Tulloch, B.M.M. Studies on factors which promote germination of raspberry seeds. J. Exp. Bot. 1965, 16, 329–340. [Google Scholar] [CrossRef]
- Dale, A.; Jarvis, B.C. Studies on germination in raspberry (Rubus idaeus L.). Crop Res. 1983, 23, 73–81. [Google Scholar]
- Miller, A.R.; Scheereus, J.C.; Erb, P.S.; Chandler, C.K. Enhanced strawberry seed germination through in vitro culture of cut achenes. J. Am. Soc. Hortic. Sci. 1992, 117, 313–316. [Google Scholar] [CrossRef] [Green Version]
- Jennings, D.L. Some genetic factors affecting fruit development in raspberries. New Phytol. 1971, 70, 361–370. [Google Scholar] [CrossRef]
- Żurawicz, E.; Studnicki, M.; Kubik, J.; Pruski, K. A Careful Choice of Compatible Pollinizers Significantly Improves the Size of Fruits in Red Raspberry (Rubus Idaeus L.). Sci. Hortic. 2018, 235, 253–257. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pergolotti, V.; Marcellini, M.; Contreras, E.; Mezzetti, B.; Gambardella, M.; Capocasa, F.; Sabbadini, S. Standardization of an In Vitro Seed Germination Protocol Compared to Acid Scarification and Cold Stratification Methods for Different Raspberry Genotypes. Horticulturae 2023, 9, 153. https://doi.org/10.3390/horticulturae9020153
Pergolotti V, Marcellini M, Contreras E, Mezzetti B, Gambardella M, Capocasa F, Sabbadini S. Standardization of an In Vitro Seed Germination Protocol Compared to Acid Scarification and Cold Stratification Methods for Different Raspberry Genotypes. Horticulturae. 2023; 9(2):153. https://doi.org/10.3390/horticulturae9020153
Chicago/Turabian StylePergolotti, Valeria, Micol Marcellini, Elida Contreras, Bruno Mezzetti, Marina Gambardella, Franco Capocasa, and Silvia Sabbadini. 2023. "Standardization of an In Vitro Seed Germination Protocol Compared to Acid Scarification and Cold Stratification Methods for Different Raspberry Genotypes" Horticulturae 9, no. 2: 153. https://doi.org/10.3390/horticulturae9020153