
Recent Advances in Natural Deastringency and Genetic Improvement of Chinese PCNA Persimmon (Diospyros kaki)

Abstract
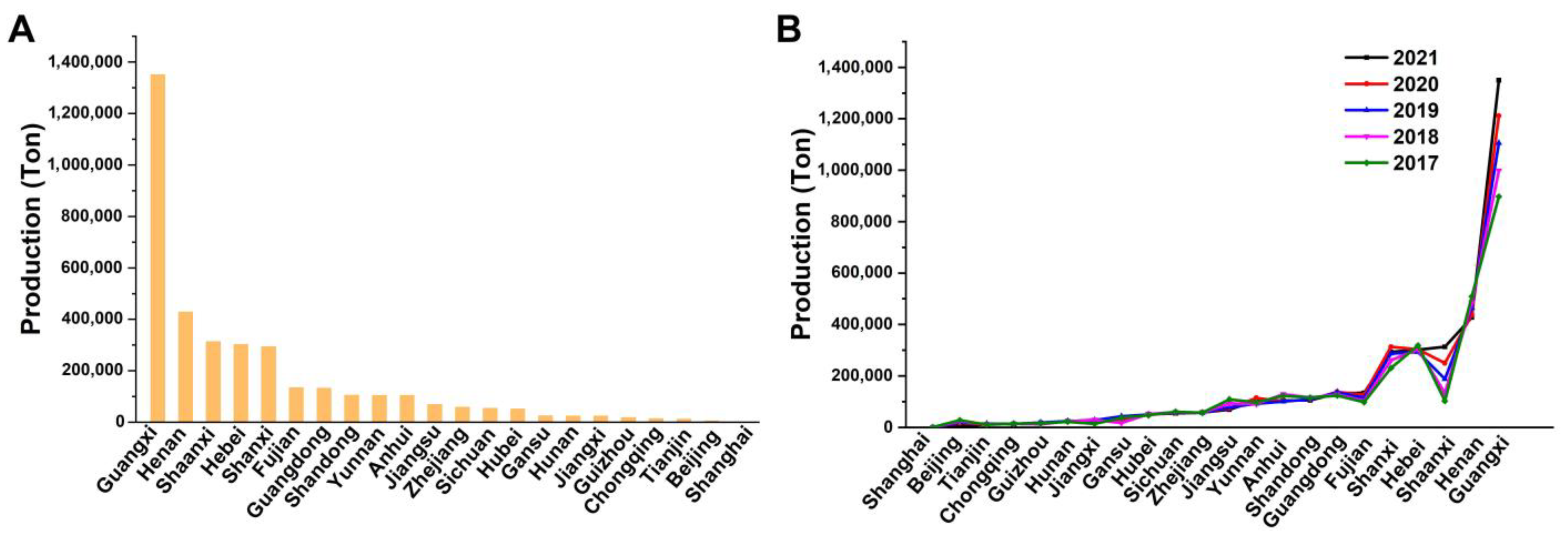
:1. Introduction
2. Tannin Composition and Content Change in C-PCNA
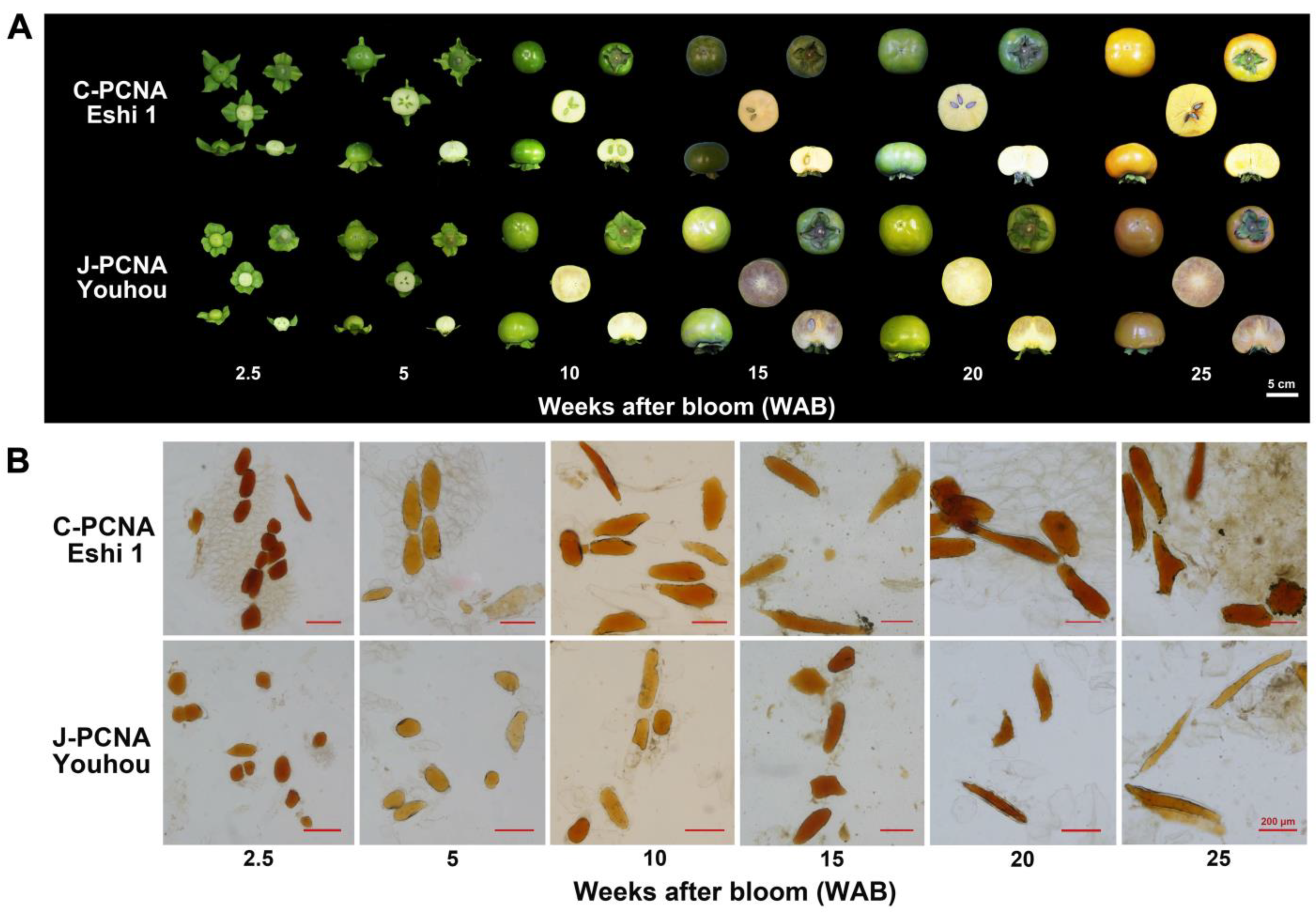
2.1. Tannin Cell Characteristics in C-PCNA
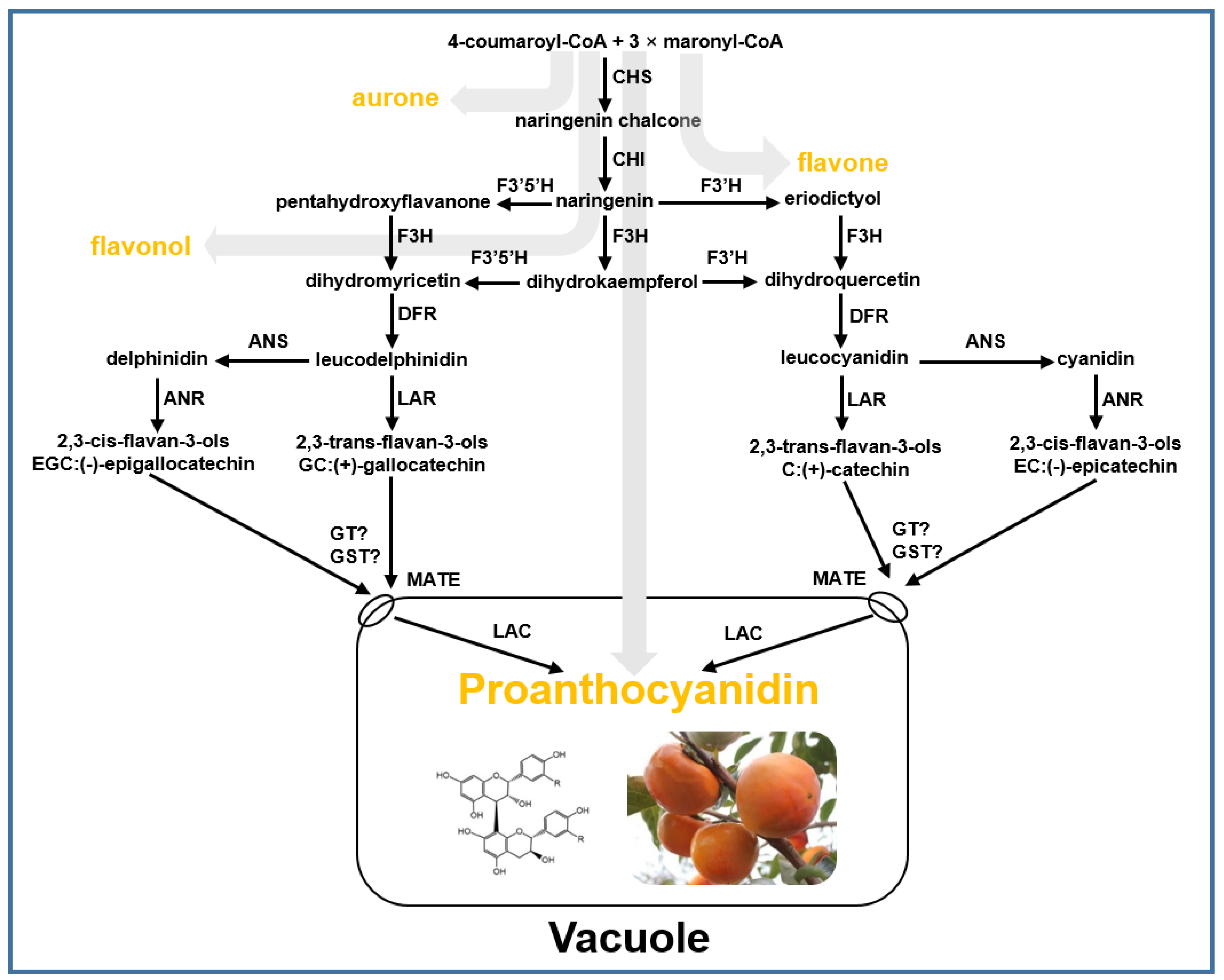
2.2. Tannin Composition in C-PCNA
2.3. Changes in Tannin Contents in C-PCNA
3. Natural Deastringency Mechanism in C-PCNA
3.1. Acetaldehyde Metabolism Is the Essential Pathway Involved in the Natural Deastringency of C-PCNA
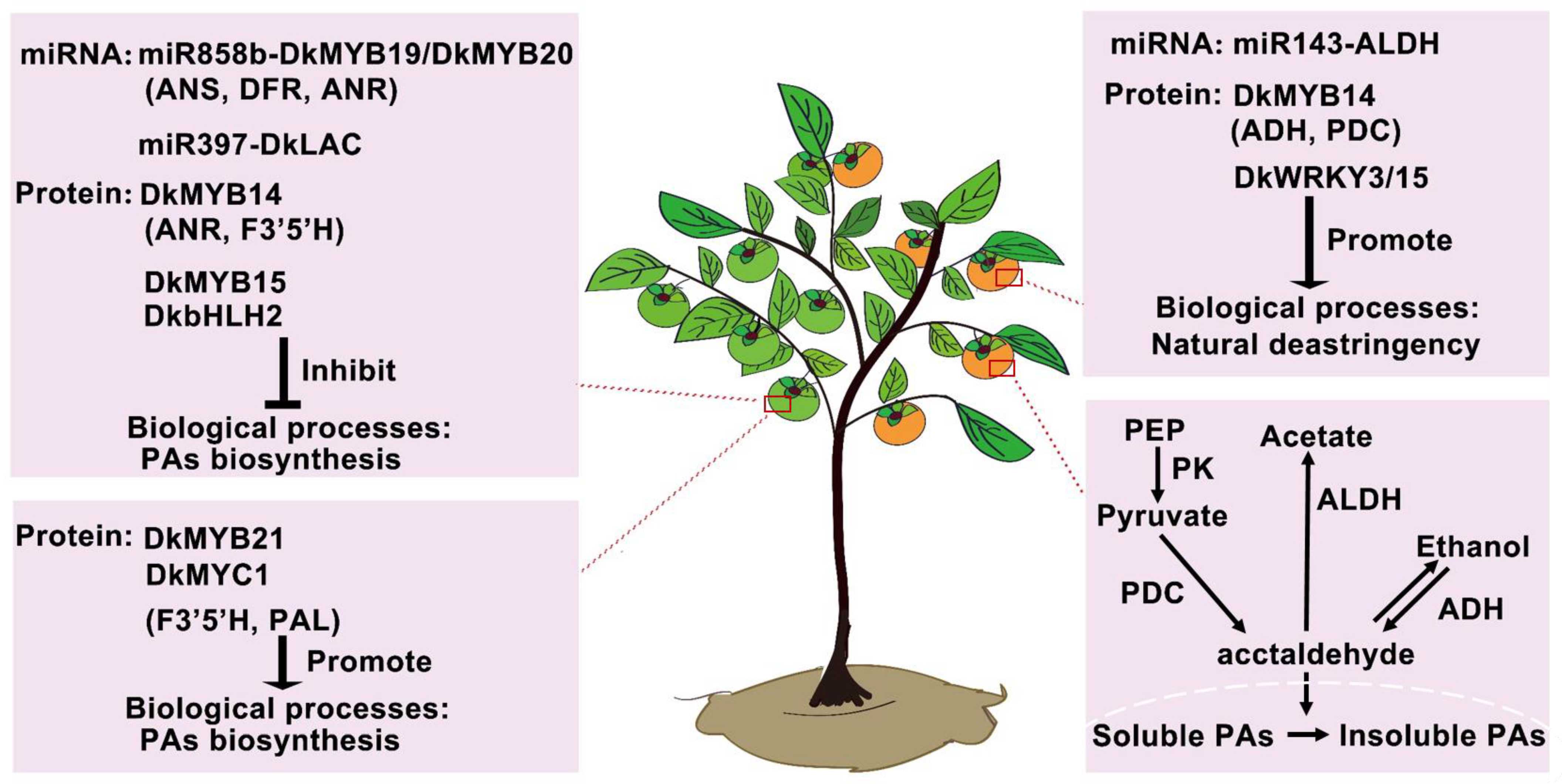
3.2. Regulation Network Is Involved in the Natural Deastringency of C-PCNA
4. Conventional Cross-Breeding of C-PCNA
4.1. Sex Classification of Chinese Persimmon Flowers
4.2. Artificial Pollination of Chinese Persimmon
4.3. Embryo Rescue Technology in Chinese Persimmon
4.4. RO2 Marker-Assisted Early Selection of the Non-Astringency/Astringency Trait in Persimmon
5. Genetic Transformation in Chinese Persimmon
5.1. The Establishment of the Persimmon Regeneration System
5.2. Establement of the Transient Expression System
5.3. Establement of the Stable Transformation System
6. Conclusions and Future Prospects
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Luo, Z.R.; Zhang, Q.L.; Xu, L.Q.; Guo, D.Y.; Chen, W.X. Fruit scientific research in New China in the past 70 years: Persimmon. J. Fruit Sci. 2019, 36, 1382–1388. [Google Scholar] [CrossRef]
- Amorim, C.; Antoniolli, L.R.; Orsi, B.; Kluge, R.A. Advances in metabolism and genetic control of astringency in persimmon (Diospyros kaki Thunb.) fruit: A review. Sci. Hortic. 2023, 308, 111561. [Google Scholar] [CrossRef]
- Akagi, T.; Katayama-Ikegami, A.; Yonemori, K. Proanthocyanidin biosynthesis of persimmon (Diospyros kaki Thunb.) fruit. Sci. Hortic. 2011, 130, 373–380. [Google Scholar] [CrossRef]
- Guan, C.F.; Zhang, Y.F.; Zhang, P.X.; Chachar, S.; Wang, R.Z.; Du, X.Y.; Yang, Y. Germplasm conservation, molecular identity and morphological characterization of persimmon (Diospyros kaki Thunb.) in the NFGP of China. Sci. Hortic. 2020, 272, 109490. [Google Scholar] [CrossRef]
- Guan, C.F.; Zhang, P.X.; Hu, C.Q.; Chachar, S.; Riaz, A.; Wang, R.Z.; Yang, Y. Genetic diversity, germplasm identification and population structure of Diospyros kaki Thunb. from different geographic regions in China using SSR markers. Sci. Hortic. 2019, 251, 233–240. [Google Scholar] [CrossRef]
- Pei, X.; Zhang, Q.L.; Guo, D.Y.; Liu, J.H.; Luo, Z.R. Development of genetic improvement in Chinese PCNA persimmon. J. Fruit Sci. 2015, 32, 313–321. [Google Scholar] [CrossRef]
- Chen, W.X. A combined transcriptome and proteome analysis for natural deastringency-related genes selection and functional characterization of DkMYB14 in Chineses PCNA persimmon. Ph.D. Thesis, Huazhong Agricuture University, Wuhan, China, 2019. [Google Scholar]
- Akagi, T.; Ikegami, A.; Tsujimoto, T.; Kobayashi, S.; Sato, A.; Kono, A.; Yonemori, K. DkMyb4 is a MYB transcription factor involved in proanthocyanidin biosynthesis in persimmon fruit. Plant Physiol. 2009, 151, 2028–2045. [Google Scholar] [CrossRef]
- Wang, Y.Z.; Liu, Y. A review on the research advances of vegetable tannin. Westleather 2014, 36, 23–30. [Google Scholar]
- Shu, C.; Zhao, H.D.; Jiao, W.X.; Fan, X.G.; Jiang, W.B. Research progress on the bioactivity of plant origin tannins. Sci. Technol. Food Ind. 2018, 39, 328–334. [Google Scholar] [CrossRef]
- Cipollini, M.L.; Stiles, E.W. Fruit rot, antifungal defense, and palatability of fleshy fruits for frugivorous birds. Ecology 1993, 74, 751–762. [Google Scholar] [CrossRef]
- Dixon, R.A. Engineering of plant natural product pathways. Curr. Opin. Plant Biol. 2005, 8, 329–336. [Google Scholar] [CrossRef]
- Santos-Buelga, C.; Scalbert, A. Proanthocyanidins and tannin-like compounds–nature, occurrence, dietary intake and effects on nutrition and health. J. Sci. Food Agric. 2000, 80, 1094–1117. [Google Scholar] [CrossRef]
- Serafini, M.; Bugianesi, R.; Maiani, G.; Valtuena, S.; De Santis, S.; Crozier, A. Plasma antioxidants from chocolate. Nature 2003, 424, 1013. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, S.E.; Frederiksen, H.; Struntze Krogholm, K.; Poulsen, L. Dietary proanthocyanidins: Occurrence, dietary intake, bioavailability, and protection against cardiovascular disease. Mol. Nutr. Food Res. 2005, 49, 159–174. [Google Scholar] [CrossRef]
- Scalbert, A.; Manach, C.; Morand, C.; Rémésy, C.; Jiménez, L. Dietary polyphenols and the prevention of diseases. Crit. Rev. Food Sci. 2005, 45, 287–306. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, T.; Ito, S. The chemical structure of kaki-tannin from immature fruit of the persimmon (Diospyros kaki L.). Agric. Biol. Chem. 1978, 42, 1637–1643. [Google Scholar] [CrossRef]
- Gorinstein, S.; Bartnikowska, E.; Kulasek, G.; Zemser, M.; Trakhtenberg, S. Dietary persimmon improves lipid metabolism in rats fed diets containing cholesterol. J. Nutr. 1998, 128, 2023–2027. [Google Scholar] [CrossRef]
- Matsumoto, K.; Yokoyama, S.; Gato, N. Bile acid-binding activity of young persimmon (Diospyros kaki) fruit and its hypolipidemic effect in mice. Phytother. Res. 2010, 24, 205–210. [Google Scholar] [CrossRef]
- Butt, M.S.; Sultan, M.T.; Aziz, M.; Naz, A.; Ahmed, W.; Kumar, N.; Imran, M. Persimmon (Diospyros kaki) fruit: Hidden phytochemicals and health claims. EXCLI J. 2015, 14, 542. [Google Scholar] [CrossRef]
- Yaqub, S.; Farooq, U.; Shafi, A.; Akram, K.; Murtaza, M.A.; Kausar, T.; Siddique, F. Chemistry and functionality of bioactive compounds present in persimmon. J. Chem. 2016, 2016, 1–13. [Google Scholar] [CrossRef]
- Aron, P.M.; Kennedy, J.A. Flavan-3-ols: Nature, occurrence and biological activity. Mol. Nutr. Food Res. 2008, 52, 79–104. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.S.; Wu, X.H.; Chen, J.P. Research progress of kaki tannin. J. Shaanxi Norm. Univ. 2008, 36, 99–105. [Google Scholar]
- Taira, S. Astringency in persimmon. Mod. Methods Plant Anal. 1996, 18, 97–110. [Google Scholar] [CrossRef]
- Yamada, M.; Taira, S.; Ohtsuki, M.; Sato, A.; Iwanami, H.; Yakushiji, H.; Wang, R.Z.; Yang, Y.; Li, G.C. Varietal differences in the ease of astringency removal by carbon dioxide gas and ethanol vapor treatments among oriental astringent persimmons of Japanese and Chinese origin. Sci. Hortic. 2002, 94, 63–72. [Google Scholar] [CrossRef]
- Guan, C.F.; Wang, M.K.; Zhang, Y.F.; Ruan, X.F.; Zhang, Q.L.; Luo, Z.R.; Yang, Y. DkWRKY interacts with pyruvate kinase gene DkPK1 and promotes natural deastringency in C-PCNA persimmon. Plant Sci. 2020, 290, 110285. [Google Scholar] [CrossRef] [PubMed]
- Yonemori, K.; Sugiura, A.; Yamada, M. Persimmon genetics and breeding. Plant Breed. Rev. 2000, 19, 191–225. [Google Scholar] [CrossRef]
- Akagi, T.; Katayama-Ikegami, A.; Kobayashi, S.; Sato, A.; Kono, A.; Yonemori, K. Seasonal abscisic acid signal and a basic leucine zipper transcription factor, DkBZIP5, regulate proanthocyanidin biosynthesis in persimmon fruit. Plant Physiol. 2012, 158, 1089–1102. [Google Scholar] [CrossRef]
- Wang, R.Z. The origin of ‘Luo Tian Tian Shi’. Chin. Fruit Tree 1983, 16–19. [Google Scholar] [CrossRef]
- Yuan, L.X.; Zhang, Q.L.; Guo, D.Y.; Luo, Z.R. Characteristics of Chinese PCNA types and their roles in science and industry of oriental persimmon. Acta Hortic. Sin. 2011, 38, 361–370. Available online: https://www.ahs.ac.cn/CN/Y2011/V38/I2/361 (accessed on 15 July 2023).
- Xu, J.C.; Zhang, Q.L.; Xu, L.Q.; Guo, D.Y.; Luo, Z.R. Recent developments in deastringency mechanism of persimmon fruit. Acta Hortic. Sin. 2016, 43, 1653–1664. [Google Scholar] [CrossRef]
- Fei, X.Q.; Zhou, L.H.; Wang, J.F. Relationship between the capacity of deastrigency and development of tannin cell in persimmon fruits. For. Res. 1996, 9, 27–31. [Google Scholar]
- Fei, X.Q.; Zhou, L.H.; Gong, B.C. Differences of the components of tannin among three types of persimmon fruits and characteristics of tannin from ‘Luotian Tianshi’. For. Res. 1999, 12, 369–373. [Google Scholar]
- Yang, Y.; Ruan, X.F.; Wang, R.Z.; Li, G.C. Study on morphological characteristics and development of tannin cells in persimmon (Diospyros kaki). J. Northwest Sci-Tech Univ. Agric. For. 2003, 31, 93–99. [Google Scholar]
- Yang, Y.; Ruan, X.F.; Wang, R.Z.; Li, G.C. Classification and morphological characteristics of tannin cells in some persimmon species and varieties. Acta Bot. Boreali-Occident. Sin. 2007, 27, 1524–1530. [Google Scholar]
- Xia, H.Y.; Yang, Y.; Yang, T.T.; Xia, L.H.; Wang, R.Z. Correlation between characteristics of tannin cells and total phenolics and condensed tannins contents in persimmon fruits. Sci. Silvae Sin. 2014, 50, 164–172. [Google Scholar]
- Zhang, N. Germplasm innovation of Chinese PCNA persimmon and preliminary evalution of economical traits. Master’s Thesis, Huazhong Agricultural University, Wuhan, China, 2017. [Google Scholar]
- Zhang, J.S.; Sobajima, Y.; Ishida, M. The observation of tannin cells of different persimmon varieties in the middle-stage of fruit development. Acta Univ. Agric. Boreali-Occident. 1993, 21, 41–45. [Google Scholar]
- Ikegami, A.; Yonemori, K.; Sugiura, A.; Sato, A.; Yamada, M. Segregation of astringency in F1 progenies derived from crosses between pollination-constant, nonastringent persimmon cultivars. HortScience 2004, 39, 371–374. [Google Scholar] [CrossRef]
- Ito, S.; Oshima, Y. Studies on the tannin of Japanese persimmon (Diospyros kaki L.) Part I. Isolation of leucoanthocyanin from kaki fruit. Agric. Biol. Chem. 1962, 26, 156–161. [Google Scholar] [CrossRef]
- Gu, H.F.; Li, C.M.; Xu, Y.J.; Hu, W.F.; Chen, M.H.; Wan, Q.H. Structural features and antioxidant activity of tannin from persimmon pulp. Food Res. Int. 2008, 41, 208–217. [Google Scholar] [CrossRef]
- Akagi, T.; Ikegami, A.; Suzuki, Y.; Yoshida, J.; Yamada, M.; Sato, A.; Yonemori, K. Expression balances of structural genes in shikimate and flavonoid biosynthesis cause a difference in proanthocyanidin accumulation in persimmon (Diospyros kaki Thunb.) fruit. Planta 2009, 230, 899–915. [Google Scholar] [CrossRef]
- Mo, R.L. Isolation and characterization of ADH and PDC genes involved in natural loss of astringency in Chinese PCNA Persimmon. Ph.D. Thesis, Huazhong Agricuture University, Wuhan, China, 2015. [Google Scholar]
- Tanaka, T.; Takahashi, R.; Kouno, I.; Nonaka, G. Chemical evidence for the de-astringency (insolubilization of tannins) of persimmon fruit. J. Chem. Soc. Perkin Trans. 1 1994, 20, 3013–3022. [Google Scholar] [CrossRef]
- Akagi, T.; Tsujimoto, T.; Ikegami, A.; Yonemori, K. Effects of seasonal temperature changes on DkMyb4 expression involved in proanthocyanidin regulation in two genotypes of persimmon (Diospyros kaki Thunb.) fruit. Planta 2011, 233, 883–894. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wu, X.; Sun, C.F.; Chen, W.X.; Zhang, M.; Liu, N.N.; Zhang, Q.L.; Xu, L.Q.; Luo, Z.R. Preferential transport activity of DkDTX5/MATE5 affects the formation of different astringency in persimmon. J. Integr. Plant Biol. 2023, 65, 2304–2319. [Google Scholar] [CrossRef] [PubMed]
- Li, J.W. Functional characterization of DkMYB15 in proanthocyanidin biosynthesis in Chineses PCNA persimmon. Master’s Thesis, Huazhong Agricuture University, Wuhan, China, 2020. [Google Scholar]
- Zheng, Q.Y. Identification of key genes in proanthocyanidin metabolism and functional characterization of DkMYB21 in Chinese PCNA persimmon. Ph.D. Thesis, Huazhong Agricuture University, Wuhan, China, 2022. [Google Scholar]
- Luo, Y.J. Identification and characterization of microRNAs from Chinese PCNA persimmon based on high throughput sequencing. Master’s Thesis, Huazhong Agricuture University, Wuhan, China, 2014. [Google Scholar]
- Zhang, M. Microrna143c mediated natural de-astringency in Chinese PCNA persimmon. Ph.D. Thesis, Huazhong Agricuture University, Wuhan, China, 2022. [Google Scholar]
- Zaman, F. DkmiR397 regulates proanthocyanidin biosynthesis via negative modulating DkLAC2 in Chinese PCNA persimmon. Ph.D. Thesis, Huazhong Agricuture University, Wuhan, China, 2022. [Google Scholar]
- Zhang, M.; Zhang, P.X.; Xu, L.Q.; Guo, D.Y.; Luo, Z.R.; Zhang, Q.L. Research progress of sex determination and sex-linked markers in persimmon. J. Fruit Sci. 2018, 35, 610–619. [Google Scholar] [CrossRef]
- Zhang, P.X. Investigation of sexuality and application of sex-linked markers in Diospyros L. Master’s Thesis, Huazhong Agricuture University, Wuhan, China, 2016. [Google Scholar]
- Huang, Y.F. The analysis of taxonomic status and application value as pollen donor of some androecious genotypes native to China. Master’s Thesis, Huazhong Agricultural University, Wuhan, China, 2013. [Google Scholar]
- Zhang, Q.L.; Luo, Z.R. Observation of giant pollen and pollen germination ability in vitro of some Diospyros spp. and their pollen germination on the stigma of Diospyros kaki cv. Luotiantianshi. J. Fruit Sci. 2006, 23, 293–296. [Google Scholar] [CrossRef]
- Xu, L.Q. Studies on the biological characters and new germplasm creation of some staminate germplasm in Diospyros L. native to China. Ph.D. Thesis, Huazhong Agricultural University, Wuhan, China, 2008. [Google Scholar]
- Yi, H.L.; Deng, X.X.; Fu, C.H. Application of embryo rescue techniques in fruit crops. J. Fruit Sci. 2001, 18, 224–228. [Google Scholar]
- Leng, P.; Yamamura, H. Fruit set and embryo rescue in crosses using parthenocarpic Mopanshi persimmon. Sci. Hortic. 2006, 107, 332–336. [Google Scholar] [CrossRef]
- Xu, L.Q.; Zhang, Q.L.; Luo, Z.R. Effect of embryo development stage on embryo rescue for Jirou persimmon culitivar. J. Fruit Sci. 2008, 25, 45–48. [Google Scholar]
- Liu, Z. Establishment of genetic improvement technical system for astringent persimmon (Diospyros kaki Thunb.) cultivars to become non-astringent. Master’s Thesis, Huazhong Agricultural University, Wuhan, China, 2016. [Google Scholar]
- Liu, H.F.; Duan, C.G.; Chen, X.S.; Feng, B.C.; Yang, H.H. Application of embryo rescue techniques in the breeding of kernelled fruit trees. Biotechnol. Bull. 2003, 3, 12–16. [Google Scholar]
- Ikegami, A.; Eguchi, S.; Akagi, T.; Sato, A.; Yamada, M.; Kanzaki, S.; Kitajima, A.; Yonemori, K. Development of molecular markers linked to the allele associated with the non-astringent trait of the Chinese persimmon (Diospyros kaki Thunb.). J. Jpn. Soc. Hort. Sci. 2011, 80, 150–155. [Google Scholar] [CrossRef]
- Pei, X. Early screening of natural deastringency trait and cross breeding in Chinese pollination constant non-astringent persimmon. Ph.D. Thesis, Huazhong Agricuture University, Wuhan, China, 2013. [Google Scholar]
- Huang, J.M. Improvement of genetic modification system and innovative germplasm characterization in Chinese PCNA persimmon. Master’s Thesis, Huazhong Agricultural University, Wuhan, China, 2016. [Google Scholar]
- Liu, Y.Y.; Ma, J.L.; Tang, X.; Song, C.L. Studies on adventitious shoot regeneration from stems without internode of ‘Mopan’ persimmon tube seedlings. J. Anhui Agric. Sci. 2006, 34, 2115–2117. [Google Scholar] [CrossRef]
- Wang, Y.; Qiu, X.; Shun, H.L.; Chang, H.; Xu, L.Q. Study on tissue culture and rapid propagation of persimmon from deyang, a new species of persimmon. J. Anhui Agric. Sci. 2020, 48, 51–55. [Google Scholar] [CrossRef]
- Qin, H.L.; Gao, Y.Y.; Huang, L.H.; Chen, S.Y.; Huang, T.K.; Li, X.G.; Zhou, X.X.; Guan, C.F.; Yang, Y. Study on tissue culture and rapid propagation of Xiaoguo Tianshi as a sweet persimmon rootstock. J. Fruit Sci. 2023, 40, 1992–2000. [Google Scholar] [CrossRef]
- Liu, B.; Du, X.Y.; Chen, W.X.; Guo, D.Y.; Xu, L.Q.; Zhang, Q.L.; Luo, Z.R. Establishment and optimization of in vitro rapid propagation technology of Xiaoguo Tianshi as compatible rootstock for persimmon. J. Fruit Sci. 2023, 40, 1980–1991. [Google Scholar] [CrossRef]
- Tao, R.; Handa, T.; Tamura, M.; Sugiura, A. Genetic Transformation of Japanese Persimmon by Agrobacterium rhizogenes Wild Type Strain A4. J. Jpn. Soc. Hortic. Sci. 1994, 63, 283–289. Available online: https://www.jstage.jst.go.jp/article/jjshs1925/63/2/63_2_283/_pdf/-char/ja (accessed on 15 July 2023). [CrossRef]
- Nakamura, Y.; Kobayashi, S.; Nakajima, I. Agrobacterium-mediated transformation and plant regeneration from hypocotyl segments of Japanese persimmon (Diospyros kaki Thunb). Plant Cell Rep. 1998, 17, 435–440. [Google Scholar] [CrossRef] [PubMed]
- Tao, R.; Dandekar, A.M.; Uratsu, S.L.; Vail, P.V.; Tebbts, J.S. Engineering genetic resistance against insects in Japanese persimmon using the cryIA(c) gene of Bacillus thuringiensis. J. Am. Soc. Hortic. Sci. 1997, 122, 764–771. [Google Scholar] [CrossRef]
- Gao, M.; Sakamoto, A.; Miura, K.; Murata, N.; Sugiura, A.; Tao, R. Transformation of Japanese persimmon (Diospyros kaki Thunb.) with a bacterial gene for choline oxidase. Mol. Breed. 2000, 6, 501–510. [Google Scholar] [CrossRef]
- Gao, M.; Tao, R.; Miura, K.; Dandekar, A.M.; Sugiura, A. Transformation of Japanese persimmon (Diospyros kaki Thunb.) with apple cDNA encoding NADP-dependent sorbitol-6-phosphate dehydrogenase. Plant Sci. 2001, 160, 837–845. [Google Scholar] [CrossRef]
- Koshita, Y.; Nakamura, Y.; Kobayashi, S.; Morinaga, K. Introduction of the rolC gene into the genome of the Japanese persimmon causes dwarfism. J. Jpn. Soc. Hortic. Sci. 2002, 71, 529–531. [Google Scholar] [CrossRef]
- Gao, M.; Takeishi, H.; Katayama, A.; Tao, R. Genetic transformation of Japanese persimmon with Flowering Locus T (FT) gene and TERMINAL FLOWER 1 (TFL1) homologues gene. Acta Hort. 2013, 996, 159–164. [Google Scholar] [CrossRef]
- Yin, H.; Song, C.L.; Ma, J.L.; Zhang, Z.D.; Tang, X. Optimization of Agrobacterium tumefaciens mediated genetic transformation system of Mopanshi. Acta Agric. Boreali-Sin. 2007, 22, 56–59. [Google Scholar]
- Li, X.H.; Jiang, Z.Y.; Shen, Y.Y.; Li, F.H.; Yu, X.Y.; Qu, S.C. In vitro regeneration and Agrobacterium tumefaciens-mediated genetic transformation of D. lotus (Diospyros lotus L.). Sci. Hortic. 2018, 236, 229–237. [Google Scholar] [CrossRef]
- Zhang, M.; Yang, S.C.; Chen, W.X.; Xu, L.Q.; Guo, D.Y.; Luo, Z.R.; Zhang, Q.L. An efficient Agrobacterium-mediated genetic transformation system for persimmon (Diospyros kaki Thunb.). Horticulturae 2022, 8, 422. [Google Scholar] [CrossRef]
- Wang, Y. The mechanism of natural astringency loss in Chinese pollination—Constant and non-astringent persimmons. Ph.D. Thesis, Huazhong Agricultural University, Wuhan, China, 2010. [Google Scholar]
- Lepiniec, L.; Debeaujon, I.; Routaboul, J.M.; Baudry, A.; Pourcel, L.; Nesi, N.; Caboche, M. Genetics and biochemistry of seed flavonoids. Annu. Rev. Plant Biol. 2006, 57, 405–430. [Google Scholar] [CrossRef]
- Yang, L.Y. Expression analysis and functional characterization of proanthocyanidin precursor transmembrane gene DkGST1 in persimmon fruit. Master’s Thesis, Huazhong Agricultural University, Wuhan, China, 2018. [Google Scholar]
- Deng, J. Characterization of proanthocyanidin precursor transport related gene DkGSTF1 in persimmon. Master’s Thesis, Huazhong Agricultural University, Wuhan, China, 2021. [Google Scholar]
- Pang, Y.Z.; Peel, G.J.; Sharma, S.B.; Tang, Y.H.; Dixon, R.A. A transcript profiling approach reveals an epicatechin-specific glucosyltransferase expressed in the seed coat of Medicago truncatula. Proc. Natl. Acad. Sci. USA 2008, 105, 14210–14215. [Google Scholar] [CrossRef]
- Wang, S.D. Cloning and function characterization of proanthocyanidin precursor transporting gene DkAHA1 in persimmon. Master’s Thesis, Huazhong Agricultural University, Wuhan, China, 2020. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Classification | Representative Varieties | Origin Area | Deastringency Method | Deastringency Period | Genetic Characteristics | Key Transcript Factor |
|---|---|---|---|---|---|---|
| C-PCNA | Eshi 1 Luotian Tianshi | China | Dilution effect and coagulation effect | 25 WAB | Quality traits/ Dominant [6] | DkMYB14 [7] |
| J-PCNA | Youhou | Japan | Dilution effect | 15 WAB | Quality traits/ Recessive [6] | DkMYB4 [8] |
| PVNA | Huangjin Fangshi | China | Partially deastringent | Not fully natural deastringency | Quantitative traits [6] | - |
| PCA | Huashi 1 | China | Partially deastringent | Not fully natural deastringency | Quantitative traits [6] | - |
| PVA | Mopanshi Gongcheng Shuishi | China | Not natural deastringency | Not natural deastringency | Quantitative traits [6] | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, S.; Zhang, M.; Xu, L.; Zhang, Q.; Zhou, C.; Hu, X.; Luo, Z. Recent Advances in Natural Deastringency and Genetic Improvement of Chinese PCNA Persimmon (Diospyros kaki). Horticulturae 2023, 9, 1273. https://doi.org/10.3390/horticulturae9121273
Yang S, Zhang M, Xu L, Zhang Q, Zhou C, Hu X, Luo Z. Recent Advances in Natural Deastringency and Genetic Improvement of Chinese PCNA Persimmon (Diospyros kaki). Horticulturae. 2023; 9(12):1273. https://doi.org/10.3390/horticulturae9121273
Chicago/Turabian StyleYang, Sichao, Meng Zhang, Lei Xu, Qinglin Zhang, Chaohua Zhou, Xinlong Hu, and Zhengrong Luo. 2023. "Recent Advances in Natural Deastringency and Genetic Improvement of Chinese PCNA Persimmon (Diospyros kaki)" Horticulturae 9, no. 12: 1273. https://doi.org/10.3390/horticulturae9121273