
The Endophytic Entomopathogenic Fungus Beauveria bassiana Alleviates Adverse Effects of Salt Stress in Potato Plants
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. A Fungal Isolate and Its Cultivation
2.2. Plant Cultivation and Experimental Design
2.3. Plant Colonization Assay
2.4. Plant Growth Parameters and Leaf Contents of Photosynthetic Pigments
2.5. Lipid Peroxidation
2.6. Activity of Antioxidant Enzymes in the Leaves
2.7. The Free-Proline Content of Plants
2.8. Statistical Analyses
3. Results
3.1. Plant Colonization by the Fungus
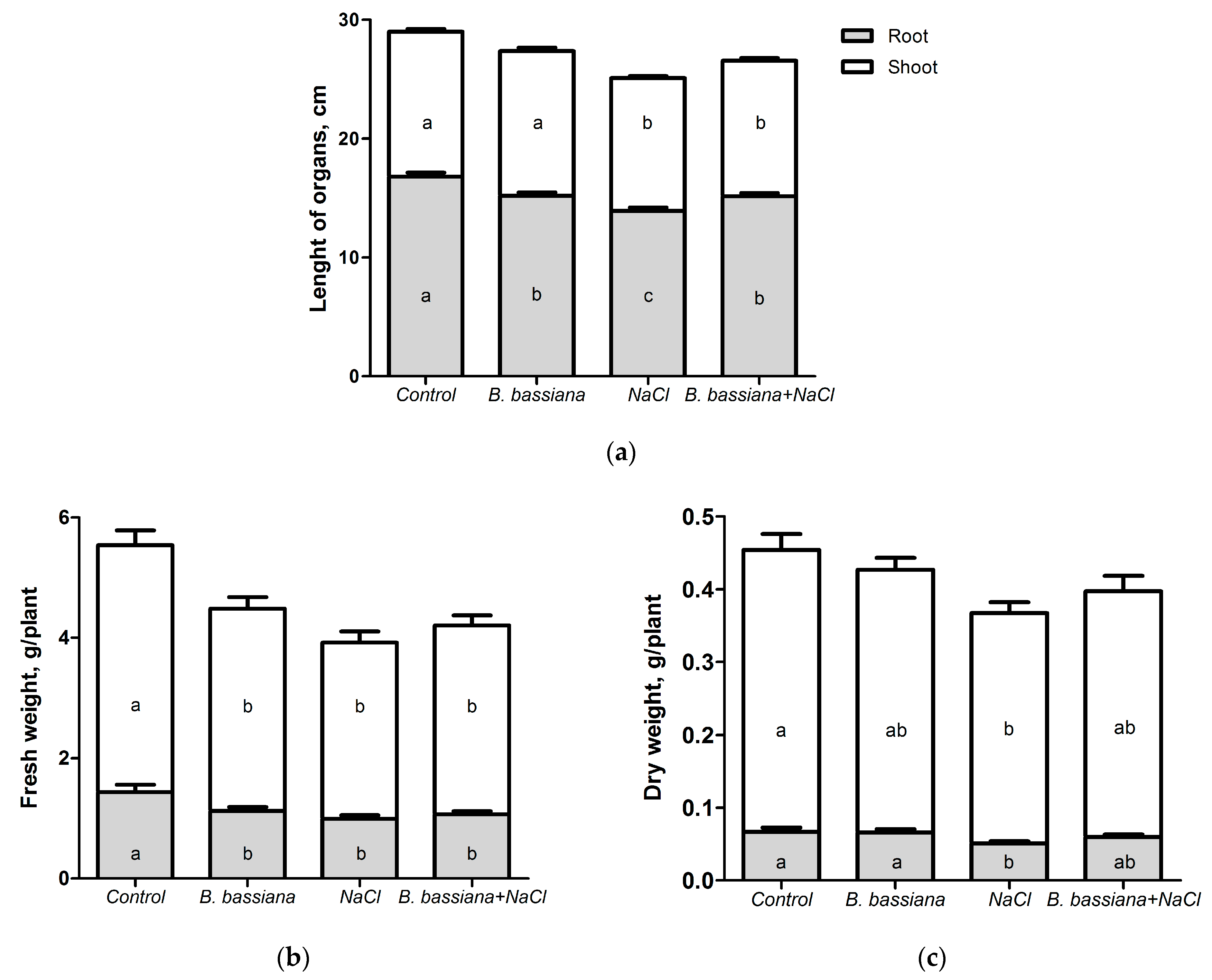
3.2. Effects of B. bassiana and Salt on Plant Growth
3.3. Effects of B. bassiana and Salt on the Photosynthetic Pigment Content
3.4. The Impact of B. bassiana and Salt on Lipid Peroxidation
3.5. Effects of B. bassiana and Salt on the Activity of Antioxidant Enzymes
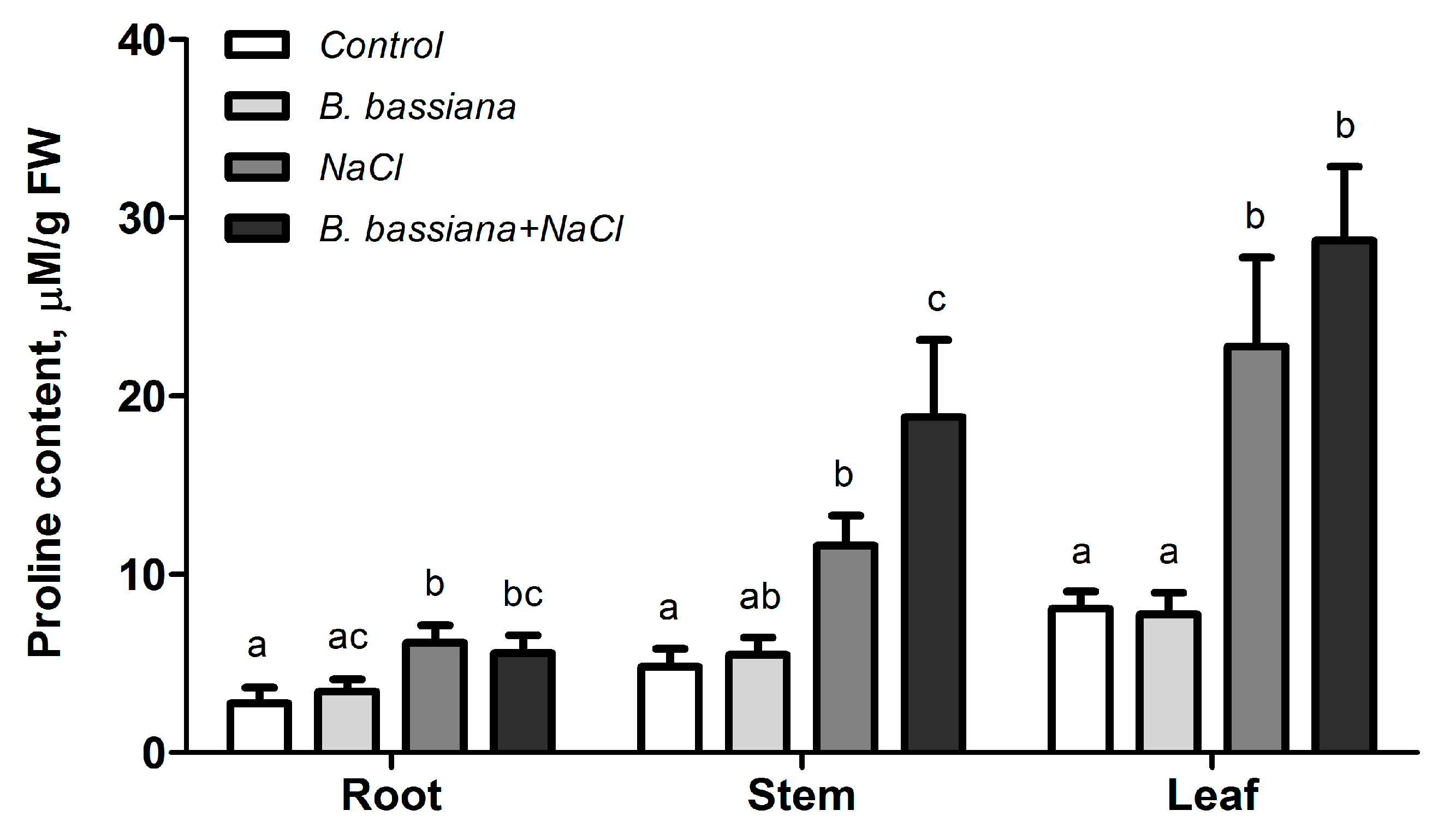
3.6. Effects of B. bassiana and Salt on Proline Accumulation in Different Parts of Plants
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

References
- Liang, W.; Ma, X.; Wan, P.; Liu, L. Plant salt-tolerance mechanism: A review. Biochem. Biophys. Res. Commun. 2018, 495, 286–291. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Guo, Y. Elucidating the molecular mechanisms mediating plant salt-stress responses. New Phytol. 2018, 217, 523–539. [Google Scholar] [CrossRef] [PubMed]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed]
- Shahid, S.A.; Rahman, K. Soil salinity development, classification, assessment, and management in irrigated agriculture. In Handbook of Plant and Crop Stress; Pessarakli, M., Ed.; CRC Press, Taylor and Francis Group: Boca Raton, FL, USA, 2011; pp. 23–38. [Google Scholar]
- Zhang, J.L.; Shi, H. Physiological and molecular mechanisms of plant salt tolerance. Photosynth. Res. 2013, 115, 1–22. [Google Scholar] [CrossRef]
- Gupta, B.; Huang, B. Mechanism of salinity tolerance in plants: Physiological, biochemical, and molecular characterization. Int. J. Genom. 2014, 2014, 701596. [Google Scholar] [CrossRef]
- Chawla, S.; Jain, S.; Jain, V.J. Salinity induced oxidative stress and antioxidant system in salt-tolerant and saltsensitive cultivars of rice (Oryza sativa L.). Plant Biochem. Biotechnol. 2013, 22, 27–34. [Google Scholar] [CrossRef]
- Schmitt, F.J.; Renger, G.; Friedrich, T.; Kreslavski, V.D.; Zharmukhamedov, S.K.; Los, D.A.; Kuznetsov, V.V.; Allakhverdiev, S.I. Reactive oxygen species: Re-evaluation of generation, monitoring and role in stress-signaling in phototrophic organisms. Biochim. Biophys. Acta 2014, 1837, 835–848. [Google Scholar] [CrossRef]
- Gao, H.-J.; Yang, H.-Y.; Bai, J.-P.; Liang, X.-Y.; Lou, Y.; Zhang, J.-L.; Wang, D.; Zhang, J.-L.; Niu, S.-Q.; Chen, Y.-L. Ultrastructural and physiological responses of potato (Solanum tuberosum L.) plantlets to gradient saline stress. Front. Plant Sci. 2015, 5, 787. [Google Scholar] [CrossRef]
- FAO Statistical Yearbook. World Food and Agricultural. Rome: Food and Agricultural Organization for the United Nations, 2022. [Electronic Resource]. Available online: https://www.fao.org/documents/card/en/c/cc2211en (accessed on 15 July 2023).
- Jaarsma, R.; de Boer, A.H. Salinity tolerance of two potato cultivars (Solanum tuberosum) correlates with differences in vacuolar transport activity. Front. Plant Sci. 2018, 9, 737. [Google Scholar] [CrossRef]
- Jaarsma, R.; de Vries, R.S.M.; de Boer, A.H. Effect of salt stress on growth, Na+ accumulation and proline metabolism in potato (Solanum tuberosum) cultivars. PLoS ONE 2013, 8, e60183. [Google Scholar] [CrossRef]
- Faried, H.F.; Ayyub, C.M.; Amjad, M.; Ahmed, R. Salinity impacts ionic, physiological and biochemical attributes in potato. Pak. J. Agric. Sci. 2016, 53, 17–25. [Google Scholar] [CrossRef]
- Chourasia, K.N.; Lal, M.K.; Tiwari, R.K.; Dev, D.; Kardile, H.B.; Patil, V.U.; Kumar, A.; Vanishree, G.; Kumar, D.; Bhardwaj, V.; et al. Salinity stress in potato: Understanding physiological, biochemical and molecular responses. Life 2021, 11, 545. [Google Scholar] [CrossRef] [PubMed]
- Schubert, S.; Neubert, A.; Schierholt, A.; Sümer, A.; Zörb, C. Development of salt-resistant maize hybrids: The combination of physiological strategies using conventional breeding methods. Plant Sci. 2009, 177, 196–202. [Google Scholar] [CrossRef]
- Afzal, M.; Hindawi, S.E.S.; Alghamdi, S.S.; Migdadi, H.H.; Khan, M.A.; Hasnain, M.U.; Arslan, M.; Habib ur Rahman, M.; Sohaib, M. Potential breeding strategies for improving salt tolerance in crop plants. J. Plant Growth Regul. 2023, 42, 3365–3387. [Google Scholar] [CrossRef]
- Vikram; Pooja; Sharma, J.; Sharma, A. Role of brassinosteroids in plants responses to salinity stress: A review. JANS 2022, 14, 582–599. [Google Scholar] [CrossRef]
- Efimova, M.V.; Khripach, V.A.; Boyko, E.V.; Malofii, M.K.; Kolomeichuk, L.V.; Murgan, O.K.; Vidershpun, A.N.; Mukhamatdinova, E.A.; Kuznetsov, V.V. The priming of potato plants induced by brassinosteroids reduces oxidative stress and increases salt tolerance. Dokl. Biol. Sci. 2018, 478, 33–36. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Yang, R.; Zhang, X.; Wang, Q.; Wang, B.; Zheng, X.; Li, Y.; Prusky, D.; Bi, Y. Brassinosteroid accelerates wound healing of potato tubers by activation of reactive oxygen metabolism and phenylpropanoid metabolism. Foods 2022, 11, 906. [Google Scholar] [CrossRef]
- Hu, Y.; Xia, S.; Su, Y.; Wang, H.; Luo, W.; Su, S.; Xiao, L. Brassinolide increases potato root growth in vitro in a dose-dependent way and alleviates salinity stress. Biomed Res. Int. 2016, 2016, 8231873. [Google Scholar] [CrossRef]
- Dodd, I.C.; Perez-Alfocea, F. Microbial amelioration of crop salinity stress. J. Exp. Bot. 2012, 63, 3415–3428. [Google Scholar] [CrossRef]
- Zhang, Y.; Yu, X.; Zhang, W.; Lang, D.; Zhang, X.; Cui, G.; Zhang, X. Interactions between endophytes and plants: Beneficial effect of endophytes to ameliorate biotic and abiotic stresses in plants. J. Plant Biol. 2019, 62, 1–13. [Google Scholar] [CrossRef]
- Kamran, M.; Imran, Q.M.; Ahmed, M.B.; Falak, N.; Khatoon, A.; Yun, B.-W. Endophyte-mediated stress tolerance in plants: A sustainable strategy to enhance resilience and assist crop improvement. Cells 2022, 11, 3292. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Schillaci, M.; Walker, R.; Smith, P.M.C.; Watt, M.; Roessner, U. Alleviation of salinity stress in plants by endophytic plant-fungal symbiosis: Current knowledge, perspectives and future directions. Plant Soil 2021, 461, 219–244. [Google Scholar] [CrossRef]
- Zheng, H.; Qiao, M.; Lv, Y.; Du, X.; Zhang, K.-Q.; Yu, Z. New Species of Trichoderma isolated as endophytes and saprobes from southwest China. J. Fungi 2021, 7, 467. [Google Scholar] [CrossRef] [PubMed]
- Manjunatha, N.; Manjunatha, N.; Li, H.; Sivasithamparam, K.; Jones, M.G.K.; Edwards, I.; Wylie, S.J.; Agarrwal, R. Fungal endophytes from salt-adapted plants confer salt tolerance and promote growth in wheat (Triticum aestivum L.) at early seedling stage. Microbiology 2022, 168, 001225. [Google Scholar] [CrossRef]
- Bouzouina, M.; Kouadria, R.; Lotmani, B. Fungal endophytes alleviate salt stress in wheat in terms of growth, ion homeostasis and osmoregulation. J. Appl. Microbiol. 2021, 130, 913–925. [Google Scholar] [CrossRef]
- Hosseyni Moghaddam, M.S.; Safaie, N.; Soltani, J.; Hagh-Doust, N. Desert-adapted fungal endophytes induce salinity and drought stress resistance in model crops. Plant Physiol. Biochem. 2021, 160, 225–238. [Google Scholar] [CrossRef]
- Bamisile, B.S.; Dash, C.K.; Akutse, K.S.; Keppanan, R.; Wang, L. Fungal endophytes: Beyond herbivore management. Front. Microbiol. 2018, 9, 544. [Google Scholar] [CrossRef]
- Vega, F.E. The use of fungal entomopathogens as endophytes in biological control: A review. Mycologia 2018, 110, 4–30. [Google Scholar] [CrossRef]
- Jaber, L.R.; Ownley, B.H. Can we use entomopathogenic fungi as endophytes for dual biological control of insect pests and plant pathogens? Biol. Control 2018, 116, 36–45. [Google Scholar] [CrossRef]
- Liao, X.; Lovett, B.; Fang, W.; St Leger, R.J. Metarhizium robertsii produces indole-3-acetic acid, which promotes root growth in Arabidopsis and enhances virulence to insects. Microbiology 2017, 163, 980–991. [Google Scholar] [CrossRef]
- Lozano-Tovar, M.D.; Garrido-Jurado, I.; Quesada-Moraga, E.; Raya-Ortega, M.C.; Trapero-Casas, A. Metarhizium brunneum and Beauveria bassiana release secondary metabolites with antagonistic activity against Verticillium dahliae and Phytophthora megasperma olive pathogens. Crop Prot. 2017, 100, 186–195. [Google Scholar] [CrossRef]
- Jaber, L.R.; Araj, S.E. Interactions among endophytic fungal entomopathogens (Ascomycota: Hypocreales), the green peach aphid Myzus persicae Sulzer (Homoptera: Aphididae), and the aphid endoparasitoid Aphidius colemani Viereck (Hymenoptera: Braconidae). Biol. Control 2018, 116, 53–61. [Google Scholar] [CrossRef]
- Raad, M.; Glare, T.R.; Brochero, H.L.; Müller, C.; Rostás, M. Transcriptional reprogramming of Arabidopsis thaliana defence pathways by the entomopathogen Beauveria bassiana correlates with resistance against a fungal pathogen but not against insects hormones, plant–microbe interaction, Plutella xylostella, Myzus persicae, Sclerotinia sclerotiorum. Front. Microbiol. 2019, 10, 615. [Google Scholar] [CrossRef]
- Mantzoukas, S.; Eliopoulos, P.A. Endophytic entomopathogenic fungi: A valuable biological control tool against plant pests. Appl. Sci. 2020, 10, 360. [Google Scholar] [CrossRef]
- Krell, V.; Unger, S.; Jakobs-Schoenwandt, D.; Patel, A.V. Endophytic Metarhizium brunneum mitigates nutrient deficits in potato and improves plant productivity and vitality. Fungal Ecol. 2018, 34, 43–49. [Google Scholar] [CrossRef]
- Kryukov, V.Y.; Kabilov, M.R.; Smirnova, N.; Tomilova, O.G.; Tyurin, M.V.; Akhanaev, Y.B.; Polenogova, O.V.; Danilov, V.P.; Zhangissina, S.K.; Alikina, T.; et al. Bacterial decomposition of insects post-Metarhizium infection: Possible influence on plant growth. Fungal Biol. 2019, 123, 927–935. [Google Scholar] [CrossRef]
- Tomilova, O.G.; Shaldyaeva, E.M.; Kryukova, N.A.; Pilipova, Y.V.; Schmidt, N.S.; Danilov, V.P.; Kryukov, V.Y.; Glupov, V.V. Entomopathogenic fungi decrease Rhizoctonia disease in potato in field conditions. PeerJ 2020, 16, e9895. [Google Scholar] [CrossRef]
- Barra-Bucarei, L.; González, M.G.; Iglesias, A.F.; Aguayo, G.S.; Peñalosa, M.G.; Vera, P.V. Beauveria bassiana multifunction as an endophyte: Growth promotion and biologic control of Trialeurodes vaporariorum, (Westwood) (Hemiptera: Aleyrodidae) in tomato. Insects 2020, 11, 591. [Google Scholar] [CrossRef]
- Barra-Bucarei, L.; Iglesias, A.F.; González, M.G.; Aguayo, G.S.; Carrasco-Fernández, J.; Castro, J.F.; Campos, J.O. Antifungal activity of Beauveria bassiana endophyte against Botrytis cinerea in two Solanaceae crops. Microorganisms 2020, 8, 65. [Google Scholar] [CrossRef]
- Khan, A.L.; Hamayun, M.; Khan, S.A.; Kang, S.-M.; Shinwari, Z.K.; Kamran, M.; ur Rehman, S.; Kim, J.-G.; Lee, I.-J. Pure culture of Metarhizium anisopliae LHL07 reprograms soybean to higher growth and mitigates salt stress. World J. Microbiol. Biotechnol. 2012, 28, 1483–1494. [Google Scholar] [CrossRef]
- Chaudhary, P.J.; Raghunandan, B.L.; Patel, H.K.; Mehta, P.V.; Patel, N.B.; Sonth, B.; Dave, A.; Bagul, S.Y.; Divya, M.; Jain, D.; et al. Plant growth-promoting potential of entomopathogenic fungus Metarhizium pinghaense AAUBC-M26 under elevated salt stress in tomato. Agronomy 2023, 13, 1577. [Google Scholar] [CrossRef]
- Kuzhuppillymyal-Prabhakarankutty, L.; Tamez-Guerra, P.; Gomez-Flores, R.; Rodriguez-Padilla, M.C.; Ek-Ramos, M.J. Endophytic Beauveria bassiana promotes drought tolerance and early flowering in corn. World J. Microbiol. Biotechnol. 2020, 36, 47. [Google Scholar] [CrossRef] [PubMed]
- Dara, S.; Dara, S.; Dara, S. Impact of entomopathogenic fungi on the growth, development, and health of cabbage growing under water stress. Am. J. Plant Sci. 2017, 8, 1224–1233. [Google Scholar] [CrossRef]
- Gana, L.P.; Etsassala, N.G.E.R.; Nchu, F. Interactive effects of water deficiency and endophytic Beauveria bassiana on plant growth, nutrient uptake, secondary metabolite contents, and antioxidant activity of Allium cepa L. J. Fungi 2022, 8, 874. [Google Scholar] [CrossRef] [PubMed]
- Akter, T.; Mimma, A.A.; Haque, M.A.; Hossain, M.M.; Ghosh, T.K.; Zinan, N.; Chowdhury, M.Z.H.; Islam, S.M.N. Seed priming with Beauveria bassiana improves growth and salt stress response in rice. Environ. Exp. Bot. 2023, 213, 105427. [Google Scholar] [CrossRef]
- Danilova, E.D.; Medvedeva, Y.V.; Efimova, M.V. The effect of chloride salinity on growth and physiological processes in mid-ripening varieties of Solanum tuberosum plants. Vestnik Tomskogo gosudarstvennogo universiteta. Biologiya Tomsk State University J. Biol. 2018, 44, 158–171. [Google Scholar] [CrossRef]
- Tomilova, O.G.; Kryukova, N.A.; Efimova, M.V.; Kovtun, I.S.; Kolomeichuk, L.V.; Kryukov, V.Y.; Glupov, V.V. Early physiological response of potato plants to entomopathogenic fungi under hydroponic conditions. Horticulturae 2021, 7, 217. [Google Scholar] [CrossRef]
- Lednev, G.; Levchenko, M.; Kazartsev, I. Members of the genus Beauveria associated with natural populations of locusts in southern european Russia. Diversity 2023, 15, 930. [Google Scholar] [CrossRef]
- Posada, F.; Aime, M.C.; Peterson, S.W.; Rehner, S.A.; Vega, F.E. Inoculation of coffee plants with the fungal entomopathogen Beauveria bassiana (Ascomycota: Hypocreales). Mycol. Res. 2007, 111, 748–757. [Google Scholar] [CrossRef]
- McKinnon, A.C.; Saari, S.; Moran-Diez, M.E.; Meyling, N.V.; Raad, M.; Glare, T.R. Beauveria bassiana as an endophyte: A critical review on associated methodology and biocontrol potential. BioControl 2017, 62, 1–17. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar] [CrossRef]
- Buege, J.A.; Aust, S.D. Microsomal lipid peroxidation. Methods Enzymol. 1978, 52, 302–310. [Google Scholar] [CrossRef] [PubMed]
- Beauchamp, C.; Fridovich, I. Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels. Anal. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef]
- Shevyakova, N.I.; Stetsenko, L.A.; Meshcheryakov, A.B.; Kuznetsov, V.V. The activity of the peroxidase system in the course of stress-induced CAM development. Russian J. Plant Physiol. 2002, 49, 598–604. [Google Scholar] [CrossRef]
- Esen, A. A simple method for quantitative, semiquantitative, and qualitive assay of protein. Anal. Biochem. 1978, 89, 264–327. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Cazorla Perfetti, D.J.; Morales Moreno, P.; Acosta Quintero, M.E. Effects of thermal, saline and pH gradients on in vitro germination of a native isolate of Beauveria bassiana (Balsamo) Vuillemin, pathogenic to Rhodnius prolixus and Triatoma maculata. Rev. Cient. 2007, 17, 627–631. [Google Scholar]
- Lubna; Asaf, S.; Hamayun, M.; Khan, A.L.; Waqas, M.; Khan, M.A.; Jan, R.; Lee, I.-J.; Hussain, A. Salt tolerance of Glycine max L induced by endophytic fungus Aspergillus flavus CSH1, via regulating its endogenous hormones and antioxidative system. Plant Physiol. Biochem. 2018, 128, 13–23. [Google Scholar] [CrossRef]
- Rouphael, Y.; Colla, G.; Giordano, M.; Raimondi, G.; Pannico, A.; Di Stasio, E.; Cardarelli, M.; Bonini, P.; De Pascale, S. Endophytic fungi induce salt stress tolerance in greenhouse-grown basil. Acta Hortic. 2020, 1268, 125–132. [Google Scholar] [CrossRef]
- Ali, R.; Gul, H.; Rauf, M.; Arif, M.; Hamayun, M.; Husna; Khilji, S.A.; Ud-Din, A.; Sajid, Z.A.; Lee, I.J. Growth-promoting endophytic fungus (Stemphylium lycopersici) ameliorates salt stress tolerance in maize by balancing ionic and metabolic status. Front. Plant Sci. 2022, 13, 890565. [Google Scholar] [CrossRef]
- Siddiqui, Z.S.; Wei, X.; Umar, M.; Abideen, Z.; Zulfiqar, F.; Chen, J.; Hanif, A.; Dawar, S.; Dias, D.A.; Yasmeen, R. Scrutinizing the application of saline endophyte to enhance salt tolerance in rice and maize plants. Front. Plant Sci. 2022, 12, 770084. [Google Scholar] [CrossRef] [PubMed]
- Jan, F.G.; Hamayun, M.; Hussain, A.; Jan, G.; Ali, S.; Khan, S.A.; Lee, I.-J. Endophytic Candida membranifaciens from Euphorbia milii L. alleviate salt stress damages in maize. Agronomy 2022, 12, 2263. [Google Scholar] [CrossRef]
- Morsy, M.; Cleckler, B.; Armuelles-Millican, H. Fungal endophytes promote tomato growth and enhance drought and salt tolerance. Plants 2020, 9, 877. [Google Scholar] [CrossRef] [PubMed]
- Azad, K.; Kaminskyj, S. A fungal endophyte strategy for mitigating the effect of salt and drought stress on plant growth. Symbiosis 2016, 1, 73–78. [Google Scholar] [CrossRef]
- Khan, A.L.; Waqas, M.; Khan, A.R.; Hussain, J.; Kang, S.-M.; Gilani, S.A.; Lee, I.-J. Fungal endophyte Penicillium janthinellum LK5 improves growth of ABA-deficient tomato under salinity. World J. Microbiol. Biotechnol. 2013, 29, 2133–2144. [Google Scholar] [CrossRef] [PubMed]
- Abdelaziz, M.E.; Abdelsattar, M.; Abdeldaym, E.A.; Atia, M.A.M.; Mahmoud, A.W.M.; Saad, M.M.; Hirt, H. Piriformospora indica alters Na+/K+ homeostasis, antioxidant enzymes and LeNHX1 expression of greenhouse tomato grown under salt stress. Sci. Hortic. 2019, 256, 108532. [Google Scholar] [CrossRef]
- Ghorbani, A.; Razavi, S.M.; Ghasemi Omran, V.O.; Pirdashti, H. Piriformospora indica inoculation alleviates the adverse effect of NaCl stress on growth, gas exchange and chlorophyll fluorescence in tomato (Solanum lycopersicum L.). Plant Biol. 2018, 20, 729–736. [Google Scholar] [CrossRef]
- Khan, A.L.; Hamayun, M.; Kang, S.M.; Kim, Y.H.; Jung, H.Y.; Lee, J.H.; Lee, I.J. Endophytic fungal association via gibberellins and indole acetic acid can improve plant growth under abiotic stress: An example of Paecilomyces formosus LHL10. BMC Microbiol. 2012, 12, 3. [Google Scholar] [CrossRef]
- Efimova, M.V.; Kolomeichuk, L.V.; Boyko, E.V.; Malofii, M.K.; Vidershpan, A.N.; Plyusnin, I.N.; Golovatskaya, I.F.; Murgan, O.K.; Kuznetsov, V.V. Physiological mechanisms of Solanum tuberosum L. plants’ tolerance to chloride salinity. Russ. J. Plant Physiol. 2018, 65, 394–403. [Google Scholar] [CrossRef]
- Proietti, S.; Falconieri, G.S.; Bertini, L.; Pascale, A.; Bizzarri, E.; Morales-Sanfrutos, J.; Sabidó, E.; Ruocco, M.; Monti, M.M.; Russo, A. Beauveria bassiana rewires molecular mechanisms related to growth and defense in tomato. J. Exp. Bot. 2023, 74, 4225–4243. [Google Scholar] [CrossRef]
- Quesada Moraga, E. Entomopathogenic fungi as endophytes: Their broader contribution to IPM and crop production. Biocontrol Sci. Technol. 2020, 30, 864–877. [Google Scholar] [CrossRef]
- Bamisile, B.S.; Siddiqui, J.A.; Akutse, K.S.; Ramos Aguila, L.C.; Xu, Y. General limitations to endophytic entomopathogenic fungi use as plant growth promoters, pests and pathogens biocontrol agents. Plants 2021, 10, 2119. [Google Scholar] [CrossRef]
- Sudhir, P.; Murthy, S. Effects of salt stress on basic processes of photosynthesis. Photosynthetica 2004, 42, 481–486. [Google Scholar] [CrossRef]
- Chaves, M.M.; Flexas, J.; Pinheiro, C. Photosynthesis under drought and salt stress: Regulation mechanisms from whole plant to cell. Ann. Bot. 2009, 103, 551–560. [Google Scholar] [CrossRef] [PubMed]
- Ben Ahmed, C.; Magdich, S.; Ben Rouina, B.; Sensoy, S.; Boukhris, M.; Ben Abdullah, F. Exogenous proline effects on water relations and ions contents in leaves and roots of young olive. Amino Acids 2011, 40, 565–573. [Google Scholar] [CrossRef]
- Ma, Q.; Yue, L.J.; Zhang, J.L.; Wu, G.Q.; Bao, A.K.; Wang, S.M. Sodium chloride improves photosynthesis and water status in the succulent xerophytes Zygophyllum xanthoxylum. Tree Physiol. 2012, 32, 4–13. [Google Scholar] [CrossRef]
- Wani, A.S.; Ahmad, A.; Hayat, S.; Tahir, I. Is foliar spray of proline sufficient for mitigation of salt stress in Brassica juncea cultivars? Environ. Sci. Pollut. Res. 2016, 23, 13413–13423. [Google Scholar] [CrossRef]
- Hnilickova, H.; Kraus, K.; Vachova, P.; Hnilicka, F. Salinity stress affects photosynthesis, malondialdehyde formation, and proline content in Portulaca oleracea L. Plants 2021, 10, 845. [Google Scholar] [CrossRef]
- Kolomeichuk, L.V.; Efimova, M.V.; Zlobin, I.E.; Kreslavski, V.D.; Murgan, O.K.; Kovtun, I.S.; Khripach, V.A.; Kuznetsov, V.V.; Allakhverdiev, S.I. 24-Epibrassinolide alleviates the toxic effects of NaCl on photosynthetic processes in potato plants. Photosynth. Res. 2020, 146, 151–163. [Google Scholar] [CrossRef]
- Wydrzynski, T.J. Water splitting by photosystem II—Where do we go from here? Photosynth. Res. 2008, 98, 43–51. [Google Scholar] [CrossRef]
- Efimova, M.V.; Danilova, E.D.; Zlobin, I.E.; Kolomeichuk, L.V.; Murgan, O.K.; Boyko, E.V.; Kuznetsov, V.V. Priming potato plants with melatonin protects stolon formation under delayed salt stress by maintaining the photochemical function of photosystem II, ionic homeostasis and activating the antioxidant system. Int. J. Mol. Sci. 2023, 24, 6134. [Google Scholar] [CrossRef] [PubMed]
- Santos, C.V. Regulation of chlorophyll biosynthesis and degradation by salt stress in sunflower leaves. Sci. Hortic. 2004, 103, 93–99. [Google Scholar] [CrossRef]
- Elsheery, N.I.; Cao, K.F. Gas exchange, chlorophyll fluorescence, and osmotic adjustment in two mango cultivars under drought stress. Acta Physiol. Plant. 2008, 30, 769–777. [Google Scholar] [CrossRef]
- Jamil, M.; Bashir, S.; Anwar, S.; Bibi, S.; Bangash, A.; Ullah, F.; Rha, E.S. Effect of salinity on physiological and biochemical characteristics of different varieties of rice. Pak. J. Bot. 2012, 44, 7–13. [Google Scholar]
- Sonone, M.; Mane, A.; Sawardekar, S.; Kunkerkar, R. Consequences of salt stress on chlorophyll pigments of rice genotypes. Pharma Innov. 2023, 12, 3272–3275. [Google Scholar]
- Flowers, T.J.; Colmer, T.D. Plant salt tolerance: Adaptations in halophytes. Ann. Bot. 2015, 115, 327–331. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Yang, Z.; Liu, J.; Zhu, T.; Wei, X.; Fan, H.; Wang, B. Adaptation mechanism of salt excluders under saline conditions and its applications. Int. J. Mol. Sci. 2018, 19, 3668. [Google Scholar] [CrossRef]
- Rahman, M.M.; Mostofa, M.G.; Keya, S.S.; Siddiqui, M.N.; Ansary, M.M.U.; Das, A.K.; Rahman, M.A.; Tran, L.S.-P. Adaptive mechanisms of halophytes and their potential in improving salinity tolerance in plants. Int. J. Mol. Sci. 2021, 22, 10733. [Google Scholar] [CrossRef]
- Carillo, P.; Annunziata, M.G.; Pontecorvo, G.; Fuggi, A.; Woodrow, P. Salinity stress and salt tolerance. In Abiotic Stress in Plants–Mechanisms and Adaptations; Shanker, A., Venkateswarlu, B., Eds.; InTechOpen: Rijeka, Croatia, 2011; Volume 1, pp. 21–38. [Google Scholar] [CrossRef]
- Nchu, F.; Macuphe, N.; Rhoda, I.; Niekerk, L.-A.; Basson, G.; Keyster, M.; Etsassala, N.G.E.R. Endophytic Beauveria bassiana induces oxidative stress and enhances the growth of Fusarium oxysporum-infected tomato plants. Plants 2022, 11, 3182. [Google Scholar] [CrossRef]
- Rahnama, A.; Fakhri, S.; Meskarbashee, M. Root growth and architecture responses of bread wheat cultivars to salinity stress crop. Agron. J. 2019, 111, 2991–2998. [Google Scholar] [CrossRef]
- Shelden, M.C.; Dias, D.A.; Jayasinghe, N.S.; Bacic, A.; Roessner, U. Root spatial metabolite profiling of two genotypes of barley (Hordeum vulgare L.) reveals differences in response to short-term salt stress. J. Exp. Bot. 2016, 67, 3731–3745. [Google Scholar] [CrossRef]
- Robin, A.H.K.; Matthew, C.; Uddin, M.J.; Bayazid, K.N. Salinity-induced reduction in root surface area and changes in major root and shoot traits at the phytomer level in wheat. J. Exp. Bot. 2016, 67, 3719–3729. [Google Scholar] [CrossRef] [PubMed]
- Verslues, P.E.; Sharma, S. Proline metabolism and its implications for plant-environment interaction. Arab. Book 2010, 8, e0240. [Google Scholar] [CrossRef] [PubMed]
- Kaur, G.; Asthir, B. Proline: A key player in plant abiotic stress tolerance. Biol. Plant. 2015, 59, 609–619. [Google Scholar] [CrossRef]
- García, J.E.; Maroniche, G.; Creus, C.; Suárez-Rodríguez, R.; Ramirez-Trujillo, J.A.; Groppa, M.D. In vitro PGPR properties and osmotic tolerance of different Azospirillum native strains and their effects on growth of maize under drought stress. Microbiol. Res. 2017, 202, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Fan, D.; Subramanian, S.; Smith, D.L. Plant endophytes promote growth and alleviate salt stress in Arabidopsis thaliana. Sci. Rep. 2020, 10, 12740. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Number of Stolons, pcs. | Number of Leaves, pcs. | Leaf Surface Area, cm2 |
|---|---|---|---|
| Control | 2.82 ± 0.17 a | 6.96 ± 0.12 ab | 71.66 ± 2.49 a |
| B. bassiana | 3.37 ± 0.17 b | 7.02 ± 0.17 b | 59.62 ± 2.23 b |
| NaCl | 2.29 ± 0.16 c | 6.52 ± 0.13 c | 53.85 ± 2.47 b |
| B. bassiana + NaCl | 2.43 ± 0.17 ac | 6.61 ± 0.14 ac | 55.55 ± 2.08 b |
| Control | B. bassiana | NaCl | B. bassiana + NaCl | |
|---|---|---|---|---|
| Chlorophyll a (mg/g FW) | 2.11 ± 0.06 a | 2.04 ± 0.09 a | 2.01 ± 0.11 a | 2.04 ± 0.10 a |
| Chlorophyll b (mg/g FW) | 0.51 ± 0.03 ab | 0.52 ± 0.03 ab | 0.48 ± 0.02 a | 0.58 ± 0.04 b |
| Carotenoids (mg/g FW) | 0.53 ± 0.02 a | 0.51± 0.09 a | 0.50± 0.03 a | 0.51± 0.03 a |
| Total chlorophyll (mg/g FW) | 2.62 ± 0.08 a | 2.56 ± 0.11 a | 2.48 ± 0.13 a | 2.61 ± 0.13 a |
| Total pigments (mg/g FW) | 3.15 ± 0.10 a | 3.06 ± 0.14 a | 2.99 ± 0.16 a | 3.12 ± 0.15 a |
| Chlorophyll/carotenoids ratio | 4.96 ± 0.10 a | 5.09 ± 0.09 a | 4.96 ± 0.08 a | 5.17 ± 0.17 a |
| Chlorophyll a/b ratio | 4.27 ± 0.14 a | 4.07 ± 0.13 a | 4.19 ± 0.08 a | 3.67 ± 0.16 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tomilova, O.G.; Kryukova, N.A.; Efimova, M.V.; Kolomeichuk, L.V.; Kovtun, I.S.; Glupov, V.V. The Endophytic Entomopathogenic Fungus Beauveria bassiana Alleviates Adverse Effects of Salt Stress in Potato Plants. Horticulturae 2023, 9, 1140. https://doi.org/10.3390/horticulturae9101140
Tomilova OG, Kryukova NA, Efimova MV, Kolomeichuk LV, Kovtun IS, Glupov VV. The Endophytic Entomopathogenic Fungus Beauveria bassiana Alleviates Adverse Effects of Salt Stress in Potato Plants. Horticulturae. 2023; 9(10):1140. https://doi.org/10.3390/horticulturae9101140
Chicago/Turabian StyleTomilova, Oksana G., Natalia A. Kryukova, Marina V. Efimova, Liliya V. Kolomeichuk, Irina S. Kovtun, and Victor V. Glupov. 2023. "The Endophytic Entomopathogenic Fungus Beauveria bassiana Alleviates Adverse Effects of Salt Stress in Potato Plants" Horticulturae 9, no. 10: 1140. https://doi.org/10.3390/horticulturae9101140