
Barrelier’s Speedwell (Veronica barrelieri Schott ex Roem. et Schult., Plantaginaceae)—Potential of Free Volatile Compounds for Horticulture


 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
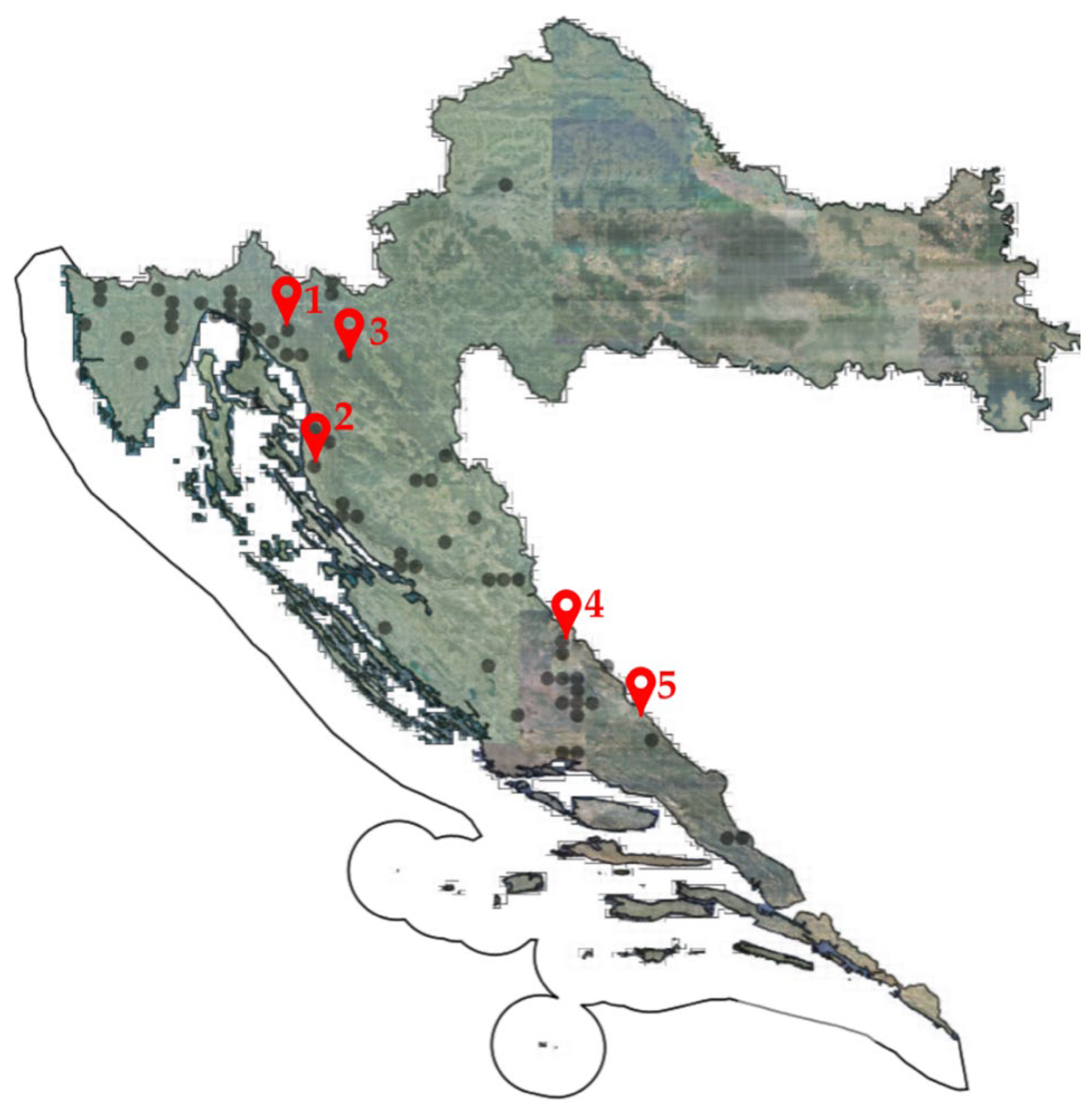
2.1. Plant Material
2.2. GC-MS Analyses
2.3. Micromorphological Analyses
2.4. Statistical Analyses
3. Results and Discussion
3.1. GC-MS Analyses
3.2. Principal Component Analyses
3.3. Micromorphological Studies
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Erhardt, W.; Götz, E.; Bödeker, N.; Seybold, S. Zander–Handwörterbuch der Pflanzennamen, 19th ed.; Eugen Ulmer GmbH und Co.: Stuttgart, Germany, 2014; p. 576. [Google Scholar]
- Albach, D.C.; Martínez-Ortega, M.M.; Fischer, M.A.; Chase, M.W. A new classification of the tribe Veroniceae–Problems and a possible solution. Taxon 2004, 53, 429–452. [Google Scholar] [CrossRef]
- Opiz, F.M. Seznam Rostlin Kveteny Ceske; Nákladem Českého Museum V kommissí u Fr. Řivnáče: Prague, Czech, 1852; p. 80. [Google Scholar]
- Lee, T.B.; Yamazaki, T. A revision of the Scrophulariaceae in Korea. Bull. Kwanak Arb. 1983, 4, 34–70. [Google Scholar]
- Xue, H.; Liu, J.; Ma, G.; Zhou, X.; Sun, L.; Shi, Y.; Zhu, W.; Chen, K.; Zhang, L.; Li, Y. Phenols and γ-Lactams from the aerial part of Pseudolysimachion linariifolium subsp. dilatatum. Chem. Biodivers. 2020, 17, e2000387. [Google Scholar] [CrossRef] [PubMed]
- Albach, D.C.; Fischer, M.A. AFLP- and genome size analyses: Contribution to the taxonomy of Veronica subg. Pseudolysimachium sect. Pseudolysimachion (Plantaginaceae), with a key to the European taxa. Phytol. Balcan. 2003, 9, 401–424. [Google Scholar]
- Nikolić, T. Flora Croatica; Alfa: Zagreb, Croatia, 2019; Volume 4, p. 612. [Google Scholar]
- Walters, S.M.; Webb, D.A. Veronica L. In Flora Europaea; Tutin, T.G., Heywood, V.H., Burges, N.A., Moore, D.M., Valentine, D.H., Walters, S.M., Webb, D.A., Eds.; Cambridge University Press: Cambridge, UK, 1972; Volume 3, pp. 242–251. [Google Scholar]
- Kremer, D.; Brkljačić, A.; Randić, M. The Flora of Premužić’s Trial on Velebit Mt; Public Institution Nature Park Velebit, Public Institution National Park North Velebit: Gospić, Croatia, 2019; p. 354. [Google Scholar]
- Popović, Z.; Smiljanić, M.; Kostić, M.; Nikić, P.; Janković, S. Wild flora and its usage in traditional phytotherpy (Deliblato Sands, Serbia, Southeast Europe). Ind. J. Trad. Knowl. 2014, 13, 9–35. [Google Scholar]
- Lee, S.U.; Sung, M.H.; Ryu, H.W.; Lee, J.; Kim, H.S.; In, H.J.; Ahn, K.S.; Lee, H.J.; Lee, H.K.; Shin, D.H.; et al. Verproside inhibits TNF-α-induced MUC5AC expression through suppression of the TNF-α/NF-κB pathway in human airway epithelial cells. Cytokine 2016, 77, 168–175. [Google Scholar] [CrossRef] [PubMed]
- Kwak, J.H.; Kim, H.J.; Lee, K.H.; Kang, S.C.; Zee, O.P. Antioxidative iridoid glycosides and phenolic compounds from Veronica peregrina. Arch. Pharm. Res. 2009, 32, 207–213. [Google Scholar] [CrossRef]
- Ertas, A.; Boga, M.; Kizil, M.; Ceken, B.; Goren, A.C.; Hasimi, N.; Demirci, S.; Topcu, G.; Kolak, U. Chemical profile and biological activities of Veronica thymoides subsp. pseudocinerea. Pharm. Biol. 2015, 53, 334–339. [Google Scholar] [CrossRef]
- Xue, H.; Chen, K.X.; Zhang, L.Q.; Li, Y.M. Review of the ethnopharmacology, phytochemistry, and pharmacology of the genus Veronica. Am. J. Chin. Med. 2019, 47, 1193–1221. [Google Scholar] [CrossRef]
- Salehi, B.; Shivaprasad Shetty, M.V.; Anil Kumar, N.; Živković, J.; Calina, D.; Oana Docea, A.; Emamzadeh-Yazdi, S.; Sibel Kılıç, C.; Goloshvili, T.; Nicola, S.; et al. Veronica plants-drifting from farm to traditional healing, food application, and phytopharmacology. Molecules 2019, 24, 2454. [Google Scholar] [CrossRef]
- Kosachev, P.A.; Mayland-Quellhorst, E.; Albach, D.C. Hybridization among species of Veronica subg. Pseudolysimachium in the Altai detected by SRAP markers. Nord. J. Bot. 2019, 37, e02209. [Google Scholar]
- Song, Y.W.; Cho, S.K. Phytol induces apoptosis and ROS-mediated protective autophagy in human gastric adenocarcinoma AGS cells. Biochem. Anal. Biochem. 2015, 4, 4. [Google Scholar] [CrossRef]
- Feng, L. Analysis of chemical constituents of essential oil in Veronica linariifolia by gas chromatography-mass spectrometry. Chin. J. Anal. Chem. 2002, 30, 822–825. [Google Scholar]
- Osmachko, A.; Kovaleva, A.; Goryacha, O.; Ili’ina, T. Components of essential oil of Veronica longifolia L. leaves and flovers. Pharm. Innov. 2014, 3, 1–6. [Google Scholar]
- Dunkić, V.; Kosalec, I.; Kosir, I.; Potocnik, T.; Cerenak, A.; Koncic, M.; Vitali, D.; Muller, I.; Kopricanec, M.; Bezic, N.; et al. Antioxidant and antimicrobial properties of Veronica spicata L. (Plantaginaceae). Curr. Drug Targets 2015, 16, 1660–1670. [Google Scholar] [CrossRef]
- Nazlić, M.; Kremer, D.; Jurišić-Grubešić, R.; Soldo, B.; Vuko, E.; Stabentheiner, E.; Ballian, D.; Bogunić, F.; Dunkić, V. Endemic Veronica saturejoides Vis. ssp. saturejoides–chemical composition and antioxidant activity of free volatile compounds. Plants 2020, 9, 1646. [Google Scholar] [CrossRef]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectrometry, 4th ed.; Allured Publishing: Carol Stream, IL, USA, 2017. [Google Scholar]
- Kremer, D.; Stabentheiner, E.; Bogunić, F.; Ballian, D.; Eleftheriadou, E.; Stešević, D.; Matevski, V.; Ranđelović, V.; Ivanova, D.; Ruščić, M.; et al. Micromorphological traits of Balcanic Micromeria and closely related Clinopodium species (Lamiaceae). Plants 2021, 10, 1666. [Google Scholar] [CrossRef]
- Payne, W.W. A glossary of plant hair terminology. Brittonia 1978, 30, 239–255. [Google Scholar] [CrossRef]
- Punt, W.; Hoen, P.P.; Blackmore, S.; Nilsson, S.; Le Thomas, A. Glossary of pollen and spore terminology. Rev. Palaeobot. Palynol. 2007, 143, 1–81. [Google Scholar] [CrossRef]
- Benelli, G.; Pavela, R.; Drenaggi, E.; Desneux, N.; Maggi, F. Phytol, (E)-nerolidol and spathulenol from Stevia rebaudiana leaf essential oil as effective and eco-friendly botanical insecticides against Metopolophium dirhodum. Ind. Crops Prod. 2020, 155, 112844. [Google Scholar] [CrossRef]
- Pavela, R.; Benelli, G. Essential oils as ecofriendly biopesticides? Challenges and constraints. Trends Plant Sci. 2016, 21, 1000–1007. [Google Scholar] [CrossRef] [PubMed]
- Fujimoto, T.; Abe, H.; Mizukubo, T.; Seo, S. Phytol, a constituent of chlorophyll, induces root-knot nematode resistance in Arabidopsis via the ethylene signaling pathway. Mol. Plant Microbe Interact. 2021, 34, 279–285. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.T.; Ali, E.S.; Uddin, S.J.; Shaw, S.; Islam, M.A.; Ahmed, M.I.; Chandra, S.M.; Karmakar, U.K.; Yarla, N.S.; Khan, I.N.; et al. Phytol: A review of biomedical activities. Food Chem. Toxicol. 2018, 121, 82–94. [Google Scholar] [CrossRef]
- Nazlić, M.; Fredotović, Ž.; Vuko, E.; Fabijanić, L.; Kremer, D.; Stabentheiner, E.; Ruščić, M.; Dunkić, V. Wild species Veronica officinalis L. and Veronica saturejoides Vis. ssp. saturejoides—Biological potential of free volatiles. Horticulturae 2021, 7, 295. [Google Scholar] [CrossRef]
- Nazlić, M.; Fredotović, Ž.; Vuko, E.; Vuletić, N.; Kremer, D.; Jurišić Grubešić, R.; Stabentheiner, E.; Randić, M.; Dunkić, V. Free volatile compounds of Veronica austriaca ssp. jacquinii (Baumg.) Eb. Fisch. and their biological activity. Plants 2021, 10, 2529. [Google Scholar] [CrossRef] [PubMed]
- Çelik, E.; Yuvali Çelik, G.; Meysun, A. Essential oil composition and antibacterial activity of some plant species. J. Appl. Biol. Sci. 2010, 1, 45–48. [Google Scholar] [CrossRef] [Green Version]
- Valyova, M.; Hadjimitova, V.; Stoyanov, S.; Ganeva, Y.; Trayko; Petkov, I. Free radical scavenging activity of extracts from Bulgarian Veronica officinalis L. and GC-MS analysis of ethanol extract. Internet J. Aesthetic Antiaging Med. 2008, 2, 1–5. [Google Scholar] [CrossRef]
- Kraehmer, H.; Baur, P. Weed Anatomy; Wiley-Blackwell: Hoboken, NJ, USA, 2013. [Google Scholar]
- Kurer, G.A. Kutikularfalten und Protuberanzen an Haaren und Epidermen; Druck Gebr. Leeman: Dresden, Germany, 1917. [Google Scholar]
- Kristen, U.; Lockhausen, J. The leaf glands of Veronica beccabunga L.: Ultrastructure and a possible pathway of secretion. Isr. J. Plant Sci. 2015, 34, 147–156. [Google Scholar] [CrossRef]
- Werker, E.; Ravid, U.; Putievsky, E. Structure of glandular hairs and identification of the main components of their secreted material in some species of the Labiatae. Isr. J. Bot. 1985, 34, 31–45. [Google Scholar] [CrossRef]
- Hanlidou, E.; Kokkini, S.; Bosabalidis, A.M.; Bessiere, J.-M. Glandular trichomes and essential oil constituents of Calamintha menthifolia (Lamiaceae). Plant Syst. Evol. 1991, 177, 17–26. [Google Scholar] [CrossRef]
- Ko, K.N.; Lee, K.W.; Lee, S.E.; Kim, E.S. Development and ultrastructure of glandular trichomes in Pelargonium × fragrans ‘Mabel Grey’ (Geraniaceae). J. Plant Biol. 2007, 50, 362–368. [Google Scholar] [CrossRef]
- Kremer, D.; Vitali Čepo, D.; Dunkić, D.; Dragojević Müller, I.; Kosalec, I.; Bezić, N.; Stabentheiner, E. Phytochemical and micromorphological traits of Geranium dalmaticum (Beck) Rech. f. and G. macrorrhizum L. (Geraniaceae). Nat. Prod. Commun. 2013, 8, 645–650. [Google Scholar] [CrossRef] [Green Version]
- Kremp, O.W. Morphologic Encyclopedia of Palynology; Univ. of Arizona Press: Tucson, AZ, USA, 1965. [Google Scholar]
- Tsymbalyuk, Z.M. Pollen morphology of species of Veronica L. (Scrophulariaceae) sect. Pseudolysimachium W. D. J. Koch in the flora of Ukraine. Ukr. Botan. J. 2008, 65, 823–835. (In Ukrainian) [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locality | Latitude | Longitude | Altitude a.s.l. (m) | Voucher No. |
|---|---|---|---|---|
| 1. Lič Polje | 45°16′27″ N | 14°45′00″ E | 728 | HFK-HR-2021-35 |
| 2. Velebit Mt | 44°43′56″ N | 14°56′57″ E | 1162 | HFK-HR-2021-49 |
| 3. Gornja Kamenica * | 45°02′43″ N | 15°07′41″ E | 645 | HFK-HR-2021-44 |
| 4. Dinara Mt | 44°01′54″ N | 16°24′36″ E | 1198 | HFK-HR-2021-72 |
| 5. Kamešnica Mt | 43°42′07″ N | 16°48′16″ E | 1250 | HFK-HR-2021-81 |
| Locality | Lič Polje | Gornja Kamenica | Velebit Mt | Dinara Mt | Kamešnica Mt | ||
|---|---|---|---|---|---|---|---|
| Component | RIa | RIb | VC ± SD | VC ± SD | VC ± SD | VC ± SD | VC ± SD |
| α-Thujene | 924 | 1012 | 0.82 ± 0.01 | 0.63 ± 0.01 | 1.15 ± 0.01 | 0.64 ± 0.01 | |
| α-Pinene * | 935 | 1017 | – | – | – | – | 0.70 ± 0.01 |
| Benzaldehyde | 952 | 1508 | 2.17 ± 0.01 | 0.533 ± 0.01 | 0.97 ± 0.01 | 1.25 ± 0.01 | 3.98 ± 0.01 |
| β -Phellandrene | 1002 | 1195 | – | – | – | 0.75 ± 0.1 | – |
| Benzene acetaldehyde | 1036 | 1633 | 3.06 ± 0.01 | 0.56 ± 0.01 | 1.33 ± 0.01 | – | 1.12 ± 0.01 |
| γ-Terpinene | 1057 | 1225 | – | – | – | 1.64 ± 0.01 | – |
| Linalool | 1095 | 1506 | 2.26 ± 0.01 | 0.70 ± 0.01 | 1.86 ± 0.01 | 2.32 ± 0.01 | 1.53 ± 0.15 |
| n-Nonanal | 1100 | 1389 | – | – | 1.69 ± 0.02 | – | 1.68 ± 0.01 |
| Terpinen-4-ol | 1174 | 1686 | 0.54 ± 0.01 | – | 1.12 ± 0.01 | – | – |
| Borneol | 1176 | 1719 | – | – | – | 1.59 ± 0.01 | – |
| α-Terpineol | 1184 | 1660 | 2.95 ± 0.01 | 3.31 ± 0.01 | 0.66 ± 0.01 | 3.08 ± 0.03 | 1.31 ± 0.01 |
| trans-p-Mentha-1(7),8-dien-2-ol | 1187 | 1803 | – | – | 0.72 ± 0.01 | – | 0.79 ± 0.01 |
| α-Copaene | 1377 | 1484 | – | – | – | – | 0.55 ± 0.01 |
| (E)-β-Damascenone | 1384 | 1819 | 3.86 ± 0.01 | 1.42 ± 0.01 | 0.42 ± 0.01 | 5.69 ± 0.01 | – |
| Methyl eugenol | 1403 | 2005 | 1.01 ± 0.01 | 0.33 ± 0.01 | 1.85 ± 0.01 | 2.34 ± 0.01 | 2.21 ± 0.01 |
| E-Caryophyllene * | 1424 | 1585 | 1.23 ± 0.01 | 2.03 ± 0.01 | 1.43 ± 0.1 | 3.48 ± 0.01 | 1.48 ± 0.01 |
| (Z)-Methyl isoeugenol | 1451 | 2070 | – | 1.35 ± 0.01 | – | – | – |
| allo-Aromadendrene | 1465 | 1662 | – | – | 0.97 ± 0.01 | 2.59 ± 0.01 | 1.21 ± 0.01 |
| β-Chamigrene | 1478 | 1724 | – | – | – | 0.67 ± 0.02 | – |
| Germacrene D | 1481 | 1692 | 0.48 ± 0.01 | 0.71 ± 0.01 | 0.56 ± 0.01 | 2.43 ± 0.01 | 2.61 ± 0.01 |
| β-Ionone | 1487 | 1935 | 4.34 ± 0.01 | 4.11 ± 0.01 | 3.56 ± 0.01 | 3.42 ± 0.01 | 3.15 ± 0.01 |
| δ-Selinene | 1492 | 1756 | 0.66 ± 0.01 | – | – | 0.54 ± 0.01 | – |
| δ-Cadinene | 1517 | 1745 | – | – | – | – | 0.81 ± 0.01 |
| Spathulenol | 1577 | 2101 | – | – | 0.45 ± 0.01 | 0.25 ± 0.01 | 0.83 ± 0.01 |
| Caryophyllene oxide * | 1581 | 1955 | 4.43 ± 0.01 | 4.18 ± 0.01 | 3.43 ± 0.01 | 3.76 ± 0.01 | 4.51 ± 0.01 |
| Viridiflorol | 1592 | 2099 | 0.65 ± 0.01 | – | – | – | 0.54 ± 0.01 |
| γ-Eudesmol | 1632 | 2175 | – | – | – | – | 0.22 ± 0.01 |
| α-Muurolol | 1645 | 2181 | 2.38 ± 0.01 | 2.06 ± 0.01 | 1.33 ± 0.01 | 2.31 ± 0.01 | 2.52 ± 0.01 |
| Hexahydrofarnesyl acetone * | 1839 | 2113 | 1.56 ± 0.01 | 1.14 ± 0.01 | 0.56 ± 0.01 | 1.31 ± 0.1 | 0.84 ± 0.01 |
| Phytol * | 1942 | 2610 | 37.78 ± 0.01 | 40.32 ± 0.1 | 30.08 ± 0.1 | 32.88 ± 0.02 | 36.43 ± 0.01 |
| Hexadecanoic acid * | 1959 | 2912 | 12.45 ± 0.01 | 12.14 ± 0.1 | 8.75 ± 0.01 | 9.04 ± 0.01 | 12.44 ± 0.01 |
| Eicosane * | 2000 | 2000 | 0.38 ± 0.0 | 3.36 ± 0.01 | 1.22 ± 0.01 | - | 1.12 ± 0.01 |
| Heneicosane * | 2100 | 2100 | – | 0.65 ± 0.01 | 1.98 ± 0.1 | 1.32 ± 0.01 | – |
| Docosane * | 2200 | 2200 | 2.01 ± 0.01 | 7.69 ± 0.01 | 2.13 ± 0.01 | 2.15 ± 0.01 | 0.43 ± 0.01 |
| Tricosane * | 2300 | 2300 | – | – | – | 1.66 ± 0.01 | – |
| Tetracosane * | 2400 | 2400 | 4.61 ± 0.01 | 3.38 ± 0.01 | 9.32 ± 0.01 | 1.72 ± 0.01 | – |
| Pentacosane * | 2500 | 2500 | 3.54 ± 0.01 | 4.03 ± 0.01 | 10.71 ± 0.1 | 5.25 ± 0.1 | 10.32 ± 0.01 |
| Octacosane * | 2800 | 2800 | 0.75 ± 0.01 | – | 6.10 ± 0.01 | – | – |
| Total identification (%) | 93.92 | 94.63 | 94.35 | 93.41 | 93.97 | ||
| Grouped compounds (%) | |||||||
| Monoterpene hydrocarbons | 0.82 | 0.63 | 1.15 | 0.75 | 1.34 | ||
| Oxygenated monoterpenes | 5.75 | 4.01 | 4.36 | 8.63 | 3.63 | ||
| Sesquiterpene hydrocarbons | 2.37 | 2.74 | 2.96 | 9.68 | 6.66 | ||
| Oxygenated sesquiterpenes | 9.02 | 7.38 | 5.77 | 7.63 | 9.46 | ||
| Oxygenated diterpenes | 37.78 | 40.32 | 30.08 | 32.88 | 36.43 | ||
| Hydrocarbons | 11.29 | 19.11 | 31.46 | 12.1 | 11.87 | ||
| Others | 26.89 | 20.44 | 18.57 | 21.74 | 24.58 |
| Locality | Trichomes | Leaf | Calyx | Stem | |
|---|---|---|---|---|---|
| Adaxial | Abaxial | ||||
| Lič Polje | NG | + | + | + | +/++ |
| capitate C1 | + | + | ± | + | |
| capitate C2 | ± | ± | ± | ± | |
| NG | + | + | + | + | |
| Gornja Kemenica | capitate C1 | + | + | + | + |
| capitate C2 | ± | +/± | ± | +/± | |
| Velebit Mt | NG | + | + | + | + |
| capitate C1 | + | + | ±/+ | + | |
| capitate C2 | ± | +/± | –/± | +/± | |
| Dinara Mt | NG | + | + | + | + |
| capitate C1 | + | + | ± | + | |
| capitate C2 | ± | ± | ± | ± | |
| Kamešnica Mt | NG | + | + | + | + |
| capitate C1 | + | + | + | + | |
| capitate C2 | –/± | ± | ± | ± | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kremer, D.; Stabentheiner, E.; Nazlić, M.; Randić, M.; Srečec, S.; Dunkić, V. Barrelier’s Speedwell (Veronica barrelieri Schott ex Roem. et Schult., Plantaginaceae)—Potential of Free Volatile Compounds for Horticulture. Horticulturae 2022, 8, 777. https://doi.org/10.3390/horticulturae8090777
Kremer D, Stabentheiner E, Nazlić M, Randić M, Srečec S, Dunkić V. Barrelier’s Speedwell (Veronica barrelieri Schott ex Roem. et Schult., Plantaginaceae)—Potential of Free Volatile Compounds for Horticulture. Horticulturae. 2022; 8(9):777. https://doi.org/10.3390/horticulturae8090777
Chicago/Turabian StyleKremer, Dario, Edith Stabentheiner, Marija Nazlić, Marko Randić, Siniša Srečec, and Valerija Dunkić. 2022. "Barrelier’s Speedwell (Veronica barrelieri Schott ex Roem. et Schult., Plantaginaceae)—Potential of Free Volatile Compounds for Horticulture" Horticulturae 8, no. 9: 777. https://doi.org/10.3390/horticulturae8090777