
Bio-Circular Perspective of Citrus Fruit Loss Caused by Pathogens: Occurrences, Active Ingredient Recovery and Applications

 , , ,
, , ,  ,
,
Abstract
:1. Introduction
2. Citrus Pre-Harvest Losses
2.1. Gum Diseases of Citrus Trees Caused by Phytophthora spp. Infection
2.2. Citrus Greening Disease
2.3. Citrus Bacterial Disease
2.3.1. Citrus Canker Disease
2.3.2. Bacterial Blast Disease

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Citrus Fruit Diseases | Disease Characteristics | Types of by-Products | Citations | |
|---|---|---|---|---|
| Citrus gumnosis |  | Pathogen: Phytophthora spp. Symptoms: Cankers and gum on citrus trunks and branches. The infection causes damage to fibrous roots in vulnerable rootstocks and crown rot, thereby causing fruit fall. | Roots, trunk, branches, shoots die off and fruit drop | [40,41,42] |
| Citrus greening |  | Pathogen: Candidatus Liberibacter Symptoms: Pathogens infiltrate the phloem and assault the vascular system, blocking water and nutrient transport. Yellow shoots and mottled leaves with yellow midribs may occur causing die-back. Infected fruits are lobbed, have aborted seeds, and prematurely drop off. | Fruit drop | [62,63] |
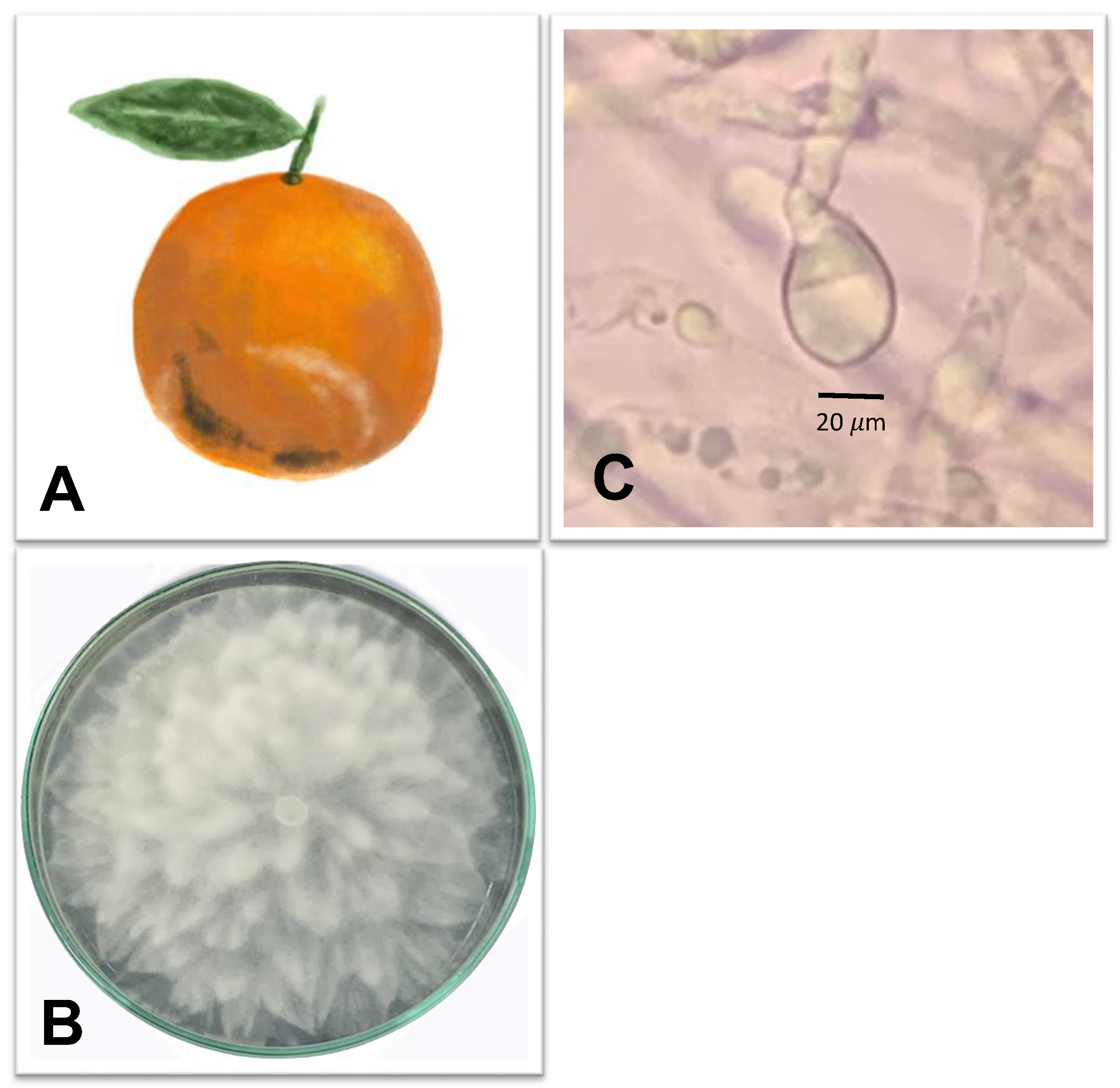
| Black rot |  | Pathogen: Alternaria spp. Symptoms: On fruits, symptoms range from light brown, depressed patches to dark brown circles. Young shoot apexes are defoliated in severely affected trees. Young infected fruits and leaves fall, and mature lesions-covered fruits are unmarketable. Alternaria spp. can cause black rot by causing latent infections on the calyx and disc, then entering the columella as the fruit grows. | Die shoots and fruit drops | [64,65,66] |
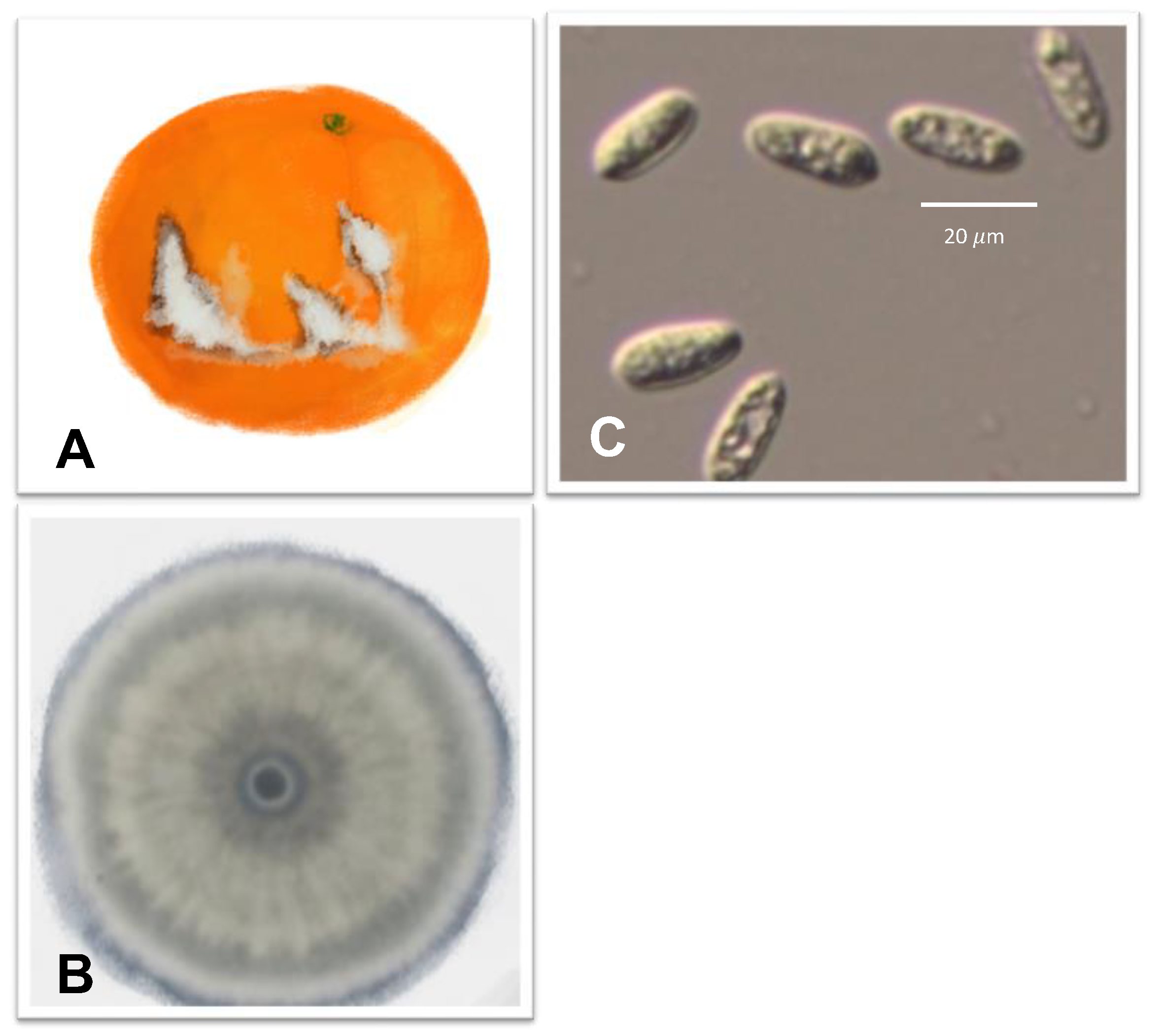
| Brown rot |  | Pathogen: Phytophthora citrophthora and Phytophthora nicotianae Symptom: light brown, leathery decay. This disease is associated with citrus gummosis or citrus foot rot. | Fruit drops | [67,68,69,70] |
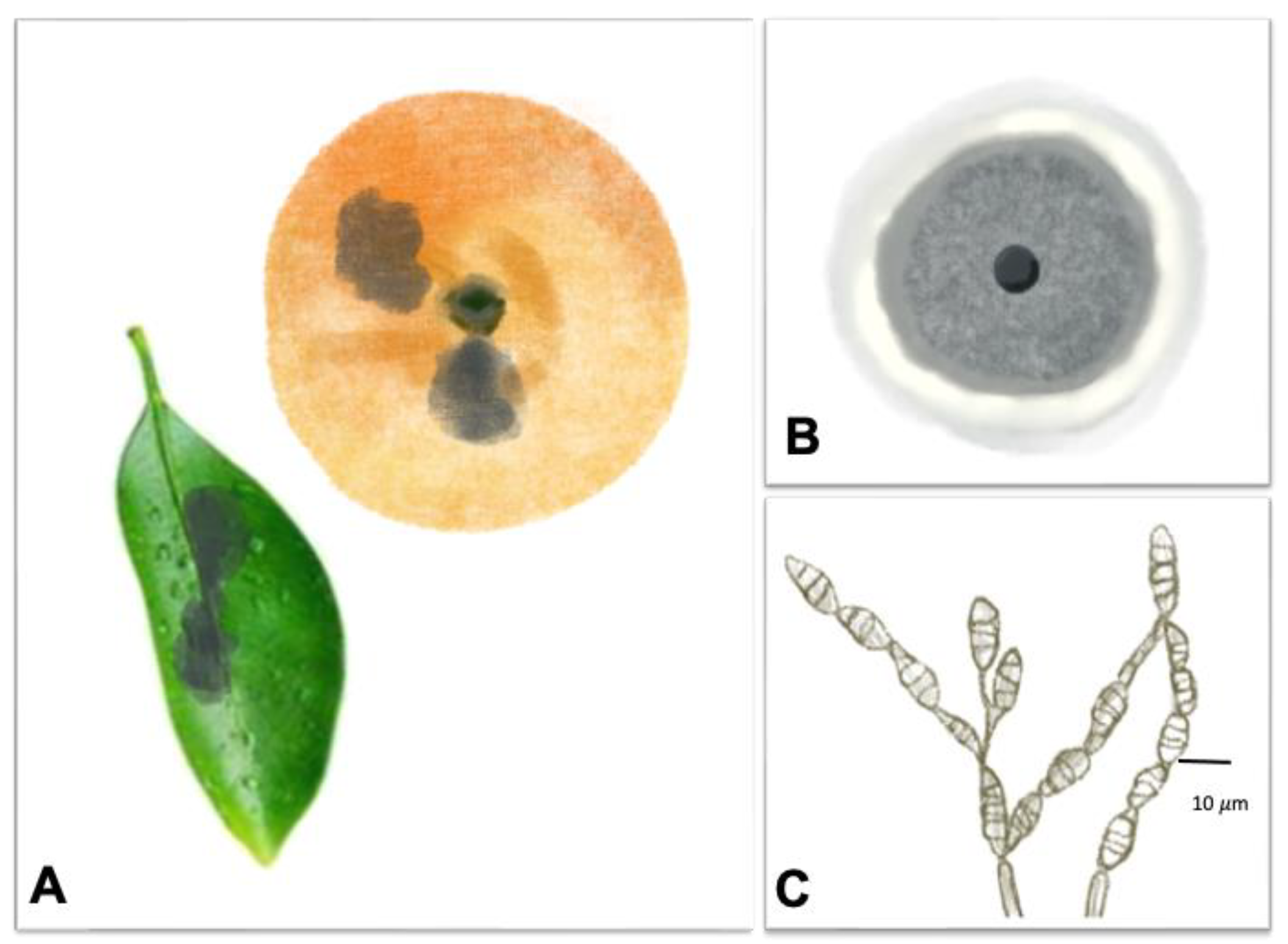
| Anthracnose |  | Pathogen: Colletotrichum spp. Symptoms: The disease caused necrotic petals and fruit lesions during pre-harvesting. Infected twigs and lesions generally had black fructifications. During postharvest, the infected fruits had brown-to-black tear stains that turned silver-gray, 1.5 mm or bigger lesions. | Branch die-off, fruit drops during pre-harvesting stage and fruit loss during postharvesting | [71,72] |
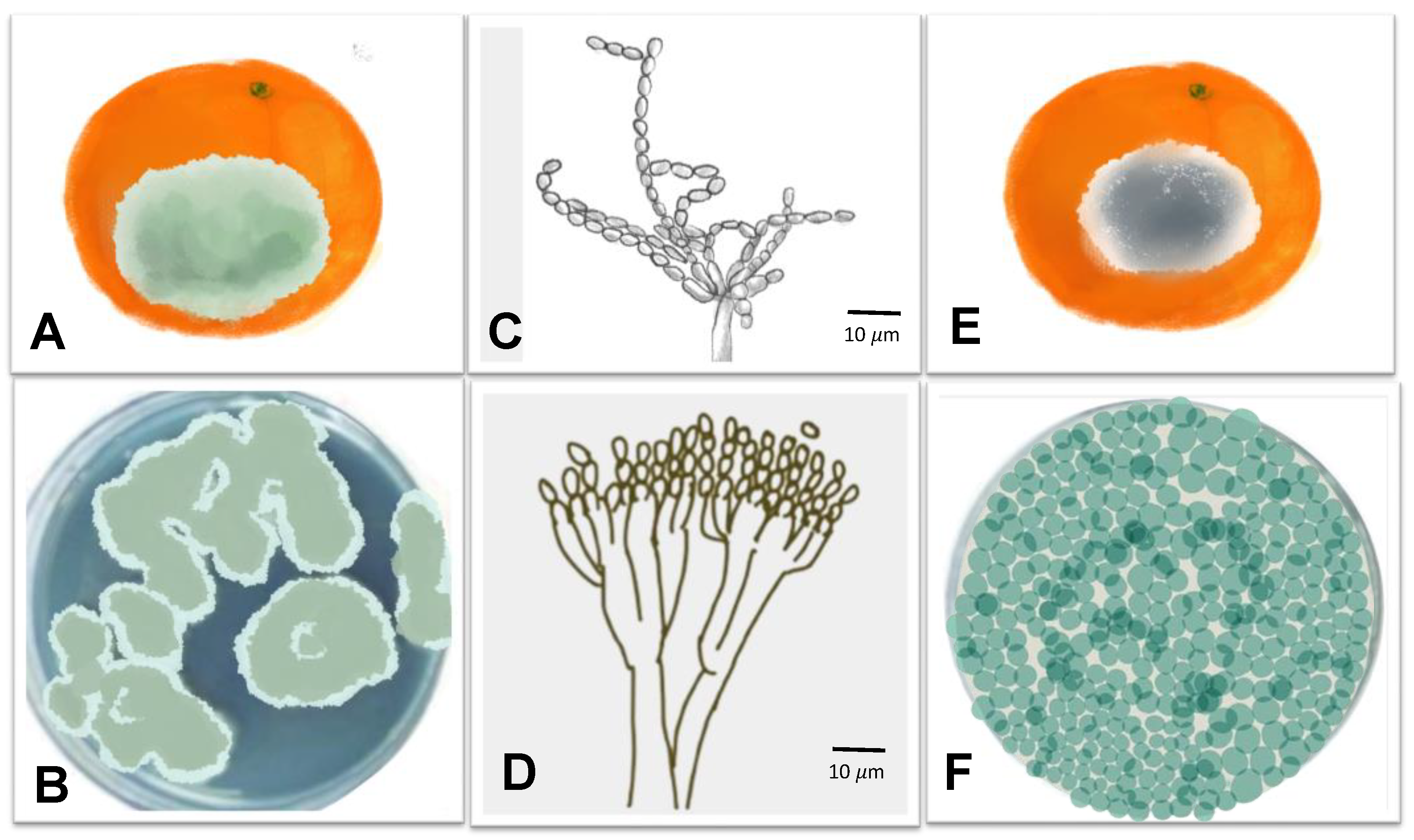
| Green and blue mold |  | Pathogen: Penicillium digitatum and P. italicum Symptoms: during postharvest, the fruit peel is soft and decolored and soaky. During disease development, the fruit surface is covered with aerial white mycelium turns to olive with spore development. | Fruit loss | [73,74] |
| Sour rot |  | Pathogen: Geotrichum citri-aurantii Symptoms: Storage fruits in the humid condition cause the fungus growth with a light brown to yellow tint. The symptom appears extensive water-soaked lesions, and artroconidia and mycelia on the fruit surface. | Fruit loss | [75,76,77] |
3. Citrus Postharvest Disease
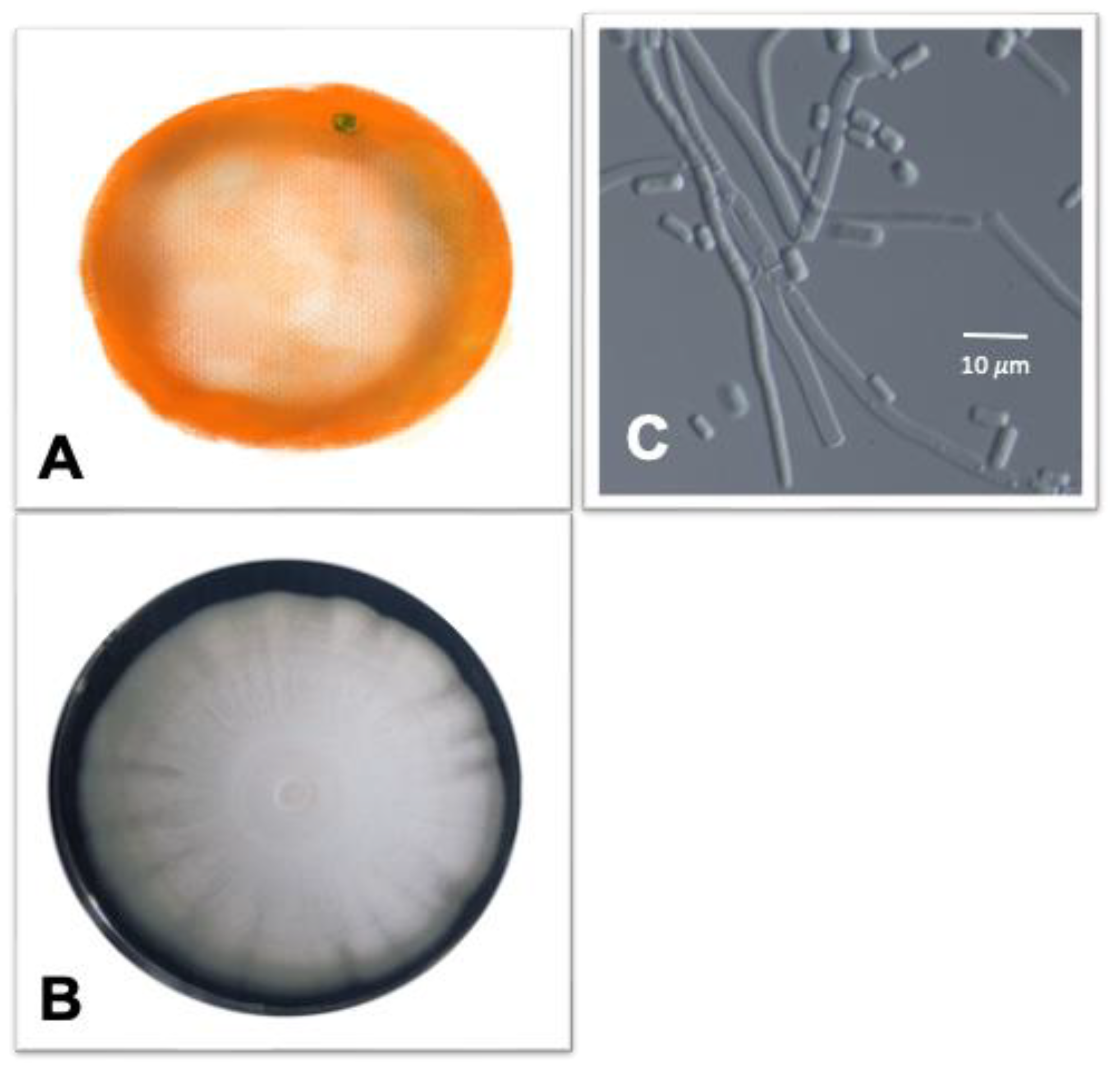
3.1. Black Rot Caused by Alternaria spp.
3.2. Brown Rot Caused by Phytophthora citrophthora
3.3. Anthracnose Caused by Colletotrichum spp.
3.4. Green and Blue Molds Caused, Respectively, by Penicillium digitatum Sacc. and P. italicum
3.5. Sour Rot Caused by Geotrichum citri-aurantii
4. High-Value Component Recovery
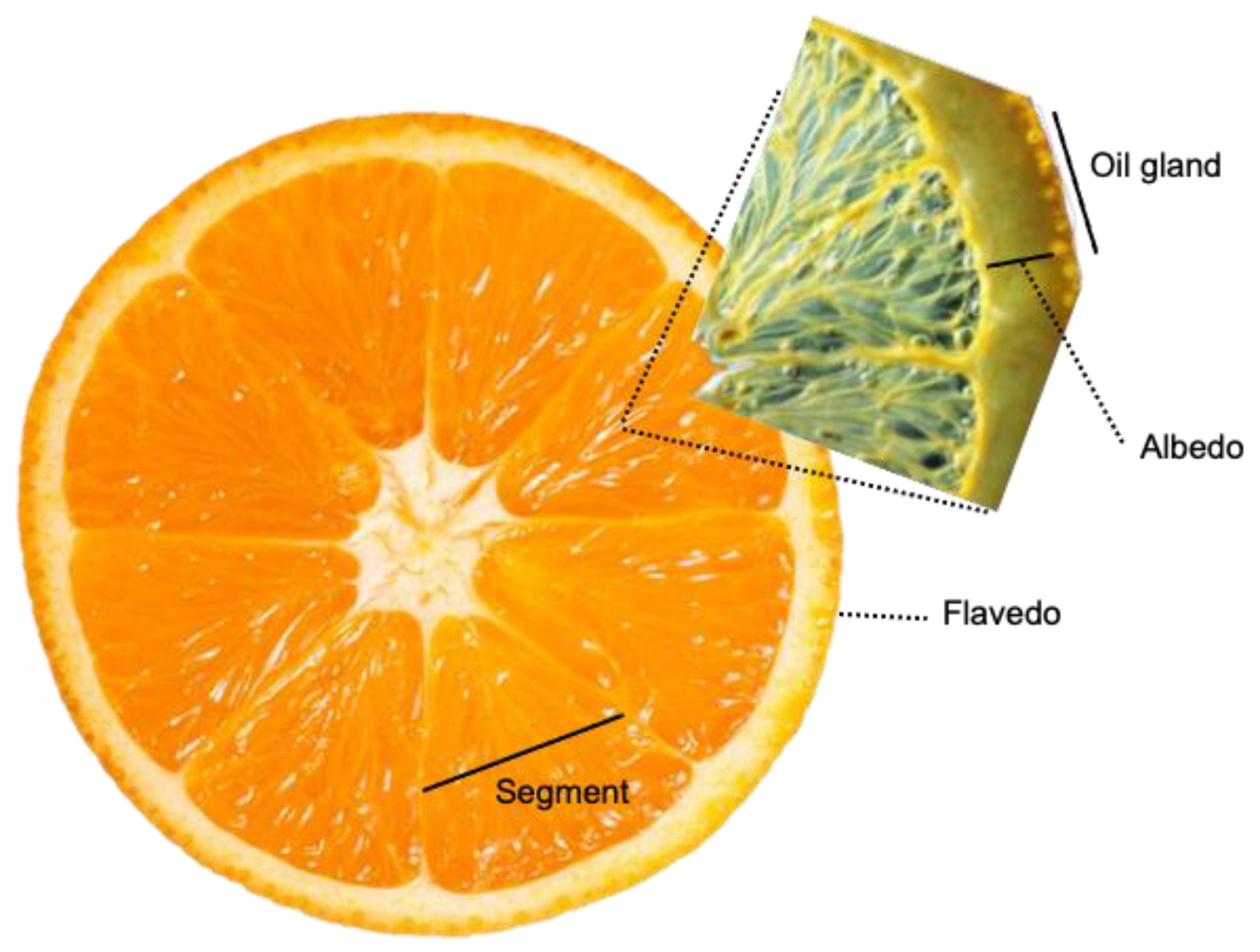
4.1. Citrus Essential Oils
4.2. Citrus Polysaccharides
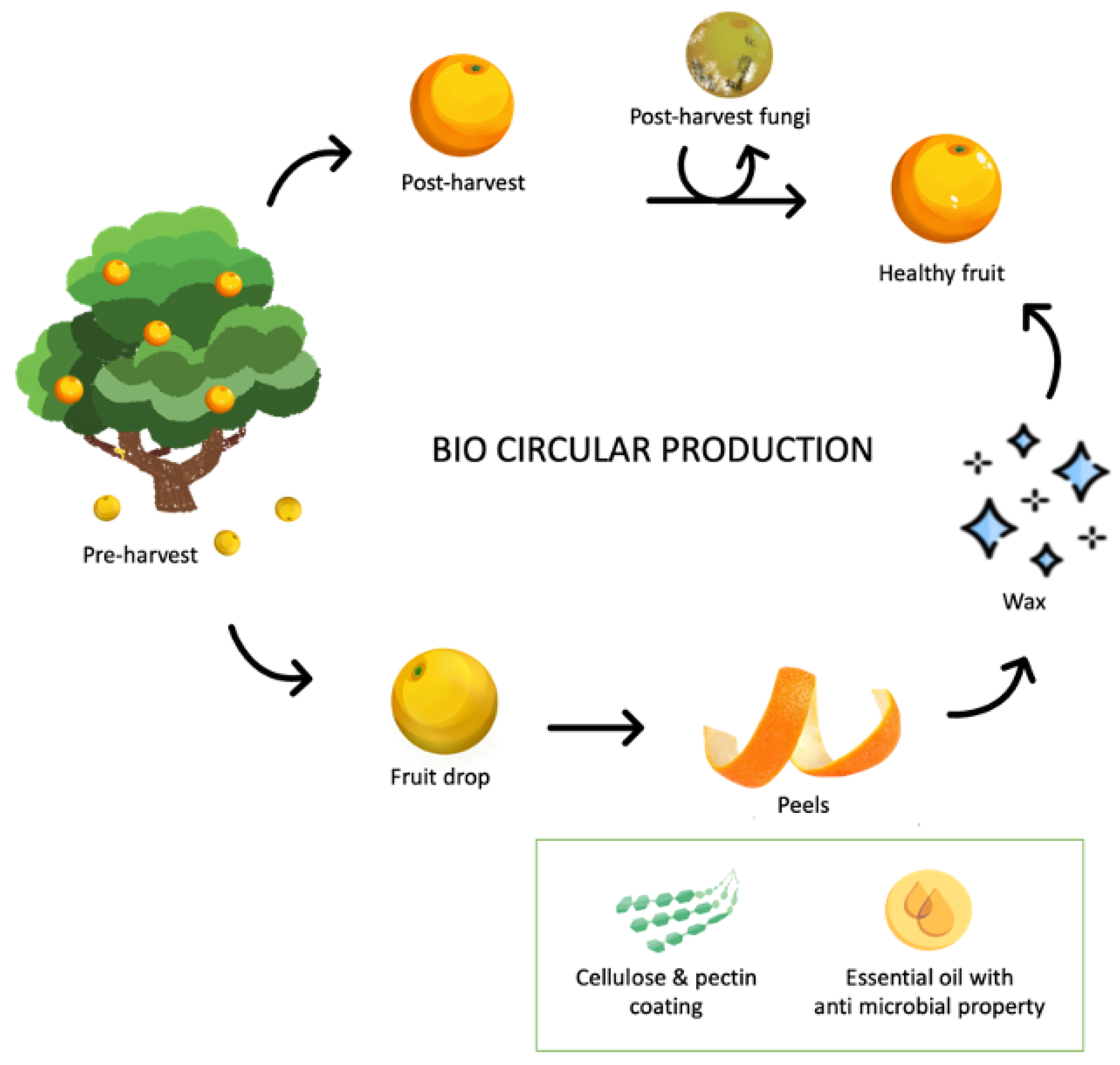
5. Bio-Circular Approaches
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- UN. Transforming Our World: The 2030 Agenda for Sustainable Development. Available online: https://sdgs.un.org/2030agenda (accessed on 17 May 2022).
- Barros, M.V.; Salvador, R.; de Francisco, A.C.; Piekarski, C.M. Mapping of research lines on circular economy practices in agriculture: From waste to energy. Renew. Sustain. Energy Rev. 2020, 131, 109958. [Google Scholar] [CrossRef]
- Jacob-John, J.; D’Souza, C.; Marjoribanks, T.; Singaraju, S. Synergistic Interactions of SDGs in Food Supply Chains: A Review of Responsible Consumption and Production. Sustainability 2021, 13, 8809. [Google Scholar] [CrossRef]
- Madau, F.A.; Arru, B.; Furesi, R.; Pulina, P. Insect Farming for Feed and Food Production from a Circular Business Model Perspective. Sustainability 2020, 12, 5418. [Google Scholar] [CrossRef]
- Lim, M.K.; Lai, M.; Wang, C.; Lee, Y. Circular economy to ensure production operational sustainability: A green-lean approach. Sustain. Prod. Consum. 2022, 30, 130–144. [Google Scholar] [CrossRef]
- Esposito, B.; Sessa, M.R.; Sica, D.; Malandrino, O. Towards circular economy in the agri-food sector. A systematic literature review. Sustainability 2020, 12, 7401. [Google Scholar] [CrossRef]
- Zhang, Q.; Dhir, A.; Kaur, P. Circular economy and the food sector: A systematic literature review. Sustain. Prod. Consum. 2022, 32, 655–668. [Google Scholar] [CrossRef]
- Van Fan, Y.; Lee, C.T.; Lim, J.S.; Klemeš, J.J.; Le, P.T.K. Cross-disciplinary approaches towards smart, resilient and sustainable circular economy. J. Clean. Prod. 2019, 232, 1482–1491. [Google Scholar] [CrossRef]
- Sherwood, J. The significance of biomass in a circular economy. Bioresour. Technol. 2020, 300, 122755. [Google Scholar] [CrossRef]
- FAO. Citrus Fruit Fresh and Process; Food and Agriculture Organization of the United Nations: Rome, Italy, 2021. [Google Scholar]
- El-Otmani, M.; Ait-Oubahou, A.; Zacarías, L. Citrus spp.: Orange, mandarin, tangerine, clementine, grapefruit, pomelo, lemon and lime. In Postharvest Biology and Technology of Tropical and Subtropical Fruits; Elsevier: Amsterdam, The Netherlands, 2011; pp. 437–516e. [Google Scholar]
- Goldenberg, L.; Yaniv, Y.; Porat, R.; Carmi, N. Mandarin fruit quality: A review. J. Sci. Food Agric. 2018, 98, 18–26. [Google Scholar] [CrossRef]
- Forsyth, J.; Damiani, J. CITRUS FRUITS | Types on the Market. In Encyclopedia of Food Sciences and Nutrition, 2nd ed.; Caballero, B., Ed.; Academic Press: Oxford, UK, 2003; pp. 1329–1335. [Google Scholar] [CrossRef]
- Dutta, S.K.; Gurung, G.; Yadav, A.; Laha, R.; Mishra, V.K. Factors associated with citrus fruit abscission and management strategies developed so far: A review. N. Z. J. Crop Hortic. Sci. 2022, 1–22. [Google Scholar] [CrossRef]
- Gulfishan, M.; Jahan, A.; Bhat, T.A.; Sahab, D. Chapter 16-Plant Senescence and Organ Abscission. In Senescence Signalling and Control in Plants; Sarwat, M., Tuteja, N., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 255–272. [Google Scholar] [CrossRef]
- Sawicki, M.; Aït Barka, E.; Clément, C.; Vaillant-Gaveau, N.; Jacquard, C. Cross-talk between environmental stresses and plant metabolism during reproductive organ abscission. J. Exp. Bot. 2015, 66, 1707–1719. [Google Scholar] [CrossRef] [PubMed]
- Iglesias, D.J.; Cercós, M.; Colmenero-Flores, J.M.; Naranjo, M.A.; Ríos, G.; Carrera, E.; Ruiz-Rivero, O.; Lliso, I.; Morillon, R.; Tadeo, F.R. Physiology of citrus fruiting. Braz. J. Plant Physiol. 2007, 19, 333–362. [Google Scholar] [CrossRef]
- Gustafsson, J.; Cederberg, C.; Sonesson, U.; Emanuelsson, A. The Methodology of the FAO Study: Global Food Losses and Food Waste-Extent, Causes and Prevention-FAO, 2011; SIK Institutet för Livsmedel och Bioteknik: Borås, Sweden, 2013. [Google Scholar]
- Perondi, D.; Fraisse, C.W.; Dewdney, M.M.; Cerbaro, V.A.; Andreis, J.H.D.; Gama, A.B.; Junior, G.J.S.; Amorim, L.; Pavan, W.; Peres, N.A. Citrus advisory system: A web-based postbloom fruit drop disease alert system. Comput. Electron. Agric. 2020, 178, 105781. [Google Scholar] [CrossRef]
- Lima, W.G.; Spósito, M.B.; Amorim, L.; Gonçalves, F.P.; de Filho, P.A.M. Colletotrichum gloeosporioides, a new causal agent of citrus post-bloom fruit drop. Eur. J. Plant Pathol. 2011, 131, 157. [Google Scholar] [CrossRef]
- Huchche, A.; Ladaniya, M. Citrus Flowering and Fruiting—Recent Research Advances. In Proceedings of the Souvenir, National Seminar-Cum-Workshop on Physiology of Flowering in Perennial Fruit Crops; Central Institute for Subtropical Horticulture (ICAR): Lucknow, India, 2014; pp. 74–88. [Google Scholar]
- Syvertsen, J.; Garcia-Sanchez, F. Multiple abiotic stresses occurring with salinity stress in citrus. Environ. Exp. Bot. 2014, 103, 128–137. [Google Scholar] [CrossRef]
- Sato, K. Influence of drought and high temperature on citrus. In Abiotic Stress Biology in Horticultural Plants; Springer: Berlin/Heidelberg, Germany, 2015; pp. 77–86. [Google Scholar]
- Urbaneja, A.; Grout, T.G.; Gravena, S.; Wu, F.; Cen, Y.; Stansly, P.A. Chapter 16-Citrus pests in a global world. In The Genus Citrus; Talon, M., Caruso, M., Gmitter, F.G., Eds.; Woodhead Publishing: Cambridge, UK, 2020; pp. 333–348. [Google Scholar] [CrossRef]
- Khamsaw, P.; Lumsangkul, C.; Karunarathna, A.; Onsa, N.E.; Kawichai, S.; Chuttong, B.; Sommano, S.R. Recovery of Orange Peel Essential Oil from ‘Sai-Namphaung’ Tangerine Fruit Drop Biomass and Its Potential Use as Citrus Fruit Postharvest Diseases Control. Agriculture 2022, 12, 701. [Google Scholar] [CrossRef]
- Moraes Bazioli, J.; Belinato, J.R.; Costa, J.H.; Akiyama, D.Y.; Pontes, J.G.d.M.; Kupper, K.C.; Augusto, F.; de Carvalho, J.E.; Fill, T.P. Biological control of citrus postharvest phytopathogens. Toxins 2019, 11, 460. [Google Scholar] [CrossRef]
- Talibi, I.; Boubaker, H.; Boudyach, E.; Ait Ben Aoumar, A. Alternative methods for the control of postharvest citrus diseases. J. Appl. Microbiol. 2014, 117, 1–17. [Google Scholar] [CrossRef]
- Boluda-Aguilar, M.; García-Vidal, L.; del Pilar González-Castañeda, F.; López-Gómez, A. Mandarin peel wastes pretreatment with steam explosion for bioethanol production. Bioresour. Technol. 2010, 101, 3506–3513. [Google Scholar] [CrossRef]
- de Barros, C.H.N.; Cruz, G.C.F.; Mayrink, W.; Tasic, L. Bio-based synthesis of silver nanoparticles from orange waste: Effects of distinct biomolecule coatings on size, morphology, and antimicrobial activity. Nanotechnol. Sci. Appl. 2018, 11, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Yi, F.; Jin, R.; Sun, J.; Ma, B.; Bao, X. Evaluation of mechanical-pressed essential oil from Nanfeng mandarin (Citrus reticulata Blanco cv. Kinokuni) as a food preservative based on antimicrobial and antioxidant activities. LWT 2018, 95, 346–353. [Google Scholar] [CrossRef]
- Joglekar, S.N.; Pathak, P.D.; Mandavgane, S.A.; Kulkarni, B.D. Process of fruit peel waste biorefinery: A case study of citrus waste biorefinery, its environmental impacts and recommendations. Environ. Sci. Pollut. Res. 2019, 26, 34713–34722. [Google Scholar] [CrossRef] [PubMed]
- Tao, N.; Jia, L.; Zhou, H. Anti-fungal activity of Citrus reticulata Blanco essential oil against Penicillium italicum and Penicillium digitatum. Food Chem. 2014, 153, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Hamad, Y.K.; Fahmi, M.M.; Zaitoun, F.M.; Ziyada, S.M. Role of essential oils in controlling fungi that cause decline disease of guava. Int. J. Pure Appl. Biosci. 2015, 3, 143–151. [Google Scholar] [CrossRef]
- Hosseini, S.S.; Khodaiyan, F.; Yarmand, M.S. Optimization of microwave assisted extraction of pectin from sour orange peel and its physicochemical properties. Carbohydr. Polym. 2016, 140, 59–65. [Google Scholar] [CrossRef]
- Meneguzzo, F.; Ciriminna, R.; Zabini, F.; Pagliaro, M. Review of evidence available on hesperidin-rich products as potential tools against COVID-19 and hydrodynamic cavitation-based extraction as a method of increasing their production. Processes 2020, 8, 549. [Google Scholar] [CrossRef]
- Chen, J.; Cheng, H.; Zhi, Z.; Zhang, H.; Linhardt, R.J.; Zhang, F.; Chen, S.; Ye, X. Extraction temperature is a decisive factor for the properties of pectin. Food Hydrocoll. 2021, 112, 106160. [Google Scholar] [CrossRef]
- Graham, J.; Feichtenberger, E. Citrus Phytophthora diseases: Management challenges and successes. J. Citrus Pathol. 2015, 2. [Google Scholar] [CrossRef]
- Singh, K.; Sharma, R.; Dubey, A.; Kamil, D.; Lekshmy, S.; Awasthi, O.; Jha, G. Phytophthora Nicotianae Breda de Haan Induced Stress Changes in Citrus Rootstock Genotypes; NISCAIR-CSIR: New Delhi, India, 2019. [Google Scholar]
- Rajput, N.A.; Atiq, M.; Tariq, H.; Saddique, W.M.; Hameed, A. Citrus Gummosis: A Formidable challenge to citrus industry: A Review. Int. J. Biosci. 2020, 16, 131–144. [Google Scholar]
- Gottwald, J.R.; Krysan, P.J.; Young, J.C.; Evert, R.F.; Sussman, M.R. Genetic evidence for the in planta role of phloem-specific plasma membrane sucrose transporters. Proc. Natl. Acad. Sci. USA 2000, 97, 13979–13984. [Google Scholar] [CrossRef]
- Erwin, D.C.; Ribeiro, O.K. Phytophthora: Diseases Worldwide; APS Press: St. Paul, MN, USA, 1996. [Google Scholar]
- Timmer, L.W.; Duncan, L.W. Citrus Health Management; APS Press: St. Paul, MN, USA, 1999. [Google Scholar]
- Savita, G.S.V.; Nagpal, A. Citrus diseases caused by Phytophthora species. GERF Bull. Biosci. 2012, 3, 18–27. [Google Scholar]
- Agrios, G.N. Plant Pathology; Elsevier: Amsterdam, The Netherlands, 2005. [Google Scholar]
- Lee, R.F. Chapter Five-Control of Virus Diseases of Citrus. In Advances in Virus Research; Loebenstein, G., Katis, N.I., Eds.; Academic Press: Cambridge, MA, USA, 2015; Volume 91, pp. 143–173. [Google Scholar]
- Berk, Z. Chapter 5-Diseases and pests. In Citrus Fruit Processing; Berk, Z., Ed.; Academic Press: San Diego, CA, USA, 2016; pp. 83–93. [Google Scholar] [CrossRef]
- Methacanon, P.; Krongsin, J.; Gamonpilas, C. Pomelo (Citrus maxima) pectin: Effects of extraction parameters and its properties. Food Hydrocoll. 2014, 35, 383–391. [Google Scholar] [CrossRef]
- Plaza, P.; Torres, R.; Usall, J.; Lamarca, N.; Vinas, I. Evaluation of the potential of commercial post-harvest application of essential oils to control citrus decay. J. Hortic. Sci. Biotechnol. 2004, 79, 935–940. [Google Scholar] [CrossRef]
- Gasparoto, M.; Coletta-Filho, H.; Bassanezi, R.; Lopes, S.; Lourenço, S.; Amorim, L. Influence of temperature on infection and establishment of ‘Candidatus Liberibacter americanus’ and ‘Candidatus Liberibacter asiaticus’ in citrus plants. Plant Pathol. 2012, 61, 658–664. [Google Scholar] [CrossRef]
- Dala-Paula, B.M.; Plotto, A.; Bai, J.; Manthey, J.A.; Baldwin, E.A.; Ferrarezi, R.S.; Gloria, M.B.A. Effect of Huanglongbing or Greening Disease on Orange Juice Quality, a Review. Front. Plant Sci. 2019, 9, 1976. [Google Scholar] [CrossRef]
- Gottwald, T.R. Current Epidemiological Understanding of Citrus Huanglongbing. Annu. Rev. Phytopathol. 2010, 48, 119–139. [Google Scholar] [CrossRef]
- Goto, M. Citrus canker. In Plant Diseases of International Importance. Vol. III. Diseases of Fruit Crops; Kumar, J., Chaube, H.S., Singh, U.S., Mukhopadhyay, A.N., Eds.; Prentice Hall: Englewood Cliffs, NJ, USA, 1992; pp. 170–208. [Google Scholar]
- Ference, C.M.; Gochez, A.M.; Behlau, F.; Wang, N.; Graham, J.H.; Jones, J.B. Recent advances in the understanding of Xanthomonas citri ssp. citri pathogenesis and citrus canker disease management. Mol. Plant Pathol. 2018, 19, 1302. [Google Scholar]
- Das, A. Citrus canker-A review. J. Appl. Hortic. 2003, 5, 52–60. [Google Scholar] [CrossRef]
- Civerolo, E. Citrus bacterial canker disease: An overview. In Proceedings of the International Society of Citriculture, Okitsu, Japan, 9–12 November 1981; pp. 390–394. [Google Scholar]
- Behlau, F.; Gochez, A.M.; Jones, J.B. Diversity and copper resistance of Xanthomonas affecting citrus. Trop. Plant Pathol. 2020, 45, 200–212. [Google Scholar] [CrossRef]
- Smith, C.O.; Fawcett, H.S. A comparative study of the Citrus blast bacterium and some other allied organisms. J. Agric. Res. 1930, 41, 233–246. [Google Scholar]
- Mirik, M.; Baloglu, S.; Aysan, Y.; Cetinkaya-Yildiz, R.; Kusek, M.; Sahin, F. First outbreak and occurrence of citrus blast disease, caused by Pseudomonas syringae pv. syringae, on orange and mandarin trees in Turkey. Plant Pathol. 2005, 54, 238. [Google Scholar]
- Abdellatif, E.; Kałużna, M.; Ferrante, P.; Scortichini, M.; Bahri, B.; Janse, J.D.; van Vaerenberg, J.; Baeyen, S.; Sobiczewski, P.; Rhouma, A. Phylogenetic, genetic, and phenotypic diversity of Pseudomonas syringae pv. syringae strains isolated from citrus blast and black pit in Tunisia. Plant Pathol. 2020, 69, 1414–1425. [Google Scholar]
- Mougou, I. Citrus Blast and Black Pit Disease: A Review. DYSONA-Life Sci. 2022, 3, 1–6. [Google Scholar]
- Cazorla, F.M.; Arrebola, E.; Olea, F.; Velasco, L.; Hermoso, J.M.; Pérez-García, A.; Tores, J.A.; Farre, J.M.; de Vicente, A. Field evaluation of treatments for the control of the bacterial apical necrosis of mango (Mangifera indica) caused by Pseudomonas syringae pv. syringae. Eur. J. Plant Pathol. 2006, 116, 279–288. [Google Scholar] [CrossRef]
- Berk, Z. Citrus Fruit Processing; Academic Press: Cambridge, MA, USA, 2016. [Google Scholar]
- Lee, R.F. Control of virus diseases of citrus. Adv. Virus Res. 2015, 91, 143–173. [Google Scholar] [PubMed]
- Brown, G.; McCornack, A. Decay caused by Alternaria citri in Florida citrus fruit. Plant Dis. Report. 1972, 909–912. [Google Scholar]
- Troncoso-Rojas, R.; Tiznado-Hernández, M.E. Alternaria alternata (black rot, black spot). In Postharvest Decay; Elsevier: Amsterdam, The Netherlands, 2014; pp. 147–187. [Google Scholar]
- Vicent, A.; Armengol, J.; Sales, R.; García-Jiménez, J.; Alfaro-Lassala, F. First report of Alternaria brown spot of citrus in Spain. Plant Dis. 2000, 84, 1044. [Google Scholar] [CrossRef]
- Fagoaga, C.; Rodrigo, I.; Conejero, V.; Hinarejos, C.; Tuset, J.J.; Arnau, J.; Pina, J.A.; Navarro, L.; Peña, L. Increased tolerance to Phytophthora citrophthora in transgenic orange plants constitutively expressing a tomato pathogenesis related protein PR-5. Mol. Breed. 2001, 7, 175–185. [Google Scholar] [CrossRef]
- Bawage, S.; Nerkar, S.; Kumar, A.; Das, A. Morphological and molecular description of Phytophthora insolita isolated from citrus orchard in India. J. Mycol. 2013, 2013, 247951. [Google Scholar] [CrossRef]
- Das, A.; Nerkar, S.; Kumar, A.; Bawage, S. Detection, identification and characterization of Phytophthora spp. Infecting citrus in India. J. Plant Pathol. 2016, 98, 55–69. [Google Scholar]
- Smith, R.E.; Smith, E.H. A new fungus of economic importance. Bot. Gaz. 1906, 42, 215–221. [Google Scholar] [CrossRef]
- Rhaiem, A.; Taylor, P.W. Colletotrichum gloeosporioides associated with anthracnose symptoms on citrus, a new report for Tunisia. Eur. J. Plant Pathol. 2016, 146, 219–224. [Google Scholar] [CrossRef]
- Pérez-Mora, J.L.; Mora-Romero, G.A.; Beltrán-Peña, H.; García-León, E.; Lima, N.B.; Camacho-Tapia, M.; Tovar-Pedraza, J.M. First Report of Colletotrichum siamense and C. gloeosporioides Causing Anthracnose of Citrus spp. in Mexico. Plant Dis. 2021, 105, 496. [Google Scholar] [CrossRef] [PubMed]
- Palou, L. Penicillium digitatum, Penicillium italicum (green mold, blue mold). In Postharvest Decay; Elsevier: Amsterdam, The Netherlands, 2014; pp. 45–102. [Google Scholar]
- Papoutsis, K.; Mathioudakis, M.M.; Hasperué, J.H.; Ziogas, V. Non-chemical treatments for preventing the postharvest fungal rotting of citrus caused by Penicillium digitatum (green mold) and Penicillium italicum (blue mold). Trends Food Sci. Technol. 2019, 86, 479–491. [Google Scholar] [CrossRef]
- Suprapta, D.N.; Arai, K.; Iwai, H. Distribution of Geotrichum candidum citrus race in citrus groves and non-citrus fields in Japan. Mycoscience 1995, 36, 277–282. [Google Scholar] [CrossRef]
- McKay, A.H. Population Structure of the Sour Rot Pathogens Galactomyces citri-aurantii and G. geotrichum and Evaluation of Sterol Demethylation Inhibitors for Postharvest Management of Citrus Decays; University of California: Riverside, CA, USA, 2011. [Google Scholar]
- Nazerian, E.; Alian, Y.M. Association of Geotrichum citri-aurantii with citrus fruits decay in Iran. Int. J. Agron. Plant Prod. 2013, 4, 1839–1843. [Google Scholar]
- Palou, L. Control of citrus postharvest diseases by physical means. Tree For. Sci. Biotechnol. 2009, 3, 127–142. [Google Scholar]
- Timmer, L.W.; Akimitsu, K.; Solel, Z.; Peever, T. Alternaria diseases of citrus-novel pathosystems. Alternaria Dis. Citrus-Nov. Pathosyst. 2003, 42, 99–112. [Google Scholar]
- Carvalho, D.D.; Alves, E.; Batista, T.R.; Camargos, R.B.; Lopes, E.A. Comparison of methodologies for conidia production by Alternaria alternata from citrus. Braz. J. Microbiol. 2008, 39, 792–798. [Google Scholar] [CrossRef]
- Aiello, D.; Guarnaccia, V.; Azzaro, A.; Polizzi, G. Alternaria brown spot on new clones of sweet orange and lemon in Italy. Phytopathol. Mediterr. 2020, 59, 131–145. [Google Scholar]
- Varano, A.; Shirahigue, L.D.; Azevedo, F.A.; Altenhofen da Silva, M.; Ceccato-Antonini, S.R. Mandarin essential oil as an antimicrobial in ethanolic fermentation: Effects on Limosilactobacillus fermentum and Saccharomyces cerevisiae. Lett. Appl. Microbiol. 2022, 74, 981–991. [Google Scholar] [CrossRef] [PubMed]
- Sommano, S.; Joyce, D.; Dinh, S.; D’arcy, B. Infection by Alternaria alternata causes discolouration of Backhousia myrtifolia foliage and flowers. J. Hortic. Sci. Biotechnol. 2012, 87, 41–46. [Google Scholar] [CrossRef]
- Martin, F.N.; Abad, Z.G.; Balci, Y.; Ivors, K. Identification and detection of Phytophthora: Reviewing our progress, identifying our needs. Plant Dis. 2012, 96, 1080–1103. [Google Scholar] [CrossRef] [PubMed]
- Vitale, A.; Aiello, D.; Azzaro, A.; Guarnaccia, V.; Polizzi, G. An eleven-year survey on field disease susceptibility of citrus accessions to Colletotrichum and Alternaria species. Agriculture 2021, 11, 536. [Google Scholar] [CrossRef]
- Wang, W.; de Silva, D.D.; Moslemi, A.; Edwards, J.; Ades, P.K.; Crous, P.W.; Taylor, P.W. Colletotrichum species causing anthracnose of citrus in Australia. J. Fungi 2021, 7, 47. [Google Scholar] [CrossRef] [PubMed]
- Riolo, M.; Aloi, F.; Pane, A.; Cara, M.; Cacciola, S.O. Twig and shoot dieback of citrus, a new disease caused by Colletotrichum species. Cells 2021, 10, 449. [Google Scholar] [CrossRef]
- Uysal, A.; Kurt, Ş.; Guarnaccia, V. Distribution and characterization of Colletotrichum species associated with Citrus anthracnose in eastern Mediterranean region of Turkey. Eur. J. Plant Pathol. 2022, 163, 125–141. [Google Scholar] [CrossRef]
- Yang, R.; Miao, J.; Shen, Y.; Cai, N.; Wan, C.; Zou, L.; Chen, C.; Chen, J. Antifungal effect of cinnamaldehyde, eugenol and carvacrol nanoemulsion against Penicillium digitatum and application in postharvest preservation of citrus fruit. LWT 2021, 141, 110924. [Google Scholar] [CrossRef]
- Wang, Z.; Sui, Y.; Li, J.; Tian, X.; Wang, Q. Biological control of postharvest fungal decays in citrus: A review. Crit. Rev. Food Sci. Nutr. 2022, 62, 861–870. [Google Scholar] [CrossRef]
- Kanashiro, A.M.; Akiyama, D.Y.; Kupper, K.C.; Fill, T.P. Penicillium italicum: An underexplored postharvest pathogen. Front. Microbiol. 2020, 11, 606852. [Google Scholar] [CrossRef]
- Costa, J.H.; Bazioli, J.M.; de Moraes Pontes, J.G.; Fill, T.P. Penicillium digitatum infection mechanisms in citrus: What do we know so far? Fungal Biol. 2019, 123, 584–593. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Torres, P.; Tuset, J.J. Molecular insights into fungicide resistance in sensitive and resistant Penicillium digitatum strains infecting citrus. Postharvest Biol. Technol. 2011, 59, 159–165. [Google Scholar] [CrossRef]
- Boubaker, H.; Karim, H.; El Hamdaoui, A.; Msanda, F.; Leach, D.; Bombarda, I.; Vanloot, P.; Abbad, A.; Boudyach, E.; Aoumar, A.A.B. Chemical characterization and antifungal activities of four Thymus species essential oils against postharvest fungal pathogens of citrus. Ind. Crops Prod. 2016, 86, 95–101. [Google Scholar] [CrossRef]
- Zhao, J.; Zhang, D.; Wang, Z.; Tian, Z.; Yang, F.; Lu, X.; Long, C.-a. Genome sequencing and transcriptome analysis of Geotrichum citri-aurantii on citrus reveal the potential pathogenic-and guazatine-resistance related genes. Genomics 2020, 112, 4063–4071. [Google Scholar] [CrossRef]
- Guenther, E.; Althausen, D. The Essential Oils; Van Nostrand: New York, NY, USA, 1948; Volume 1. [Google Scholar]
- Prins, C.L.; Vieira, I.J.; Freitas, S.P. Growth regulators and essential oil production. Braz. J. Plant Physiol. 2010, 22, 91–102. [Google Scholar] [CrossRef]
- Tranchida, P.Q.; Bonaccorsi, I.; Dugo, P.; Mondello, L.; Dugo, G. Analysis of Citrus essential oils: State of the art and future perspectives. A review. Flavour Fragr. J. 2012, 27, 98–123. [Google Scholar] [CrossRef]
- Hussain, S.; Nawaz, H.; Ahmad, M.M.; Murtaza, M.A.; Rizvi, A.J. Inhibitory effect of citrus peel essential oils on the microbial growth of bread. Pak. J. Nutr. 2007, 6, 558–561. [Google Scholar]
- Rao, V.P.; Pandey, D. Extraction of Essential Oil and Its Applications. Bachelor degree in Technology (Chemical Engineering); Department of Chemical Engineering National Institute of Technology Rourkela: Orissa, India, 2012. [Google Scholar]
- Hanif, M.A.; Nisar, S.; Khan, G.S.; Mushtaq, Z.; Zubair, M. Essential oils. In Essential Oil Research; Springer: Berlin/Heidelberg, Germany, 2019; pp. 3–17. [Google Scholar]
- Saad, N.Y.; Muller, C.D.; Lobstein, A. Major bioactivities and mechanism of action of essential oils and their components. Flavour Fragr. J. 2013, 28, 269–279. [Google Scholar] [CrossRef]
- Perazzini, H.; Perazzini, M.T.; Freire, F.B.; Freire, F.B.; Freire, J.T. Modeling and cost analysis of drying of citrus residues as biomass in rotary dryer for bioenergy. Renew. Energy 2021, 175, 167–178. [Google Scholar] [CrossRef]
- Bhatti, H.N.; Bajwa, I.I.; Hanif, M.A.; Bukhari, I.H. Removal of lead and cobalt using lignocellulosic fiber derived from Citrus reticulata waste biomass. Korean J. Chem. Eng. 2010, 27, 218–227. [Google Scholar] [CrossRef]
- Porto, D.S.; Forim, M.R.; Costa, E.S.; Fernandes, J.B.; da Silva, M.F. Evaluation of lignins of trunk and roots from Citrus sinensis L. Osbeck: A large available Brazilian biomass. J. Braz. Chem. Soc. 2021, 32, 29–39. [Google Scholar] [CrossRef]
- Bruno, M.R.; Russo, D.; Cetera, P.; Faraone, I.; Lo Giudice, V.; Milella, L.; Todaro, L.; Sinisgalli, C.; Fritsch, C.; Dumarçay, S. Chemical analysis and antioxidant properties of orange-tree (Citrus sinensis L.) biomass extracts obtained via different extraction techniques. Biofuels Bioprod. Biorefin. 2020, 14, 509–520. [Google Scholar] [CrossRef]
- Raspo, M.A.; Vignola, M.B.; Andreatta, A.E.; Juliani, H.R. Antioxidant and antimicrobial activities of citrus essential oils from Argentina and the United States. Food Biosci. 2020, 36, 100651. [Google Scholar] [CrossRef]
- Ollitrault, P.; Curk, F.; Krueger, R. Citrus taxonomy. In The Genus Citrus; Elsevier: Amsterdam, The Netherlands, 2020; pp. 57–81. [Google Scholar]
- Sadka, A.; Shlizerman, L.; Kamara, I.; Blumwald, E. Primary metabolism in citrus fruit as affected by its unique structure. Front. Plant Sci. 2019, 10, 1167. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, M.M.; Rehman, S.U.; Anjum, F.M.; Bajwa, E.E. Comparative physical examination of various citrus peel essential oils. Int. J. Agric. Biol. 2006, 8, 186–190. [Google Scholar]
- Teigiserova, D.A.; Tiruta-Barna, L.; Ahmadi, A.; Hamelin, L.; Thomsen, M. A step closer to circular bioeconomy for citrus peel waste: A review of yields and technologies for sustainable management of essential oils. J. Environ. Manag. 2021, 280, 111832. [Google Scholar] [CrossRef]
- Maurya, A.K.; Mohanty, S.; Pal, A.; Chanotiya, C.S.; Bawankule, D.U. The essential oil from Citrus limetta Risso peels alleviates skin inflammation: In-vitro and in-vivo study. J. Ethnopharmacol. 2018, 212, 86–94. [Google Scholar] [CrossRef]
- Mahato, N.; Sharma, K.; Koteswararao, R.; Sinha, M.; Baral, E.; Cho, M.H. Citrus essential oils: Extraction, authentication and application in food preservation. Crit. Rev. Food Sci. Nutr. 2019, 59, 611–625. [Google Scholar] [CrossRef]
- Javed, S.; Javaid, A.; Nawaz, S.; Saeed, M.; Mahmood, Z.; Siddiqui, S.; Ahmad, R. Phytochemistry, GC-MS analysis, antioxidant and antimicrobial potential of essential oil from five citrus species. J. Agric. Sci. 2014, 6, 201. [Google Scholar] [CrossRef]
- Chandharakool, S.; Koomhin, P.; Sinlapasorn, J.; Suanjan, S.; Phungsai, J.; Suttipromma, N.; Songsamoe, S.; Matan, N.; Sattayakhom, A. Effects of Tangerine Essential Oil on Brain Waves, Moods, and Sleep Onset Latency. Molecules 2020, 25, 4865. [Google Scholar] [CrossRef]
- Persistence Market Research. Citrus Oil Market Outlook (2021–2031). Available online: https://www.persistencemarketresearch.com/market-research/citrus-oil-market.asp (accessed on 11 July 2022).
- Palazzolo, E.; Laudicina, V.A.; Germanà, M.A. Current and potential use of citrus essential oils. Curr. Org. Chem. 2013, 17, 3042–3049. [Google Scholar] [CrossRef]
- Mahato, N.; Sharma, K.; Sinha, M.; Cho, M.H. Citrus waste derived nutra-/pharmaceuticals for health benefits: Current trends and future perspectives. J. Funct. Foods 2018, 40, 307–316. [Google Scholar] [CrossRef]
- Belkheiri, A.; Forouhar, A.; Ursu, A.V.; Dubessay, P.; Pierre, G.; Delattre, C.; Djelveh, G.; Abdelkafi, S.; Hamdami, N.; Michaud, P. Extraction, characterization, and applications of pectins from plant by-products. Appl. Sci. 2021, 11, 6596. [Google Scholar] [CrossRef]
- Mesbahi, G.; Jamalian, J.; Farahnaky, A. A comparative study on functional properties of beet and citrus pectins in food systems. Food Hydrocoll. 2005, 19, 731–738. [Google Scholar] [CrossRef]
- Willats, W.G.; Knox, J.P.; Mikkelsen, J.D. Pectin: New insights into an old polymer are starting to gel. Trends Food Sci. Technol. 2006, 17, 97–104. [Google Scholar] [CrossRef]
- Hosseini, S.S.; Khodaiyan, F.; Kazemi, M.; Najari, Z. Optimization and characterization of pectin extracted from sour orange peel by ultrasound assisted method. Int. J. Biol. Macromol. 2019, 125, 621–629. [Google Scholar] [CrossRef]
- Thakur, B.R.; Singh, R.K.; Handa, A.K.; Rao, M. Chemistry and uses of pectin—a review. Crit. Rev. Food Sci. Nutr. 1997, 37, 47–73. [Google Scholar] [CrossRef]
- Ciriminna, R.; Fidalgo, A.; Delisi, R.; Ilharco, L.M.; Pagliaro, M. Pectin production and global market. Agro Food Ind. Hi-Tech 2016, 27, 17–20. [Google Scholar]
- Kanmani, P.; Dhivya, E.; Aravind, J.; Kumaresan, K. Extraction and analysis of pectin from citrus peels: Augmenting the yield from Citrus limon using statistical experimental design. Iran. J. Energy Environ. 2014, 5, 81–84. [Google Scholar] [CrossRef]
- Bagde, P.P.; Dhenge, S.; Bhivgade, S. Extraction of pectin from orange peel and lemon peel. Int. J. Eng. Technol. Sci. Res. 2017, 4, 1–7. [Google Scholar]
- Ververis, C.; Georghiou, K.; Danielidis, D.; Hatzinikolaou, D.; Santas, P.; Santas, R.; Corleti, V. Cellulose, hemicelluloses, lignin and ash content of some organic materials and their suitability for use as paper pulp supplements. Bioresour. Technol. 2007, 98, 296–301. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Labrador, A.; Moreno, R.; Villamiel, M.; Montilla, A. Preparation of citrus pectin gels by power ultrasound and its application as an edible coating in strawberries. J. Sci. Food Agric. 2018, 98, 4866–4875. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Widmer, W.; Grohmann, K. Economic analysis of ethanol production from citrus peel waste. Proc. Fla. State Hortic. Soc. 2007, 120, 310–315. [Google Scholar]
- Ali, A.; Ambreen, S.; Maqbool, Q.; Naz, S.; Shams, M.F.; Ahmad, M.; Phull, A.R.; Zia, M. Zinc impregnated cellulose nanocomposites: Synthesis, characterization and applications. J. Phys. Chem. Solids 2016, 98, 174–182. [Google Scholar] [CrossRef]
- Jhalegar, M.J.; Sharma, R.; Singh, D. In vitro and in vivo activity of essential oils against major postharvest pathogens of Kinnow (Citrus nobilis× C. deliciosa) mandarin. J. Food Sci. Technol. 2015, 52, 2229–2237. [Google Scholar] [CrossRef]
- Ghidelli, C.; Pérez-Gago, M.B. Recent advances in modified atmosphere packaging and edible coatings to maintain quality of fresh-cut fruits and vegetables. Crit. Rev. Food Sci. Nutr. 2018, 58, 662–679. [Google Scholar] [CrossRef]
- Prasad, K.; Guarav, A.K.; Preethi, P.; Neha, P. Edible coating technology for extending market life of horticultural produce. Acta Sci. Agric. 2018, 2, 55–64. [Google Scholar]
- Sapper, M.; Chiralt, A. Starch-based coatings for preservation of fruits and vegetables. Coatings 2018, 8, 152. [Google Scholar] [CrossRef]
- Hasan, S.K.; Ferrentino, G.; Scampicchio, M. Nanoemulsion as advanced edible coatings to preserve the quality of fresh-cut fruits and vegetables: A review. Int. J. Food Sci. Technol. 2020, 55, 1–10. [Google Scholar] [CrossRef]
- Panahirad, S.; Dadpour, M.; Peighambardoust, S.H.; Soltanzadeh, M.; Gullón, B.; Alirezalu, K.; Lorenzo, J.M. Applications of carboxymethyl cellulose-and pectin-based active edible coatings in preservation of fruits and vegetables: A review. Trends Food Sci. Technol. 2021, 110, 663–673. [Google Scholar] [CrossRef]
- Hassan, B.; Chatha, S.A.S.; Hussain, A.I.; Zia, K.M.; Akhtar, N. Recent advances on polysaccharides, lipids and protein based edible films and coatings: A review. Int. J. Biol. Macromol. 2018, 109, 1095–1107. [Google Scholar] [CrossRef] [PubMed]
- Arnon-Rips, H.; Poverenov, E. Improving food products’ quality and storability by using Layer by Layer edible coatings. Trends Food Sci. Technol. 2018, 75, 81–92. [Google Scholar] [CrossRef]
- Šuput, D.Z.; Lazić, V.L.; Popović, S.Z.; Hromiš, N.M. Edible films and coatings: Sources, properties and application. Food Feed Res. 2015, 42, 11–22. [Google Scholar] [CrossRef]
- Malmiri, J.H.; Osman, A.; Tan, C.; Rahman, A.R. Evaluation of effectiveness of three cellulose derivative-based edible coatings on changes of physico-chemical characteristics of’Berangan’banana (Musa sapientum cv. Berangan) during storage at ambient conditions. Int. Food Res. J. 2011, 18, 1381. [Google Scholar]
- Lara-Espinoza, C.; Carvajal-Millán, E.; Balandrán-Quintana, R.; López-Franco, Y.; Rascón-Chu, A. Pectin and pectin-based composite materials: Beyond food texture. Molecules 2018, 23, 942. [Google Scholar] [CrossRef]
- Menezes, J.; Athmaselvi, K. Study on effect of pectin based edible coating on the shelf life of sapota fruits. Biosci. Biotechnol. Res. Asia 2016, 13, 1195–1199. [Google Scholar] [CrossRef]
- Valdés, A.; Burgos, N.; Jiménez, A.; Garrigós, M.C. Natural pectin polysaccharides as edible coatings. Coatings 2015, 5, 865–886. [Google Scholar] [CrossRef]
- Palou, L.; Valencia-Chamorro, S.A.; Pérez-Gago, M.B. Antifungal edible coatings for fresh citrus fruit: A review. Coatings 2015, 5, 962–986. [Google Scholar] [CrossRef]
- Alvarez, M.V.; Taberner, V.; Fernández-Catalán, A.; Martínez-Blay, V.; Argente-Sanchis, M.; Palou, L.; Pérez-Gago, M.B. Natural pectin-based edible composite coatings with antifungal properties to control postharvest decay and reduce losses of ‘valencia’oranges. In Proceedings of the First International Conference in Sustainable Food Packaging, online, 26–29 September 2021. [Google Scholar]
- De Azeredo, H.; Rosa, M.; De Sá, M.; Souza Filho, M.; Waldron, K. The use of biomass for packaging films and coatings. In Advances in Biorefineries; Elsevier: Amsterdam, The Netherlands, 2014; pp. 819–874. [Google Scholar]
- Barhoum, A.; Jeevanandam, J.; Rastogi, A.; Samyn, P.; Boluk, Y.; Dufresne, A.; Danquah, M.K.; Bechelany, M. Plant celluloses, hemicelluloses, lignins, and volatile oils for the synthesis of nanoparticles and nanostructured materials. Nanoscale 2020, 12, 22845–22890. [Google Scholar] [CrossRef]
- Alvarez, M.V.; Palou, L.; Taberner, V.; Fernández-Catalán, A.; Argente-Sanchis, M.; Pitta, E.; Pérez-Gago, M.B. Natural Pectin-Based Edible Composite Coatings with Antifungal Properties to Control Green Mold and Reduce Losses of ‘Valencia’ Oranges. Foods 2022, 11, 1083. [Google Scholar] [CrossRef]
- Jafarzadeh, S.; Nafchi, A.M.; Salehabadi, A.; Oladzad-Abbasabadi, N.; Jafari, S.M. Application of bio-nanocomposite films and edible coatings for extending the shelf life of fresh fruits and vegetables. Adv. Colloid Interface Sci. 2021, 291, 102405. [Google Scholar] [CrossRef] [PubMed]
- Kocira, A.; Kozłowicz, K.; Panasiewicz, K.; Staniak, M.; Szpunar-Krok, E.; Hortyńska, P. Polysaccharides as edible films and coatings: Characteristics and influence on fruit and vegetable quality—A review. Agronomy 2021, 11, 813. [Google Scholar] [CrossRef]
- Hou, Z.; Chen, S.; Ye, X. High pressure processing accelarated the release of RG-I pectic polysaccharides from citrus peel. Carbohydr. Polym. 2021, 263, 118005. [Google Scholar] [CrossRef] [PubMed]
- Park, H.-R.; Lee, S.J.; Im, S.-B.; Shin, M.-S.; Choi, H.-J.; Park, H.-Y.; Shin, K.-S. Signaling pathway and structural features of macrophage-activating pectic polysaccharide from Korean citrus, Cheongkyool peels. Int. J. Biol. Macromol. 2019, 137, 657–665. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khamsaw, P.; Sangta, J.; Chaiwan, P.; Rachtanapun, P.; Sirilun, S.; Sringarm, K.; Thanakkasaranee, S.; Sommano, S.R. Bio-Circular Perspective of Citrus Fruit Loss Caused by Pathogens: Occurrences, Active Ingredient Recovery and Applications. Horticulturae 2022, 8, 748. https://doi.org/10.3390/horticulturae8080748
Khamsaw P, Sangta J, Chaiwan P, Rachtanapun P, Sirilun S, Sringarm K, Thanakkasaranee S, Sommano SR. Bio-Circular Perspective of Citrus Fruit Loss Caused by Pathogens: Occurrences, Active Ingredient Recovery and Applications. Horticulturae. 2022; 8(8):748. https://doi.org/10.3390/horticulturae8080748
Chicago/Turabian StyleKhamsaw, Pattarapol, Jiraporn Sangta, Pirawan Chaiwan, Pornchai Rachtanapun, Sasithorn Sirilun, Korawan Sringarm, Sarinthip Thanakkasaranee, and Sarana Rose Sommano. 2022. "Bio-Circular Perspective of Citrus Fruit Loss Caused by Pathogens: Occurrences, Active Ingredient Recovery and Applications" Horticulturae 8, no. 8: 748. https://doi.org/10.3390/horticulturae8080748