
Balance of Carotenoid Synthesis and Degradation Contributes to the Color Difference between Chinese Narcissus and Its Yellow-Tepal Mutant

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Pigment Semiquantification
2.3. Total RNA Extraction and RNA-Seq Analysis
2.4. Identification of Differentially Expressed Genes
2.5. Quantitative Real-Time PCR
3. Results
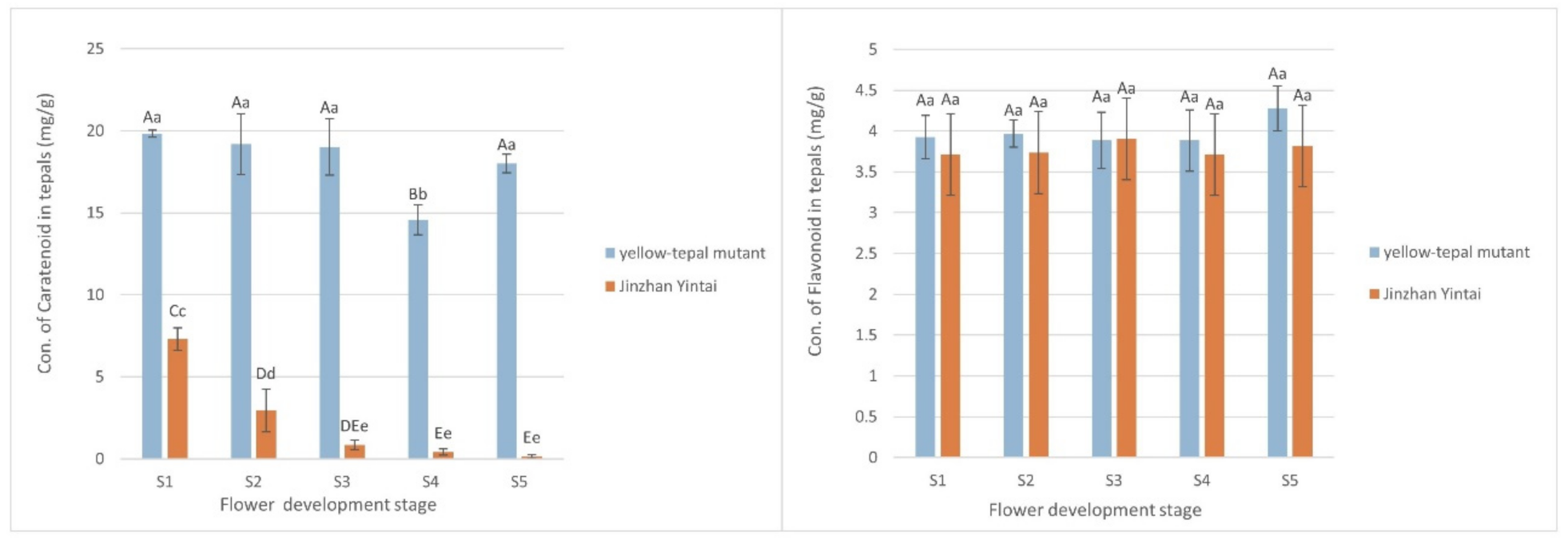
3.1. Tepal Pigmentation Characteristics of Chinese Narcissus
3.2. Tepal Transcriptome Sequencing, Assembly and Annotation
3.3. Expression Profiles of DEGs in Tepals
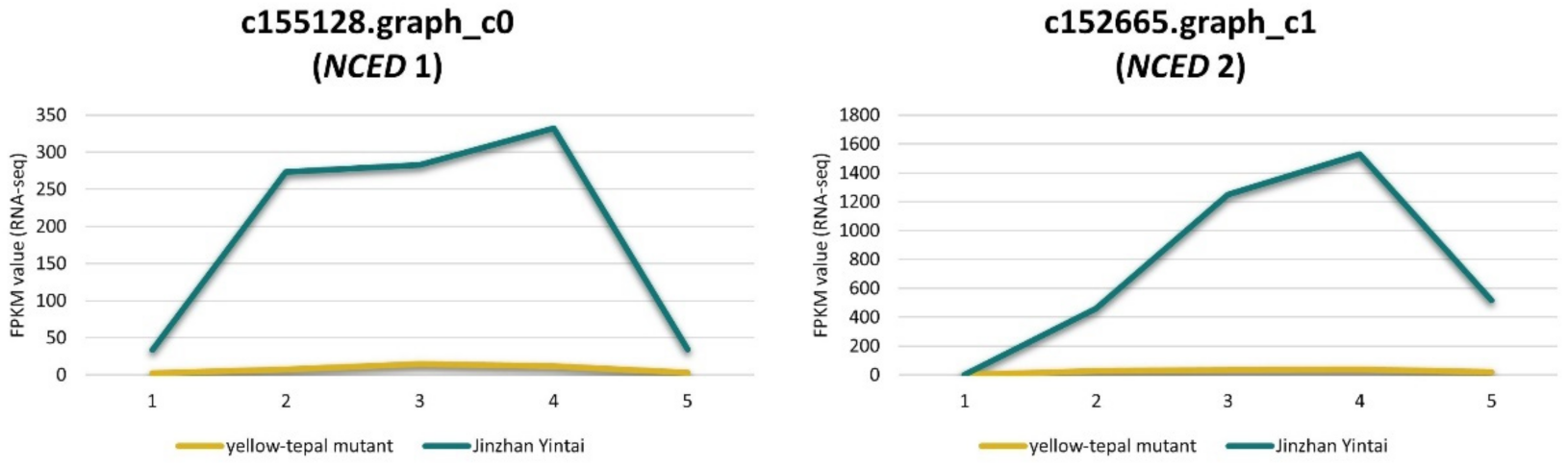
3.4. DEGs Involved in Carotenoid Biosynthesis
3.5. Transcription Factors Involved in Narcissus Tepal Pigmentation
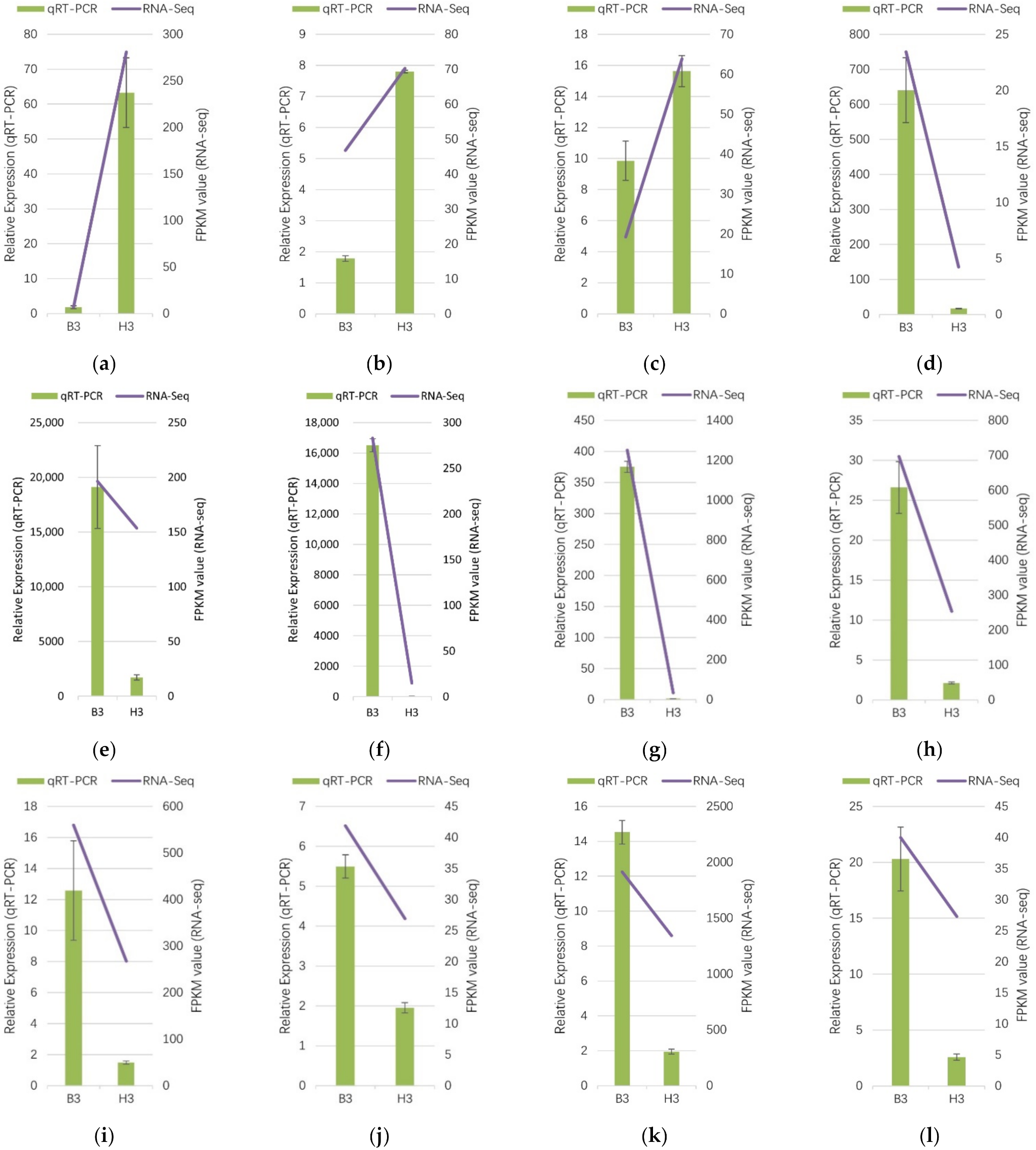
3.6. Verification of Gene Expression by qRT-PCR
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ren, Y.; Liu, C.; Li, X. Comparison of karyotype between univalve Chinese narcissus and multiple-valve Chinese narcissus. J. Agriculyural Univ. Hebei 2010, 33, 9–12. [Google Scholar]
- Liu, Y.; Song, X.; Zhang, D.; Zhou, F.; Wang, D.; Wei, Y. Blueberry anthocyanins: Protection against ageing and light-induced damage in retinal pigment epithelial cells. Br. J. Nutr. 2012, 108, 16–27. [Google Scholar] [CrossRef] [PubMed]
- Becker, A.; Alix, K.; Damerval, C. The evolution of flower development: Current understanding and future challenges. Ann. Bot. 2011, 107, 1427–1431. [Google Scholar] [CrossRef] [PubMed]
- Valadon, L.R.; Mummery, R.S. Carotenoids in floral parts of a narcissus, a daffodil and a tulip. Biochem. J. 1968, 106, 479–484. [Google Scholar] [CrossRef] [PubMed]
- Beyer, P. Carotene biosynthesis in daffodil chromoplasts: On the membrane-integral desaturation and cyclization reactions. Curr. Top. Plant Physiol. 1989, 2, 157–170. [Google Scholar]
- Li, X.; Lu, M.; Tang, D.; Shi, Y. Composition of Carotenoids and Flavonoids in Narcissus Cultivars and their Relationship with Flower Color. PLoS ONE 2015, 10, e0142074. [Google Scholar] [CrossRef]
- Sun, T.; Rao, S.; Zhou, X. Plant carotenoids: Recent advances and future perspectives. Mol. Hortic. 2022, 2, 3. [Google Scholar] [CrossRef]
- Krinsky, N.I.; Johnson, E.J. Carotenoid actions and their relation to health and disease. Mol. Asp. Med. 2005, 26, 459–516. [Google Scholar] [CrossRef]
- Nisar, N.; Li, L.; Lu, S.; Khin, N.C.; Pogson, B.J. Carotenoid metabolism in plants. Mol. Plant. 2015, 8, 68–82. [Google Scholar] [CrossRef]
- Rodriguez-Concepcion, M.; Avalos, J.; Bonet, M.L.; Boronat, A.; Gomez-Gomez, L.; Hornero-Mendez, D. A global perspective on carotenoids: Metabolism, biotechnology, and benefits for nutrition and health. Prog. Lipid Res. 2018, 70, 62–93. [Google Scholar] [CrossRef]
- Sun, T.; Yuan, H.; Cao, H.; Yazdani, M.; Tadmor, Y.; Li, L. Carotenoid Metabolism in Plants: The Role of Plastids. Mol. Plant 2018, 11, 58–74. [Google Scholar] [CrossRef] [PubMed]
- Wurtzel, E.T. Changing Form and Function through Carotenoids and Synthetic Biology. Plant Physiol. 2019, 179, 830–843. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Yuan, H.; Zeng, Y.; Xu, Q. Plastids and Carotenoid Accumulation. Carotenoids Nat. 2016, 79, 273–293. [Google Scholar]
- Sun, T.; Tadmor, Y.; Li, L. Pathways for Carotenoid Biosynthesis, Degradation, and Storage. In Plant and Food Carotenoids; Rodríguez-Concepción, M., Welsch, R., Eds.; Humana: New York, NY, USA, 2020; Volume 2083, pp. 3–23. [Google Scholar]
- Yuan, H.; Zhang, J.; Nageswaran, D.; Li, L. Carotenoid metabolism and regulation in horticultural crops. Hortic. Res. 2015, 2, 15036. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.C.; Molnár, P.; Schwab, W. Cloning and functional characterization of carotenoid cleavage dioxygenase 4 genes. J. Exp. Bot. 2009, 60, 3011–3022. [Google Scholar] [CrossRef]
- Schwartz, S.H.; Tan, B.C.; Gage, D.A.; Zeevaart, J.A.; McCarty, D.R. Specific oxidative cleavage of carotenoids by VP14 of maize. Science 1997, 276, 1872–1874. [Google Scholar] [CrossRef]
- Tan, B.C.; Joseph, L.M.; Deng, W.T.; Liu, L.; Li, Q.B.; Cline, K. Molecular characterization of the Arabidopsis 9-cis epoxycarotenoid dioxygenase gene family. Plant J. 2003, 35, 44–56. [Google Scholar] [CrossRef]
- Wang, J.Y.; Haider, I.; Jamil, M.; Fiorilli, V.; Saito, Y.; Mi, J. The apocarotenoid metabolite zaxinone regulates growth and strigolactone biosynthesis in rice. Nat. Commun. 2019, 10, 810. [Google Scholar] [CrossRef]
- Ablazov, A.; Mi, J.; Jamil, M.; Jia, K.P.; Wang, J.Y.; Feng, Q. The Apocarotenoid Zaxinone Is a Positive Regulator of Strigolactone and Abscisic Acid Biosynthesis in Arabidopsis Roots. Front. Plant Sci. 2020, 11, 578. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Tang, D.; Du, H.; Shi, Y.H. Transcriptome sequencing and biochemical analysis of perianths and coronas reveal flower color formation in narcissus pseudonarcissus. Int. J. Mol. Sci. 2018, 19, 4006. [Google Scholar] [CrossRef] [PubMed]
- Stanley, L.; Yuan, Y.W. Transcriptional Regulation of Carotenoid Biosynthesis in Plants: So Many Regulators, So Little Consensus. Front. Plant Sci. 2019, 10, 1017. [Google Scholar] [CrossRef] [PubMed]
- Hermanns, A.S.; Zhou, X.S.; Xu, Q.; Tadmor, Y.; Li, L. Carotenoid Pigment Accumulation in Horticultural Plants. Hortic. Plant J. 2020, 6, 343–360. [Google Scholar] [CrossRef]
- Afonso, T.; Moresco, R.; Uarrota, V.G.; Navarro, B.B.; Nunes, E.D.C.; Maraschin, M.; Rocha, M. UV-Vis and CIELAB Based Chemometric Characterization of Manihot esculenta Carotenoid Contents. J. Integr. Bioinform. 2017, 14, 20170056. [Google Scholar] [CrossRef] [PubMed]
- Cheng, B.X.; Wan, H.H.; Han, Y.; Yu, C.; Luo, L.; Pan, H.T.; Zhang, Q.X. Identification and QTL Analysis of Flavonoids and Carotenoids in Tetraploid Roses Based on an Ultra-High-Density Genetic Map. Front. Plant Sci. 2021, 12, 1071. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Morikawa, T.; Uraguchi, Y.; Sanda, S. Overexpression of DnaJ-Like Chaperone Enhances Carotenoid Synthesis in Chlamydomonas reinhardtii. Appl. Biochem. Biotechnol. 2017, 184, 80–91. [Google Scholar] [CrossRef]
- Schledz, M.; Al-Babili, S.; von Lintig, J.; Haubruck, H.; Rabbani, S.; Kleinig, H.; Beyer, P. Phytoene synthase from Narcissus pseudonarcissus: Functional expression, galactolipid requirement, topological distribution in chromoplasts and induction during flowering. Plant J. Cell Mol. Biol. 1996, 10, 781–792. [Google Scholar] [CrossRef]
- Welsch, R.; Zhou, X.; Yuan, H.; Alvarez, D.; Sun, T.; Schlossarek, D.; Li, L. Clp protease and OR directly control the proteostasis of phytoene synthase, the crucial enzyme for carotenoid biosynthesis in arabidopsis. Mol. Plant 2018, 11, 149–162. [Google Scholar] [CrossRef]
- Xu, M.; Liu, C.L.; Luo, J.; Qi, Z.; Yan, Z.; Fu, Y.; Tang, H. Transcriptomic de novo analysis of pitaya (Hylocereus polyrhizus) canker disease caused by Neoscytalidium dimidiatum. BMC Genom. 2019, 20, 10. [Google Scholar] [CrossRef]
- Yamasaki, K.; Kigawa, T.; Seki, M.; Shinozaki, K.; Yokoyama, S. DNA-binding domains of plant-specific transcription factors: Structure, function, and evolution. Trends Plant Sci. 2013, 18, 267–276. [Google Scholar] [CrossRef] [PubMed]
- Lutova, L.A.; Dodueva, I.E.; Lebedeva, M.A.; Tvorogova, V.E. Transcription factors in developmental genetics and the evolution of higher plants. Genetika 2015, 51, 539–557. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.S.; Chen, L.J.; Yang, Q.L.; Hu, Z.L.; Guo, P.Y.; Xie, Q.L.; Chen, G.P. New insight into the pigment composition and molecular mechanism of flower coloration in tulip (Tulipa gesneriana L.) cultivars with various petal colors. Plant Sci. 2022, 317, 111193. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Xu, M.; Li, R. Transcriptomic investigation of the basis of Corona and petal colour in Chinese narcissus. J. Hortic. Sci. Biotechnol. 2020, 95, 565–577. [Google Scholar] [CrossRef]
- Yan, J.; Wang, M.; Zhang, L. Light induces petal color change in Quisqualis indica (Combretaceae). Plant Divers. 2018, 40, 28–34. [Google Scholar] [CrossRef]
- Toledo-Ortiz, G.; Huq, E.; Rodríguez-Concepción, M. Direct regulation of phytoene synthase gene expression and carotenoid biosynthesis by phytochrome-interacting factors. Proc. Natl. Acad. Sci. USA 2010, 107, 11626–11631. [Google Scholar] [CrossRef]
- Zou, L.P.; Gao, H.P.; Zhong, Y.Q. Construction of Overexpression Vector for Phytoene Dehydrogenase Gene and Its Expression Identification in Tomato. Hubei Agric. Sci. 2012, 51, 393–395. [Google Scholar]
- Qi, Y.W.; Wu, H.X.; Liu, J.; Chen, L.J.; Jiang, Z.T.; Zhang, Y.J.; Tian, X.T.; Li, R.; Yang, Y.; Ren, X.L. Lycopene β-cyclase plays a critical role in carotenoid biosynthesis during persimmon fruit development and postharvest ripening. Sci. Hortic. 2021, 287, 110265. [Google Scholar] [CrossRef]
- Isaacson, T.; Ronen, G.; Hirschberg, Z.J. Cloning of tangerine from tomato reveals a carotenoid isomerase essential for the production of beta-carotene and xanthophylls in plants. Plant Cell 2002, 14, 333–342. [Google Scholar] [CrossRef]
- Zhang, X.; Dong, X.; Song, X. Photoperiodic Responses and Characterization of the Cmvvd Gene Encoding a Blue Light Photoreceptor from the Medicinal Caterpillar Fungus Cordyceps militaris (Ascomycetes). Int. J. Med. Mushrooms 2017, 19, 163. [Google Scholar] [CrossRef]
- Moreno, J.C.; Pizarro, L.; Fuentes, P. Levels of Lycopene β-Cyclase 1 Modulate Carotenoid Gene Expression and Accumulation in Daucus carota. PLoS ONE 2013, 8, e58144. [Google Scholar]
- Wu, X.; Zhu, L.; Li, O. Less Dynamic Sphingomonas Bacteria crtZ Cell Gene, crtG Gene and Its Application. CN Patent CN102286495 A, 2011. [Google Scholar]
- Petry, F.C.; Nadai, F.D.; Cristofani-Yaly, M. Carotenoid biosynthesis and quality characteristics of new hybrids between tangor (Citrus reticulata × C. sinensis) cv. ‘Murcott’ and sweet orange (C. sinensis) cv. ‘Pêra’. Food Res. Int. 2019, 122, 461–470. [Google Scholar] [CrossRef]
- Arumingtyas, E.L.; Fuadati, A.Z.; Dwinianti, E.F. Study on the Profile of Capsanthin-Capsurobin Synthase (Ccs) Gene responsible for Carotenoid Synthesis in Chili Pepper (Capsicum frutescens L.) Mutants G1M6 M2 Generation. IOP Conf. Ser. Earth Environ. Sci. 2019, 391, 012019. [Google Scholar] [CrossRef]
- Jiang, J.; Ma, S.; Ye, N.; Jiang, M.; Cao, J.; Zhang, J. WRKY transcription factors in plant responses to stresses. J. Integr. Plant Biol. 2017, 59, 86–101. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Yang, H.; Liu, H.; Yang, J.; Hu, Y. Extensive transcriptome changes underlying the flower c intensity variation in Paeonia ostii. Front. Plant Sci. 2015, 6, 1205. [Google Scholar] [CrossRef] [PubMed]
- Feng, S.; Sun, S.; Chen, X.; Wu, S.; Wang, D.; Chen, X. PyMYB10 and PyMYB10.1 Interact with bHLH to enhance anthocyanin accumulation in Pears. PLoS ONE 2015, 10, e0142112. [Google Scholar] [CrossRef]
- Quattrocchio, F.; Verweij, W.; Kroon, A.; Spelt, C.; Mol, J.; Koes, R. PH4 of Petunia is an R2R3 MYB protein that activates vacuolar acidification through interactions with basic-helix-loop-helix transcription factors of the anthocyanin pathway. Plant Cell 2006, 18, 1274–1291. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| # Anno_Database | Annotated_Number | 300 ≤ Length < 1000 | Length ≥ 1000 |
|---|---|---|---|
| COG_Annotation | 8713 | 1932 | 6112 |
| GO_Annotation | 18,219 | 5610 | 10,598 |
| KEGG_Annotation | 11,156 | 3425 | 6524 |
| KOG_Annotation | 18,928 | 5548 | 11,215 |
| Pfam_Annotation | 20,011 | 4961 | 13,657 |
| Swissprot_Annotation | 19,326 | 5713 | 12,008 |
| eggNOG_Annotation | 29,824 | 9238 | 17,439 |
| nr_Annotation | 31,761 | 10,221 | 18,147 |
| All_Annotated | 32,816 | 10,642 | 18,345 |
| Number of DEGs | |||
|---|---|---|---|
| Comparison | Total | Up-Regulated | Down-Regulated |
| H1 vs. B1 | 4512 | 2744 | 1768 |
| H2 vs. B2 | 5301 | 3147 | 2154 |
| H3 vs. B3 | 5648 | 3264 | 2384 |
| H4 vs. B4 | 4732 | 2671 | 2061 |
| H5 vs. B5 | 4836 | 2629 | 2207 |
| Function | Gene | No. Unigene | |
|---|---|---|---|
| Yellow | White | ||
| MEP pathway | DXS | 7 | 7 |
| DXR | 2 | 2 | |
| MCT | 1 | 1 | |
| CMK | 1 | 1 | |
| MDS | 1 | 1 | |
| HDS | 1 | 3 | |
| HDR | 1 | 1 | |
| IPPI | 1 | 1 | |
| Carotenoid degradation | D27 | 2 | 2 |
| CCD8 | 2 | 2 | |
| ABA2 | 1 | 1 | |
| NCED | 3 | 5 | |
| AAO3 | 4 | 4 | |
| CYP707A | 5 | 3 | |
| Carotenoid biosynthesis | GGPPS | 5 | 5 |
| PSY | 4 | 5 | |
| PDS | 2 | 2 | |
| Z-ISO | 1 | 1 | |
| ZDS | 1 | 1 | |
| CRTISO | 1 | 1 | |
| LCY-ε | 1 | 1 | |
| LCY-β | 1 | 1 | |
| CRTZ | 2 | 2 | |
| CYP97A3 | 2 | 2 | |
| CYP97C1 | 1 | 1 | |
| BCH | 2 | 2 | |
| ZEP | 2 | 1 | |
| VDE | 1 | 1 | |
| CCS | 1 | 1 | |
| TF Family | Number of Genes Encoding TFs | Gene ID |
|---|---|---|
| WRKY | 36 | c120255.graph_c0,c138929.graph_c0,c142805.graph_c1,c144839.graph_c1,c149972.graph_c3,c110362.graph_c0,c124494.graph_c1,c129251.graph_c0,c130489.graph_c0,c131406.graph_c0,c131659.graph_c0,c133316.graph_c0,c137603.graph_c0,c137677.graph_c0,c139214.graph_c1,c139631.graph_c0,c139892.graph_c0,c140033.graph_c0,c140154.graph_c0,c142031.graph_c0,c143733.graph_c0,c144189.graph_c0,c144839.graph_c0,c144888.graph_c0,c145392.graph_c0,c146928.graph_c0,c147917.graph_c0,c148304.graph_c0,c150502.graph_c0,c151842.graph_c0,c152031.graph_c0,c153038.graph_c0,c153300.graph_c0,c153389.graph_c0,c155651.graph_c0,c155651.graph_c1 |
| Ethylene-related | 35 | c107037.graph_c0,c112846.graph_c0,c120537.graph_c1,c125725.graph_c0,c126949.graph_c0,c127348.graph_c1,c128898.graph_c0,c129027.graph_c0,c129751.graph_c0,c130471.graph_c0,c131179.graph_c0,c135333.graph_c1,c135717.graph_c0,c136264.graph_c0,c136742.graph_c0,c139514.graph_c1,c141546.graph_c0,c141972.graph_c0,c144698.graph_c0,c144956.graph_c0,c145381.graph_c0,c145740.graph_c0,c145780.graph_c0,c145804.graph_c0,c146869.graph_c1,c148131.graph_c0,c148462.graph_c0,c148623.graph_c3,c150790.graph_c0,c150997.graph_c0,c151480.graph_c0,c152024.graph_c0,c152045.graph_c1,c154630.graph_c0,c154679.graph_c0 |
| bHLH | 34 | c101571.graph_c0,c110725.graph_c0,c113899.graph_c0,c116420.graph_c0,c116420.graph_c1,c117084.graph_c0,c119738.graph_c0,c128396.graph_c0,c133455.graph_c0,c135719.graph_c0,c139052.graph_c0,c139096.graph_c0,c140371.graph_c0,c140801.graph_c0,c141137.graph_c1,c141908.graph_c0,c142597.graph_c0,c142892.graph_c1,c143923.graph_c0,c144843.graph_c0,c146133.graph_c0,c147563.graph_c1,c147745.graph_c0,c148127.graph_c0,c148475.graph_c1,c148812.graph_c0,c149767.graph_c1,c150152.graph_c1,c150167.graph_c1,c151829.graph_c1,c151905.graph_c0,c154012.graph_c0,c154152.graph_c0,c154210.graph_c0 |
| MYB/MYB-related | 32 | c101588.graph_c0,c102290.graph_c0,c112191.graph_c0,c117005.graph_c0,c124182.graph_c0,c126634.graph_c1,c127645.graph_c0,c133547.graph_c0,c134396.graph_c0,c134396.graph_c1,c134934.graph_c0,c136246.graph_c0,c140058.graph_c1,c141363.graph_c0,c141774.graph_c0,c142114.graph_c0,c142462.graph_c0,c143527.graph_c0,c144577.graph_c0,c144693.graph_c2,c145610.graph_c0,c145904.graph_c0,c145968.graph_c0,c147959.graph_c0,c149164.graph_c0,c149369.graph_c0,c149914.graph_c1,c150703.graph_c1,c152921.graph_c1,c154714.graph_c1,c154951.graph_c0,c155785.graph_c0 |
| GATA | 11 | c115991.graph_c0,c129217.graph_c0,c129697.graph_c0,c133305.graph_c0,c133774.graph_c0,c143087.graph_c1,c146337.graph_c1,c149095.graph_c0,c151207.graph_c0,c154092.graph_c0,c155538.graph_c0 |
| NAC | 10 | c135189.graph_c0,c135248.graph_c0,c135248.graph_c1,c142337.graph_c0,c144310.graph_c0,c148978.graph_c0,c149737.graph_c0,c149737.graph_c1,c151067.graph_c1,c151067.graph_c2 |
| Heat Stress Related | 8 | c101690.graph_c0,c128110.graph_c0,c139765.graph_c0,c139765.graph_c1,c145598.graph_c0,c145840.graph_c0,c153258.graph_c0,c153504.graph_c0 |
| MADS | 6 | c101753.graph_c0,c114770.graph_c0,c132637.graph_c0,c139000.graph_c0,c142609.graph_c0,c142660.graph_c0 |
| PIF | 5 | c133943.graph_c0,c141950.graph_c1,c149048.graph_c0,c150297.graph_c2,c127986.graph_c0 |
| TCP | 4 | c142930.graph_c2,c143047.graph_c0,c155677.graph_c0,c146807.graph_c0 |
| DIVARICATA | 2 | c115684.graph_c0,c149481.graph_c0 |
| ILI | 2 | c136920.graph_c0,c138271.graph_c2 |
| bZIP | 2 | c126053.graph_c0,c139664.graph_c0 |
| Trihelix | 2 | c142626.graph_c0,c152569.graph_c0 |
| GLABRA | 2 | c112384.graph_c0,c148751.graph_c0 |
| GLK1 | 2 | c125082.graph_c0,c154436.graph_c0 |
| JUNGBRUNNEN 1 | 2 | c110936.graph_c0,c137167.graph_c0 |
| RF2a | 2 | c140153.graph_c0,c143983.graph_c0 |
| PCF | 1 | c150295.graph_c0 |
| CAULIFLOWER A-like | 1 | c132998.graph_c0 |
| IBH | 1 | c133190.graph_c0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Zhou, Y.; Wu, Y.; Cai, J.; Zhang, Y.; Shen, J.; Wu, S.; Liu, W.; Cai, M.; Cheng, T.; et al. Balance of Carotenoid Synthesis and Degradation Contributes to the Color Difference between Chinese Narcissus and Its Yellow-Tepal Mutant. Horticulturae 2022, 8, 727. https://doi.org/10.3390/horticulturae8080727
Zhang Y, Zhou Y, Wu Y, Cai J, Zhang Y, Shen J, Wu S, Liu W, Cai M, Cheng T, et al. Balance of Carotenoid Synthesis and Degradation Contributes to the Color Difference between Chinese Narcissus and Its Yellow-Tepal Mutant. Horticulturae. 2022; 8(8):727. https://doi.org/10.3390/horticulturae8080727
Chicago/Turabian StyleZhang, Yiming, Yi Zhou, Yijing Wu, Junhuo Cai, Yiqiang Zhang, Jincai Shen, Shibin Wu, Wenjie Liu, Ming Cai, Tangren Cheng, and et al. 2022. "Balance of Carotenoid Synthesis and Degradation Contributes to the Color Difference between Chinese Narcissus and Its Yellow-Tepal Mutant" Horticulturae 8, no. 8: 727. https://doi.org/10.3390/horticulturae8080727