
Comparison of Waterlogging Tolerance of Three Peach Rootstock Seedlings Based on Physiological, Anatomical and Ultra-Structural Changes

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material Preparation and Growth Conditions
2.2. Experimental Design and Treatments
2.3. Plant Vegetative Growth Measurements
2.4. Observation of Stomatal Morphology
2.5. Photosynthetic Parameters Measurements
2.6. Determination of Chlorophyll Fluorescence Parameters
2.7. Determination of Antioxidant Response System
2.7.1. Preparation of Crude Enzyme Extract
2.7.2. Determination of Antioxidant Enzyme Activities, Malondialdehyde, and Proline Content
2.8. Determination of Root Vigor and Root Respiration Intensity
2.9. Observation of Anatomical and Ultra-Structure of Roots, Stems and Leaves
2.10. Statistical Analysis
3. Results
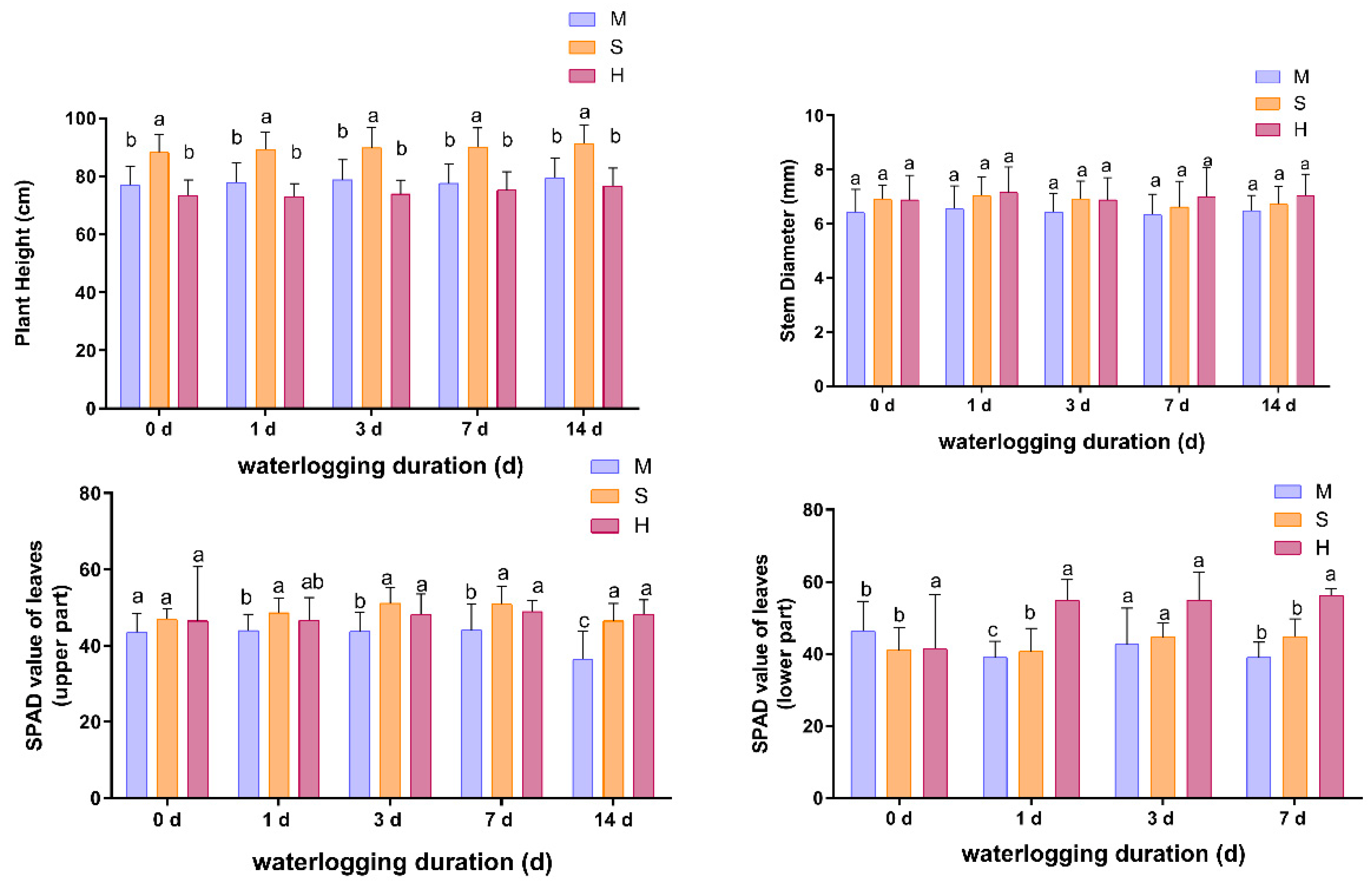
3.1. Effect of Waterlogging on Plant Height, Stem Diameter and Leaf SPAD Values
3.2. Effect of Waterlogging on Leaf Stomatal Density and Morphology
3.3. Effect of Waterlogging on Leaf Stomatal Morphological Parameters
3.4. Effect of Waterlogging on the Photosynthetic Capacity
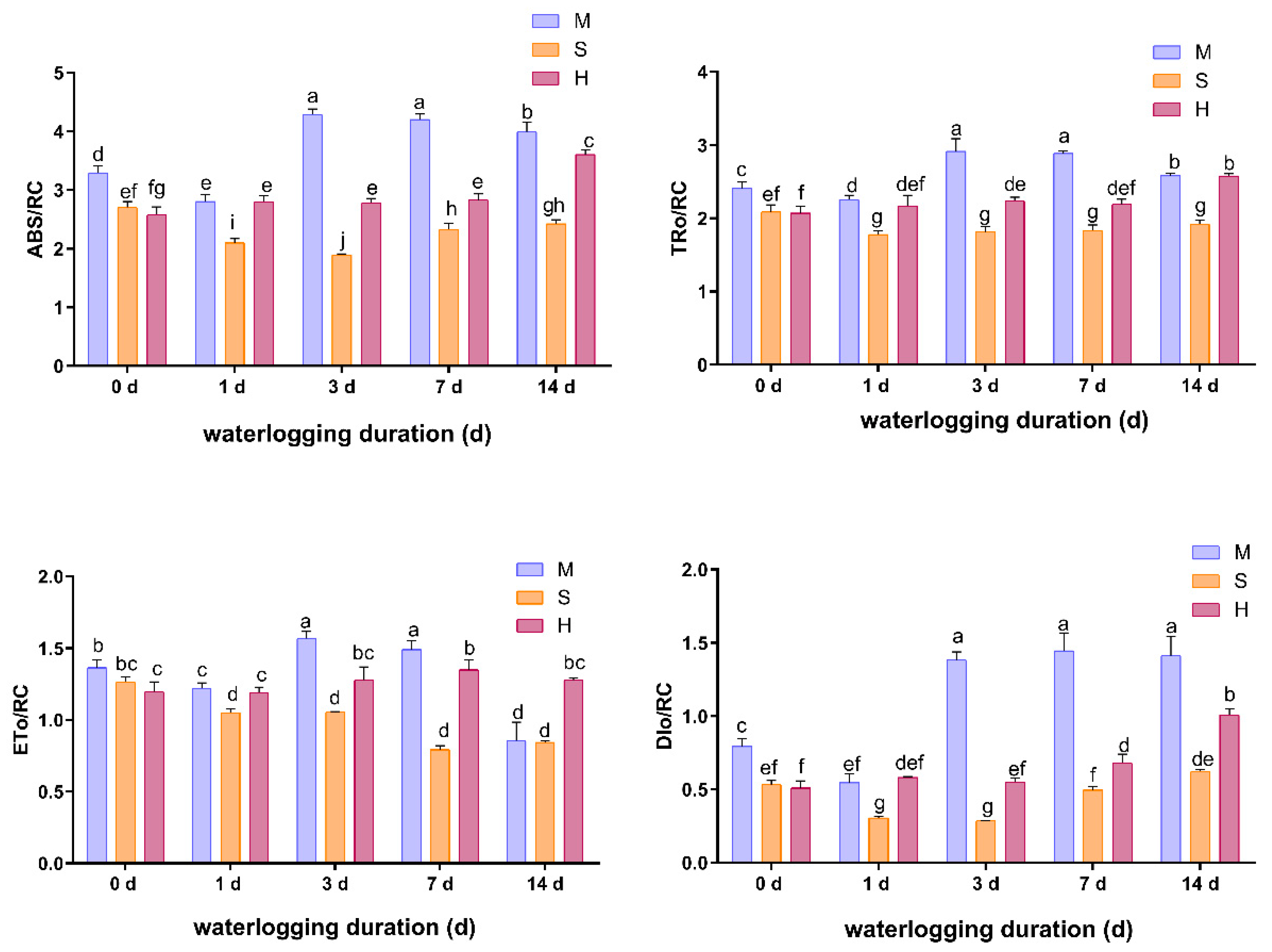
3.5. Effects of Waterlogging on Chlorophyll Indexes
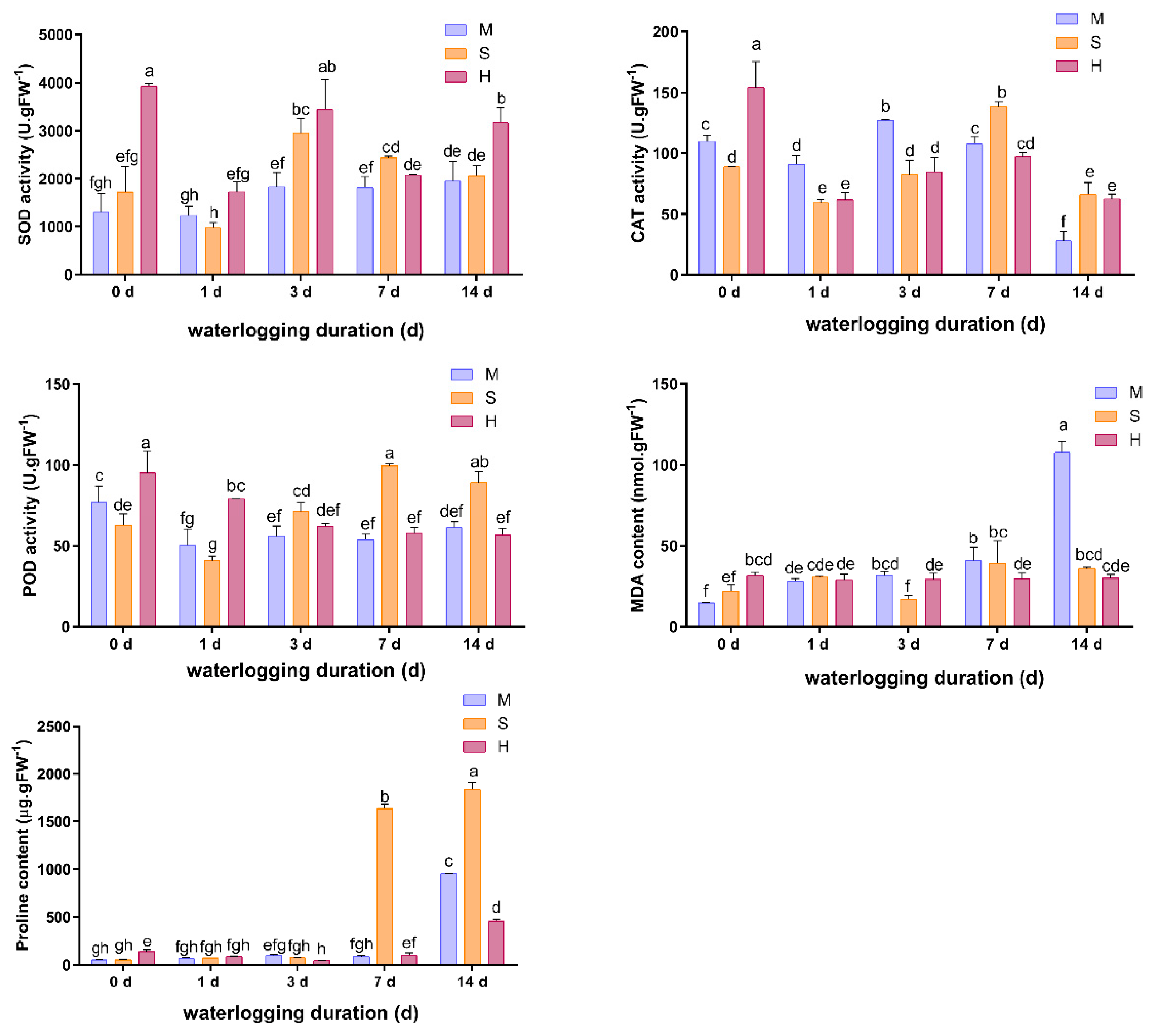
3.6. Effects of Waterlogging on Antioxidant and Osmoregulation Responses
3.7. Observation on the Anatomical Structure of Leaves and Fine Roots
3.7.1. Morphological Observations
3.7.2. Parameters of Leaf Anatomical Characteristics
3.7.3. Observation of the Anatomical Structure of Fine Roots
3.7.4. Parameters of Fine Roots Anatomical Characters
3.8. SEM Observation of Stems and Roots
3.9. TEM Observation of Leaves and Roots
3.10. Evaluation of Waterlogging Tolerance of Different Peach Rootstocks
4. Discussion
4.1. Morphological Responses
4.2. Photosynthetic Physiological Responses
4.3. Antioxidant Defense Responses
4.4. Osmoregulation Responses
4.5. Anatomical and Ultrastructural Responses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bhusal, N.; Kim, H.S.; Hanf, S.G.; Yoona, T.M. Photosynthetic traits and plant–water relations of two apple cultivars grown as bi-leader trees under long-term waterlogging conditions. Environ. Exp. Bot. 2020, 176, 104111. [Google Scholar] [CrossRef]
- Peng, Y.J.; Zhou, Z.X.; Tong, R.G.; Hu, X.Y.; Du, K.B. Anatomy and ultrastructure adaptation to soil flooding of two full-sib poplar clones differing in flood-tolerance. Flora-Morphol. Distrib. Funct. Ecol. Plants 2017, 233, 90–98. [Google Scholar] [CrossRef]
- Olorunwa, O.J.; Adhikari, B.; Shi, A.N.; Barickman, T.C. Screening of cowpea (Vigna unguiculata (L.) Walp.) genotypes for waterlogging tolerance using morpho-physiological traits at early growth stage. Plant Sci. 2022, 315, 111136. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.S.; Wu, X.; Sun, M.; Peng, F. Hydrogen sulfide alleviates waterlogging-induced damage in peach seedlings via enhancing antioxidative system and inhibiting ethylene synthesis. Front. Plant Sci. 2020, 11, 696. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, V.H.; Ploschuk, E.; Weibel, A.; Insausti, P. Short-term responses to flooding stress of three Prunus rootstocks. Sci. Hortic. 2017, 224, 135–141. [Google Scholar] [CrossRef]
- Gu, X.B.; Xue, L.; Lu, L.H.; Xiao, J.P.; Song, G.H.; Xie, M.; Zhang, H.Q. Melatonin enhances the waterlogging tolerance of Prunus persica by modulating antioxidant metabolism and anaerobic respiration. J. Plant Growth Regul. 2020, 4, 2178–2190. [Google Scholar] [CrossRef]
- Pantelidis, G.; Mavromatis, T.; Drogoudi, P. Consecutive wet days may impede fruit quality of peach and nectarine and cause fruit drop. Sci. Hortic. 2021, 282, 11011. [Google Scholar] [CrossRef]
- Nicotra, A.; Moser, L. Breeding program for peach rootstocks resistant to waterlogging. Acta Hortic. 1988, 224, 345–354. [Google Scholar] [CrossRef]
- Huang, C.; Gao, Y.; Qin, A.Z.; Liu, Z.G.; Zhao, B.; Ning, D.F.; Ma, S.T.; Duan, A.W.; Liu, Z.D. Effects of waterlogging at different stages and duration on maize growth and grain yields. Agric. Wat. Manag. 2022, 261, 107334. [Google Scholar] [CrossRef]
- Salah, A.; Nwafor, C.C.; Han, Y.L.; Liu, L.; Rashid, M.; Batool, M.; El-Badri, A.M.; Cao, C.G.; Zhan, M. Spermidine and brassinosteroid regulate root anatomical structure, photosynthetic traits and antioxidant defense systems to alleviate waterlogging stress in maize seedlings. S. Afr. J. Bot. 2022, 144, 389–402. [Google Scholar] [CrossRef]
- Fukao, T.; Barrera-Figueroa, B.E.; Juntawong, P.; Peña-Castro, J.M. Submergence and waterlogging stress in plants: A review highlighting research opportunities and understudied aspects. Front. Plant Sci. 2019, 10, 340. [Google Scholar] [CrossRef] [PubMed]
- Insausti, P.; Gorjón, S. Floods affect physiological and growth variables of peach trees (Prunus persica (L.) Batsch), as well as the postharvest behavior of fruits. Sci. Hortic. 2013, 152, 56–60. [Google Scholar] [CrossRef]
- Zhao, T.; Pan, X.J.; Qu, Z.G.; Li, Q.; Zhang, W.E. Comprehensive evaluation of waterlogging tolerance of eleven Canna cultivars at flowering stage. Sci. Hortic. 2022, 296, 110890. [Google Scholar] [CrossRef]
- Bailey-Serres, J.; Voesenek, L. Flooding stress: Acclimations and genetic diversity. Annu. Rev. Plant Biol. 2008, 59, 313–339. [Google Scholar] [CrossRef] [Green Version]
- Bertolde, F.Z.; De Almeida, A.-A.F.; Corrêa, R.X.; Gomes, F.P.; Gaiotto, F.A.; Baligar, V.C.; Loguercio, L.L. Molecular, physiological and morphological analysis of waterlogging tolerance in clonal genotypes of Theobroma cacao L. Tree Physiol. 2010, 30, 56–67. [Google Scholar] [CrossRef] [Green Version]
- Michelle, M.O.; Mauricio, O.; Gabriel, S. Effects of transient soil waterlogging and its importance for rootstock selection. Chil. J. Agric. Res. 2015, 75, 45–56. [Google Scholar]
- Pérez-Jiménez, M.; Pérez-Tornero, O. Short-term waterlogging in citrus rootstocks. Plants 2021, 10, 2772. [Google Scholar] [CrossRef]
- Pistelli, L.; Iacona, C.; Miano, D.; Cirilli, M.; Colao, M.C.; Mensuali-Sodi, A.; Muleo, R. Novel Prunus rootstock somaclonal variants with divergent ability to tolerate waterlogging. Tree Physiol. 2012, 32, 355–368. [Google Scholar] [CrossRef] [Green Version]
- Guajardo, V.; Hinrichsen, P.; Muñoz, C. Breeding rootstocks for Prunus species: Advances in genetic and genomics of peach and cherry as a model. Chil. J. Agric. Res. 2015, 75, 17–27. [Google Scholar] [CrossRef] [Green Version]
- Rubio-Cabetas, M.J.; Clara, P.; Beatriz, B.; Amador, M.L.; Cristina, M.; Antonio, G. Preformed and induced mechanisms underlies the differential responses of Prunus rootstock to hypoxia. J. Plant Physiol. 2018, 228, 134–149. [Google Scholar] [CrossRef]
- Purnobasuki, H.; Nurhidayati, T.; Hariyanto, S.; Jadid, N. Data of root anatomical responses to periodic waterlogging stress of tobacco (Nicotiana tabacum) varieties. Data Brief 2018, 20, 2012–2016. [Google Scholar] [CrossRef] [PubMed]
- Baghel, L.; Kataria, S.; Jain, M. Mitigation of adverse effects of salt stress on germination, growth, photosynthetic efficiency and yield in maize (Zea mays L.) through magnetopriming. Acta Agrobot. 2019, 72, 1757–1772. [Google Scholar] [CrossRef] [Green Version]
- Falqueto, A.R.; Silva, F.S.P.; Daniela, C.; Magalhães Júnior, A.M. Chlorophyll fluorescence in rice: Probing of senescence driven changes of PSII activity on rice varieties differing in grain yield capacity. Braz. J. Plant Physiol. 2010, 22, 35–41. [Google Scholar] [CrossRef]
- Marashi, S.K.; Mojaddam, M. Adventitious root and aerenchyma development in wheat (Triticum aestivum L.) subjected to waterlogging. Int. J. Biosci. 2014, 5, 168–173. [Google Scholar]
- Ren, B.; Zhang, J.; Dong, S.; Peng, L.; Zhao, B. Effects of waterlogging on leaf mesophyll cell ultra-structure and photosynthetic characteristics of summer maize. PLoS ONE 2016, 11, e0161424. [Google Scholar] [CrossRef] [Green Version]
- Wu, W.M.; Li, J.C.; Chen, H.J.; Wang, S.J.; Yi, Z. Effects of nitrogen fertilization on chlorophyll fluorescence change in maize (Zea mays L.) under waterlogging at seedlings stage. J. Food Agric. Environ. 2013, 11, 545–552. [Google Scholar]
- Zhang, F.; Zhu, K.; Wang, Y.Q.; Zhang, Z.P.; Zou, J.Q. Changes in photosynthetic and chlorophyll fluorescence characteristics of sorghum under drought and waterlogging stress. Photosynthetica 2019, 57, 1156–1164. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.J.; Liu, G.; Dong, H.; Li, C. Waterlogging stress in cotton: Damage, adaptability, alleviation strategies, and mechanisms. Crop. J. 2021, 9, 257–270. [Google Scholar] [CrossRef]
- Zhu, M.; Wang, J.; Li, F.H.; Shi, Z.S. Physiological and photosynthesis response of popcorn inbred seedlings to waterlogging stress. Pak. J. Bot. 2015, 47, 2069–2075. [Google Scholar]
- He, L.Z.; Yu, L.; Li, B.; Du, N.S.; Guo, S.R. The effect of exogenous calcium on cucumber fruit quality, photosynthesis, chlorophyll fluorescence, and fast chlorophyll fluorescence during the fruiting period under hypoxic stress. BMC Plant Biol. 2018, 18, 180–189. [Google Scholar] [CrossRef]
- Tuo, X.Q.; Li, S.; Wu, Q.S.; Zou, Y.N. Alleviation of waterlogged stress in peach seedlings inoculated with Funneliformis mosseae: Changes in chlorophyll and proline metabolism. Sci. Hortic. 2015, 197, 130–134. [Google Scholar] [CrossRef]
- Zheng, F.L.; Liang, S.M.; Chu, X.N.; Yang, Y.L.; Wu, Q.S. Mycorrhizal fungi enhance flooding tolerance of peach through inducing proline accumulation and improving root architecture. Plant Soil. Environ. 2020, 66, 624–631. [Google Scholar] [CrossRef]
- Wang, P.; Qiu, N.W.; Hou, W.Y.; Song, X.X.; Shu, F.Y.; Zhou, F. Experimental design reoptimization for “The effects of light and K+ on stomatal opening”. Plant Physiol. J. 2020, 56, 779–788. (In Chinese) [Google Scholar]
- Kolton, A.; Keska, K.; Czernicka, M. Selection of tomato and cucumber accessions for waterlogging sensitivity through morpho-physiological assessment at early vegetative stage. Agronomy 2020, 10, 1490. [Google Scholar] [CrossRef]
- Sánchez-Mata, D.; de la Fuente, V.; Rufo, L.; Rodríguez, N.; Amils, R. Localization of nickel in tissues of Streptanthus polygaloides Gray (Cruciferae) and endemic nickel hyperaccumulators from California. Biol. Trace Elem. Res. 2014, 157, 75–83. [Google Scholar] [CrossRef]
- El-Taher, A.M.; El-Raouf, H.S.A.; Osman, N.A.; Azoz, S.N.; Omar, M.A.; Elkelish, A.; El-Hady, M.A.M.A. Effect of salt stress and foliar application of salicylic acid on morphological, biochemical, anatomical, and productivity characteristics of Cowpea (Vigna unguiculata L.) plants. Plants 2022, 11, 115. [Google Scholar] [CrossRef]
- Jia, W.T.; Ma, M.H.; Chen, J.L.; Wu, S.J. Plant Morphological, physiological and anatomical adaption to flooding stress and the underlying molecular mechanisms. Int. J. Mol. Sci. 2021, 22, 1088. [Google Scholar] [CrossRef]
- Iacona, C.; Cirilli, M.; Zega, A.; Frionni, E.; Silvestri, C.; Muleo, R. A somaclonal myrobalan rootstock increases waterlogging tolerance to peach cultivar in controlled conditions. Sci. Hortic. 2013, 156, 1–8. [Google Scholar] [CrossRef]
- Gravatt, D.A.; Kirby, C.J. Patterns of photosynthesis and starch allocation in seedlings of four bottomland hardwood tree species subjected to flooding. Tree Physiol. 1998, 18, 411–417. [Google Scholar] [CrossRef]
- Toral-Juárez, M.A.; Avila, R.T.; Cardoso, A.A.; Brito, F.A.L.; Machado, K.L.G.; Almeida, W.L.; Souza, R.P.B.; Martins, S.C.V.; DaMatta, F.M. Drought-tolerant coffee plants display increased tolerance to waterlogging and post-waterlogging reoxygenation. Environ. Exp. Bot. 2021, 182, 104311. [Google Scholar] [CrossRef]
- Meloni, D.A.; Martínez, C.A. Physiological responses of Eucalyptus camaldulensis (Dehnh.) to simulated glyphosate drift. BIOFIX Sci. J. 2021, 6, 46–53. [Google Scholar] [CrossRef]
- Du, K.B.; Xu, L.; Wu, H.; Tu, B.K.; Zheng, B. Ecophysiological and morphological adaptation to soil flooding of two poplar clones differing in flood-tolerance. Flora-Morphol. Distrib. Funct. Ecol. Plants 2012, 207, 96–106. [Google Scholar] [CrossRef]
- Pimentel, P.; Almada, R.; Salvatierra, A.; Toro, G. Physiological and morphological responses of Prunus species with different degree of tolerance to long-term root hypoxia. Sci. Hortic. 2014, 180, 14–23. [Google Scholar] [CrossRef]
- McGee, T.; Shahid, M.A.; Beckman, T.G.; Chaparro, J.X.; Schaffer, B.; Sarkhosh, A. Physiological and biochemical characterization of six Prunus rootstocks in response to flooding. Environ. Exp. Bot. 2021, 183, 104368. [Google Scholar] [CrossRef]
- Ahmad, P.; Jaleel, C.A.; Salem, M.A.; Nabi, G.; Sharma, S. Roles of enzymatic and non-enzymatic antioxidants in plants during abiotic stress. Crit. Rev. Biotechnol. 2010, 30, 161–175. [Google Scholar] [CrossRef] [PubMed]
- Alves, J.D.; Zanandrea, I.; Deuner, S.; Goulart, P.F.P.; de Souza, R.D.; Santos, O.M. Antioxidative responses and morpho-anatomical adaptations to waterlogging in Sesbania virgata. Trees 2013, 27, 717–728. [Google Scholar] [CrossRef]
- Miller, G.; Shulaev, V.; Mittler, R. Reactive oxygen signaling and abiotic stress. Physiol. Plant. 2010, 133, 481–489. [Google Scholar] [CrossRef]
- Cheng, Z.Y.; Sun, L.; Wang, X.J.; Sun, R.; An, Y.Q.; An, B.L.; Zhu, M.X.; Zhao, C.F.; Bai, J.G. Ferulic acid pretreatment alleviates heat stress in blueberry seedlings by inducing antioxidant enzymes, proline, and soluble sugars. Biol. Plant. 2018, 62, 534–542. [Google Scholar] [CrossRef]
- Chen, J.; Shang, Y.T.; Wang, W.H.; Chen, X.Y.; He, E.M.; Zheng, H.L.; Shangguan, Z.P. Hydrogen sulfide-induced polyamines and sugar changes are involved in hydrogen sulfide-induced drought tolerance in spinacia oleracea seedlings. Front. Plant Sci. 2016, 7, 1173. [Google Scholar] [CrossRef] [Green Version]
- Evans, D.E. Aerenchyma formation. New Phytol. 2003, 161, 35–49. [Google Scholar] [CrossRef]
- Sarkar, P.; Gladish, D.K. Hypoxic stress triggers a programmed cell death pathway to induce vascular cavity formation in Pisum sativum roots. Physiol. Plant 2012, 146, 413–426. [Google Scholar] [CrossRef]
- Yin, D.M.; Zhang, Z.G.; Luo, H.L. Anatomical responses to waterlogging in Chrysanthemum zawadskii. Sci. Hortic. 2012, 146, 86–91. [Google Scholar] [CrossRef]
- Omi, S.K.; Robin, R. Notes: Amyloglucosidase and α-amylase enzymes for determining starch concentration in ponderosa pine root tissue. For. Sci. 1991, 5, 1479–1483. [Google Scholar]
- Rutto, K.L.; Mizutani, F.; Kadoya, K. Effect of root-zone flooding on mycorrhizal and non-mycorrhizal peach (Prunus persica Batsch) seedlings. Sci. Hortic. 2002, 94, 285–295. [Google Scholar] [CrossRef]
- Rutto, K.L.; Mizutani, F.; Moon, D.G.; Cho, Y.S.; Kadoya, K. Seasonal fluctuations in mycorrhizal spore populations and infection rates of vineyard soils planted with five legume cover crops. J. Jpn. Soc. Hortic. Sci. 2003, 72, 262–267. [Google Scholar] [CrossRef] [Green Version]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stomatal Density (Numbers/mm2) | Stomatal Closure Ratio (%) | |||||
|---|---|---|---|---|---|---|
| Rootstocks | WT-0 d | WT-3 d | WT-14 d | WT-0 d | WT-3 d | WT-14 d |
| M | 286.61 ± 24.61 ab | 301.25 ± 7.62 a | 216.33 ± 5.69 c | 7.70% | 21.90% | 67.20% |
| S | 205.62 ± 23.65 cd | 215.33 ± 12.03 c | 203.58 ± 5.68 cd | 14.50% | 36.00% | 52.20% |
| H | 275.74 ± 13.21 b | 205.62 ± 17.56 cd | 306.35 ± 8.19 a | 10.20% | 36.20% | 56.30% |
| Cultivar | Waterlogged Duration (d) | Stomatal Width (μm) | Stomatal Length (μm) |
|---|---|---|---|
| M | WT-0 d | 6.41 ± 1.42 a | 16.59 ± 2.20 b |
| WT-3 d | 6.54 ± 0.56 a | 15.46 ± 1.49 b | |
| WT-14 d | 5.74 ± 0.93 b | 21.90 ± 1.80 a | |
| S | WT-0 d | 8.03 ± 0.75 a | 23.96 ± 1.52 a |
| WT-3 d | 5.95 ± 1.13 b | 19.79 ± 1.69 b | |
| WT-14 d | 3.89 ± 0.52 c | 19.00 ± 1.39 b | |
| H | WT-0 d | 6.45 ± 1.20 a | 19.26 ± 1.97 ab |
| WT-3 d | 5.78 ± 0.73 b | 19.81 ± 0.91 a | |
| WT-14 d | 3.26 ± 0.69 c | 12.79 ± 1.10 c |
| Cultivar | Waterlogged Duration (d) | Upper Epidermis Thickness (μm) | Lower Epidermis Thickness (μm) | Leaf Thickness (μm) | Palisade Cell Length/Width Ratio | CTR% | SR% | P/S Ratio |
|---|---|---|---|---|---|---|---|---|
| M | WT-0 d | 20.39 ± 3.53 b | 10.01 ± 0.84 a | 134.02 ± 2.20 c | 7.23 | 0.34 | 0.37 | 0.92 |
| WT-3 d | 21.47 ± 3.76 b | 10.32 ± 0.90 a | 147.76 ± 2.03 b | 6.60 | 0.33 | 0.43 | 0.75 | |
| WT-14 d | 27.35 ± 3.20 a | 10.97 ± 2.88 a | 177.18 ± 2.72 a | 12.96 | 0.40 | 0.39 | 1.01 | |
| S | WT-0 d | 34.03 ± 6.92 a | 12.93 ± 1.30 ab | 185.60 ± 3.05 a | 12.42 | 0.44 | 0.35 | 1.24 |
| WT-3 d | 29.88 ± 5.65 ab | 16.33 ± 3.15 a | 172.60 ± 2.69 b | 17.10 | 0.41 | 0.37 | 1.12 | |
| WT-14 d | 23.87 ± 2.32 c | 13.15 ± 3.29 ab | 118.38 ± 2.47 c | 9.80 | 0.34 | 0.37 | 0.93 | |
| H | WT-0 d | 18.86 ± 2.01 b | 12.72 ± 1.34 a | 170.91 ± 2.69 b | 16.77 | 0.39 | 0.36 | 1.09 |
| WT-3 d | 21.86 ± 4.33 a | 12.66 ± 2.25 a | 182.92 ± 0.49 a | 10.39 | 0.31 | 0.36 | 0.87 | |
| WT-14 d | 18.29 ± 1.65 b | 9.46 ± 0.44 b | 141.95 ± 3.51 c | 13.38 | 0.38 | 0.41 | 0.93 |
| Cultivar | Waterlogged Duration (d) | Thickness of Peripheral Epidermis (μm) | Thickness of Phloem (μm) | The Diameter of Root Column (μm) |
|---|---|---|---|---|
| M | WT-0 d | - | 101.41 ± 1.95 a | 55.35 ± 10.62 c |
| WT-3 d | - | 104.74 ± 15.38 a | 104.67 ± 15.25 a | |
| WT-14 d | - | 105.61 ± 3.32 a | 74.77 ± 1.53 c | |
| S | WT-0 d | 31.57 ± 8.58 bc | 69.86 ± 5.88 c | 156.52 ± 15.68 b |
| WT-3 d | 39.15 ± 2.98 b | 91.33 ± 3.68 b | 175.87 ± 7.60 b | |
| WT-14 d | 56.96 ± 4.05 a | 112.38 ± 30.48 a | 278.54 ± 5.24 a | |
| H | WT-0 d | - | 177.82 ± 14.18 b | 124.52 ± 6.61 b |
| WT-3 d | - | 82.70 ± 4.43 c | 205.14 ± 28.05 a | |
| WT-14 d | - | 225.09 ± 15.21 a | 127.49 ± 15.17 b |
| Principal Component | Parameters | Initial Eigenvalue | Total Variance of Interpretation | |
|---|---|---|---|---|
| Contribution Rate % | Cumulative Contribution Rate % | |||
| 1 | PH (Plant Height) | 8.03 | 29.74 | 29.74 |
| 2 | SD (Stem Diameter) | 7.178 | 26.587 | 56.327 |
| 3 | SPAD upper | 2.703 | 10.01 | 66.337 |
| 4 | SPAD lower | 2.074 | 7.683 | 74.021 |
| 5 | Ci | 1.315 | 4.871 | 78.892 |
| 6 | Pn | 1.182 | 4.379 | 83.272 |
| 7 | gs | 1.08 | 4 | 87.271 |
| 8 | E | 0.983 | 3.639 | 90.911 |
| 9 | WUE | 0.549 | 2.032 | 92.942 |
| 10 | Ls | 0.424 | 1.57 | 94.513 |
| 11 | Fo | 0.317 | 1.175 | 95.688 |
| 12 | Fm | 0.239 | 0.886 | 96.574 |
| 13 | Fv/Fm | 0.229 | 0.848 | 97.421 |
| 14 | Fv/Fo | 0.186 | 0.69 | 98.111 |
| 15 | ABS/RC | 0.126 | 0.467 | 98.578 |
| 16 | TRo/RC | 0.112 | 0.415 | 98.993 |
| 17 | ETo/RC | 0.084 | 0.31 | 99.302 |
| 18 | DIo/RC | 0.047 | 0.175 | 99.477 |
| 19 | PIabs | 0.041 | 0.152 | 99.629 |
| 20 | PItotal | 0.031 | 0.114 | 99.743 |
| 21 | SOD | 0.029 | 0.106 | 99.849 |
| 22 | CAT | 0.015 | 0.054 | 99.904 |
| 23 | POD | 0.011 | 0.041 | 99.945 |
| 24 | MDA | 0.006 | 0.022 | 99.968 |
| 25 | Proline | 0.005 | 0.018 | 99.985 |
| 26 | TTC | 0.004 | 0.014 | 100 |
| 27 | Root respiration rate | 7.62 × 10−5 | 0 | 100 |
| Parameters | M-0 d | M-1 d | M-3 d | M-7 d | M-14 d | S-0 d | S-1 d | S-3 d | S-7 d | S-14 d | H-0 d | H-1 d | H-3 d | H-7 d | H-14 d |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Plant Height | 0.21 | 0.26 | 0.31 | 0.25 | 0.35 | 0.83 | 0.89 | 0.92 | 0.93 | 1.00 | 0.02 | 0.00 | 0.05 | 0.12 | 0.20 |
| Stem Diameter | 0.10 | 0.26 | 0.11 | 0.00 | 0.17 | 0.69 | 0.85 | 0.70 | 0.34 | 0.47 | 0.65 | 1.00 | 0.66 | 0.81 | 0.86 |
| SPAD upper | 0.45 | 0.54 | 0.50 | 0.50 | 0.00 | 0.70 | 0.82 | 1.00 | 0.95 | 0.68 | 0.68 | 0.69 | 0.79 | 0.86 | 0.79 |
| SPAD lower | 0.53 | 0.12 | 0.34 | 0.13 | 0.00 | 0.21 | 0.21 | 0.40 | 0.42 | 0.50 | 0.26 | 0.92 | 0.94 | 1.00 | 0.59 |
| Ci | 0.35 | 0.10 | 0.00 | 0.32 | 0.31 | 0.12 | 0.73 | 0.32 | 1.00 | 0.49 | 0.38 | 0.26 | 0.31 | 0.24 | 0.67 |
| Pn | 0.75 | 0.53 | 0.45 | 0.15 | 0.09 | 0.68 | 0.18 | 0.28 | 0.00 | 0.17 | 1.00 | 0.67 | 0.74 | 0.16 | 0.12 |
| gs | 0.67 | 0.29 | 0.17 | 0.00 | 0.03 | 0.61 | 0.03 | 0.05 | 0.03 | 0.05 | 1.00 | 0.39 | 0.30 | 0.01 | 0.00 |
| E | 0.88 | 0.52 | 0.40 | 0.01 | 0.04 | 0.21 | 0.09 | 0.18 | 0.05 | 0.10 | 1.00 | 0.58 | 0.88 | 0.04 | 0.00 |
| WUE | 0.82 | 0.94 | 0.94 | 0.71 | 0.46 | 1.00 | 0.61 | 0.57 | 0.00 | 0.70 | 0.92 | 0.87 | 0.85 | 0.70 | 0.72 |
| Ls | 0.65 | 0.90 | 1.00 | 0.68 | 0.69 | 0.88 | 0.27 | 0.68 | 0.00 | 0.51 | 0.62 | 0.74 | 0.69 | 0.75 | 0.33 |
| Fo | 0.62 | 0.32 | 0.74 | 0.77 | 1.00 | 0.24 | 0.01 | 0.00 | 0.17 | 0.17 | 0.21 | 0.20 | 0.28 | 0.29 | 0.65 |
| Fm | 1.00 | 0.72 | 0.79 | 0.49 | 0.60 | 0.77 | 0.69 | 0.60 | 0.30 | 0.00 | 0.68 | 0.79 | 0.63 | 0.67 | 0.61 |
| Fv/Fm | 0.57 | 0.80 | 0.23 | 0.17 | 0.00 | 0.79 | 1.00 | 0.98 | 0.69 | 0.51 | 0.74 | 0.86 | 0.74 | 0.68 | 0.35 |
| Fv/Fo | 0.36 | 0.60 | 0.15 | 0.04 | 0.00 | 0.63 | 1.00 | 0.85 | 0.48 | 0.29 | 0.58 | 0.83 | 0.55 | 0.55 | 0.19 |
| ABS/RC | 0.58 | 0.38 | 1.00 | 0.96 | 0.88 | 0.34 | 0.09 | 0.00 | 0.18 | 0.22 | 0.29 | 0.38 | 0.37 | 0.39 | 0.71 |
| TRo/RC | 0.55 | 0.42 | 1.00 | 0.97 | 0.71 | 0.27 | 0.00 | 0.03 | 0.05 | 0.13 | 0.26 | 0.35 | 0.40 | 0.36 | 0.70 |
| ETo/RC | 0.74 | 0.55 | 1.00 | 0.90 | 0.08 | 0.61 | 0.33 | 0.34 | 0.00 | 0.07 | 0.52 | 0.52 | 0.63 | 0.72 | 0.63 |
| DIo/RC | 0.44 | 0.22 | 0.95 | 1.00 | 0.97 | 0.21 | 0.02 | 0.00 | 0.18 | 0.29 | 0.19 | 0.25 | 0.23 | 0.34 | 0.62 |
| PIabs | 0.18 | 0.29 | 0.12 | 0.05 | 0.00 | 0.38 | 0.79 | 1.00 | 0.19 | 0.15 | 0.35 | 0.32 | 0.37 | 0.32 | 0.10 |
| PItotal | 0.41 | 0.29 | 0.43 | 0.13 | 0.00 | 0.18 | 0.53 | 0.61 | 0.04 | 0.04 | 1.00 | 0.63 | 0.56 | 0.34 | 0.16 |
| SOD | 0.11 | 0.09 | 0.29 | 0.28 | 0.33 | 0.25 | 0.00 | 0.67 | 0.49 | 0.37 | 1.00 | 0.25 | 0.83 | 0.38 | 0.74 |
| CAT | 0.65 | 0.50 | 0.79 | 0.63 | 0.00 | 0.49 | 0.25 | 0.43 | 0.87 | 0.30 | 1.00 | 0.27 | 0.45 | 0.55 | 0.27 |
| POD | 0.62 | 0.16 | 0.26 | 0.22 | 0.35 | 0.38 | 0.00 | 0.52 | 1.00 | 0.83 | 0.93 | 0.65 | 0.36 | 0.29 | 0.27 |
| MDA | 0.00 | 0.14 | 0.18 | 0.28 | 1.00 | 0.08 | 0.17 | 0.02 | 0.26 | 0.23 | 0.18 | 0.15 | 0.16 | 0.16 | 0.16 |
| Proline | 0.00 | 0.01 | 0.03 | 0.02 | 0.51 | 0.00 | 0.01 | 0.02 | 0.89 | 1.00 | 0.05 | 0.02 | 0.00 | 0.03 | 0.23 |
| TTC | 0.10 | 0.21 | 1.00 | 0.56 | 0.06 | 0.07 | 0.14 | 0.55 | 0.14 | 0.00 | 0.29 | 0.60 | 0.86 | 0.32 | 0.27 |
| Root respiration | 0.24 | 0.27 | 1.00 | 0.45 | 0.19 | 0.46 | 0.00 | 0.55 | 0.10 | 0.02 | 0.52 | 0.09 | 0.62 | 0.39 | 0.21 |
| Average value | 0.47 | 0.39 | 0.52 | 0.40 | 0.33 | 0.45 | 0.36 | 0.45 | 0.36 | 0.34 | 0.57 | 0.49 | 0.53 | 0.43 | 0.41 |
| Ranking * | 5 | 10 | 3 | 9 | 13 | 6 | 11 | 6 | 11 | 12 | 1 | 4 | 2 | 7 | 8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, F.; Cai, H.; Zhang, X.; Su, M.; Zhou, H.; Li, X.; Zhang, M.; Hu, Y.; Gu, C.; Du, J.; et al. Comparison of Waterlogging Tolerance of Three Peach Rootstock Seedlings Based on Physiological, Anatomical and Ultra-Structural Changes. Horticulturae 2022, 8, 720. https://doi.org/10.3390/horticulturae8080720
Xu F, Cai H, Zhang X, Su M, Zhou H, Li X, Zhang M, Hu Y, Gu C, Du J, et al. Comparison of Waterlogging Tolerance of Three Peach Rootstock Seedlings Based on Physiological, Anatomical and Ultra-Structural Changes. Horticulturae. 2022; 8(8):720. https://doi.org/10.3390/horticulturae8080720
Chicago/Turabian StyleXu, Fangjie, Huaqing Cai, Xianan Zhang, Mingshen Su, Huijuan Zhou, Xiongwei Li, Minghao Zhang, Yang Hu, Chao Gu, Jihong Du, and et al. 2022. "Comparison of Waterlogging Tolerance of Three Peach Rootstock Seedlings Based on Physiological, Anatomical and Ultra-Structural Changes" Horticulturae 8, no. 8: 720. https://doi.org/10.3390/horticulturae8080720