
Optimizing Phytochemical and Physiological Characteristics of Balangu (Lallemantia iberica) by Foliar Application of Chitosan Nanoparticles and Myco-Root Inoculation under Water Supply Restrictions

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Experimental Setup
2.2. Photosynthetic Pigments
2.3. Essential Oil Isolation
2.4. Identification of Essential Oil Compounds
2.5. Malondialdehyde (MDA)
2.6. Superoxide Dismutase (SOD) Activity
2.7. Ascorbate Peroxidase (APX) Activity
2.8. Peroxidase (POX) Activity
2.9. Proline
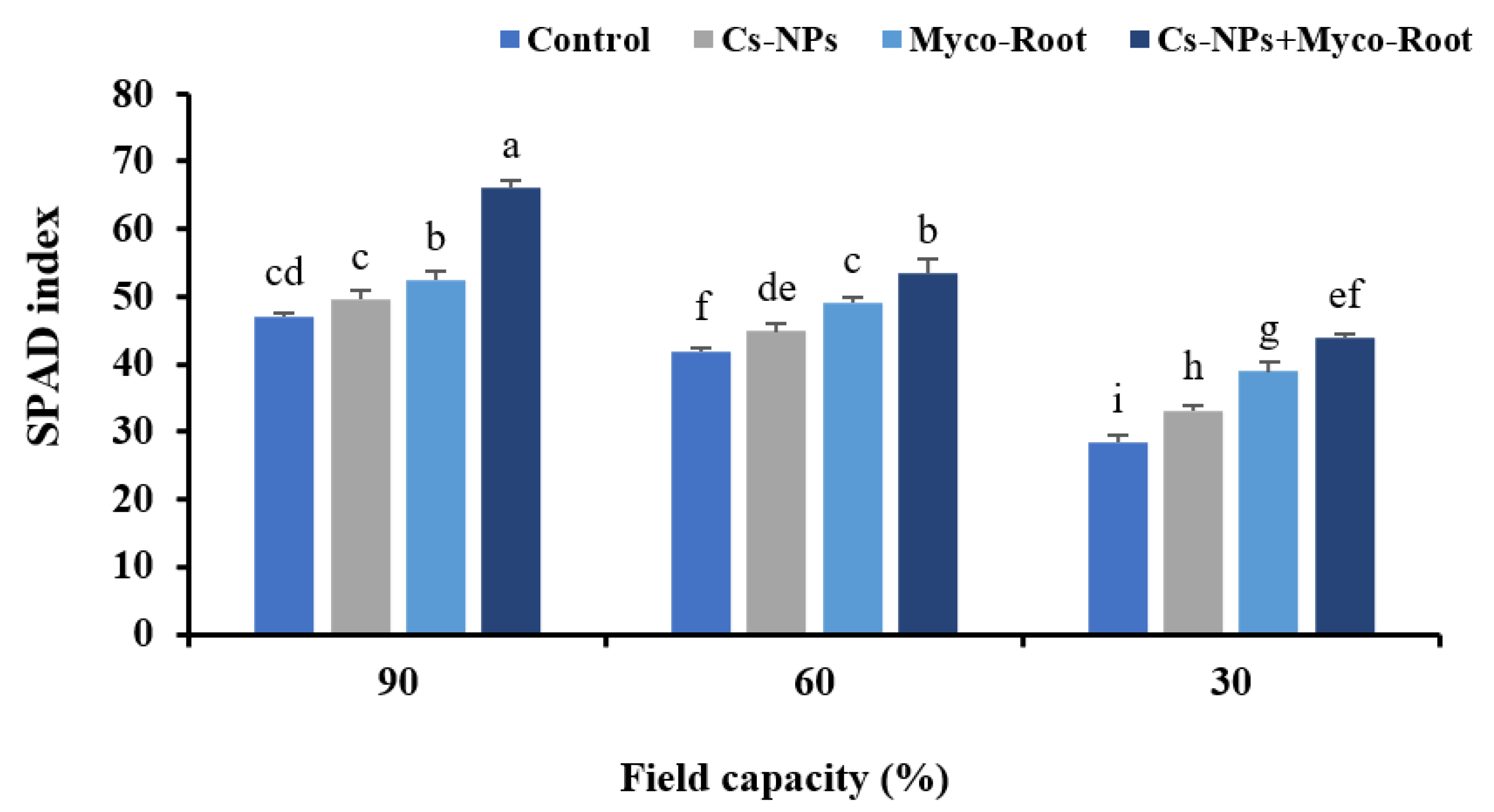
2.10. SPAD Index
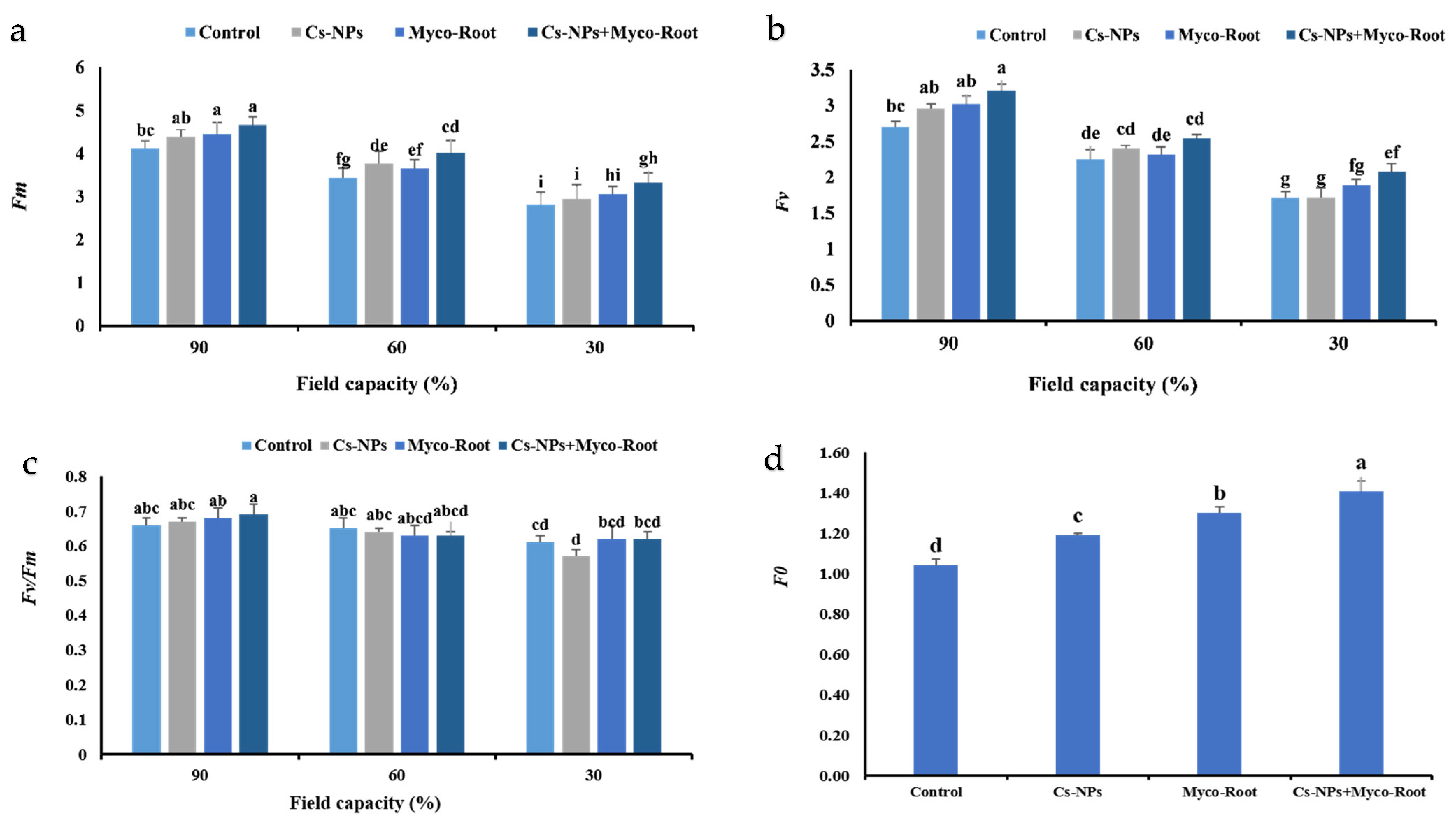
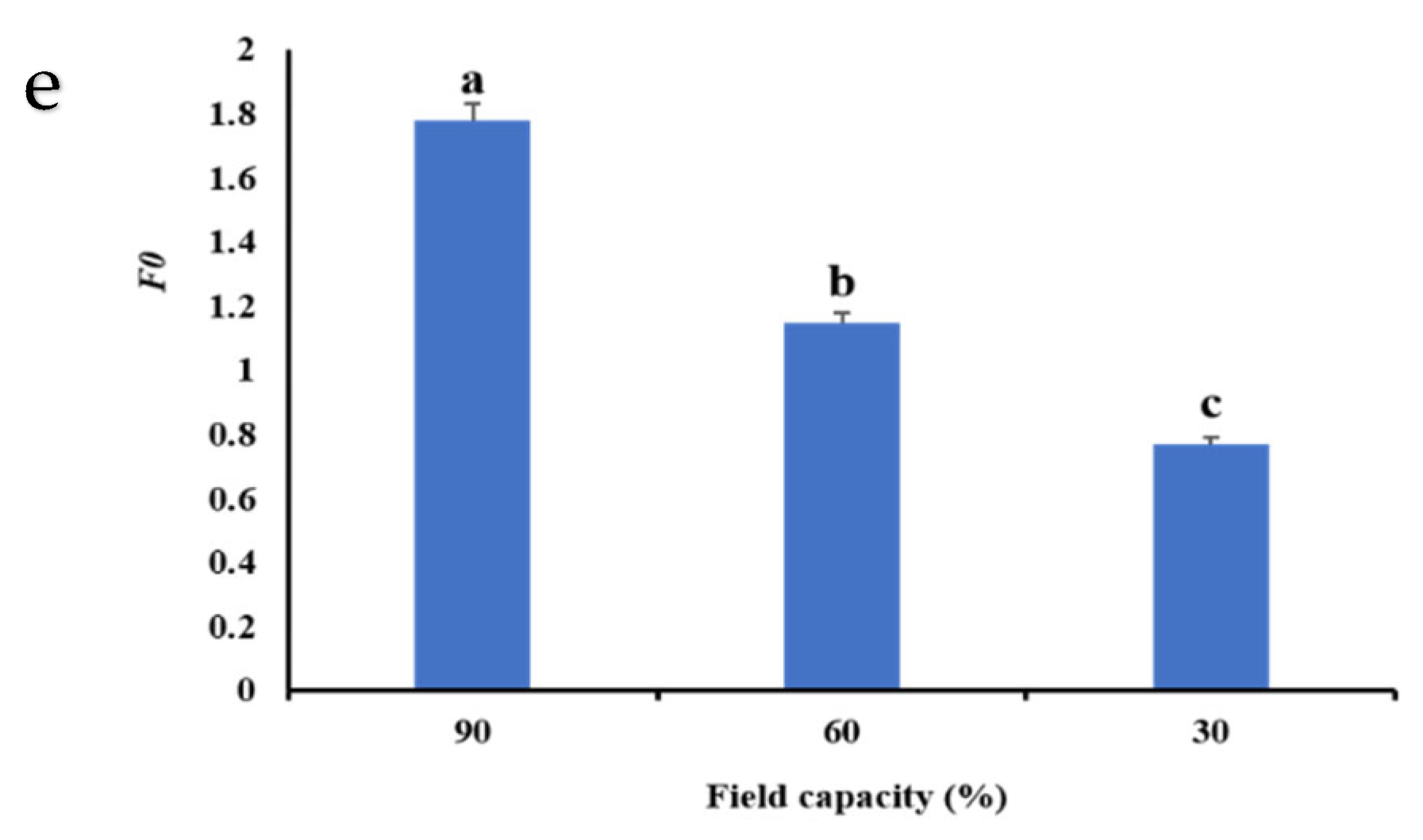
2.11. Chlorophyll Fluorescence Parameters
2.12. Statistical Analysis
3. Results
3.1. Fresh and Dry Weight (FW and DW)
3.2. Chlorophyll a, b and Carotenoid
3.3. Essential Oil Content (EO) and Essential Oil Yield (EOY)
3.4. EO Compositions
3.5. Chlorophyll Fluorescence Indices
3.6. Proline
3.7. Malondialdehyde (MDA)
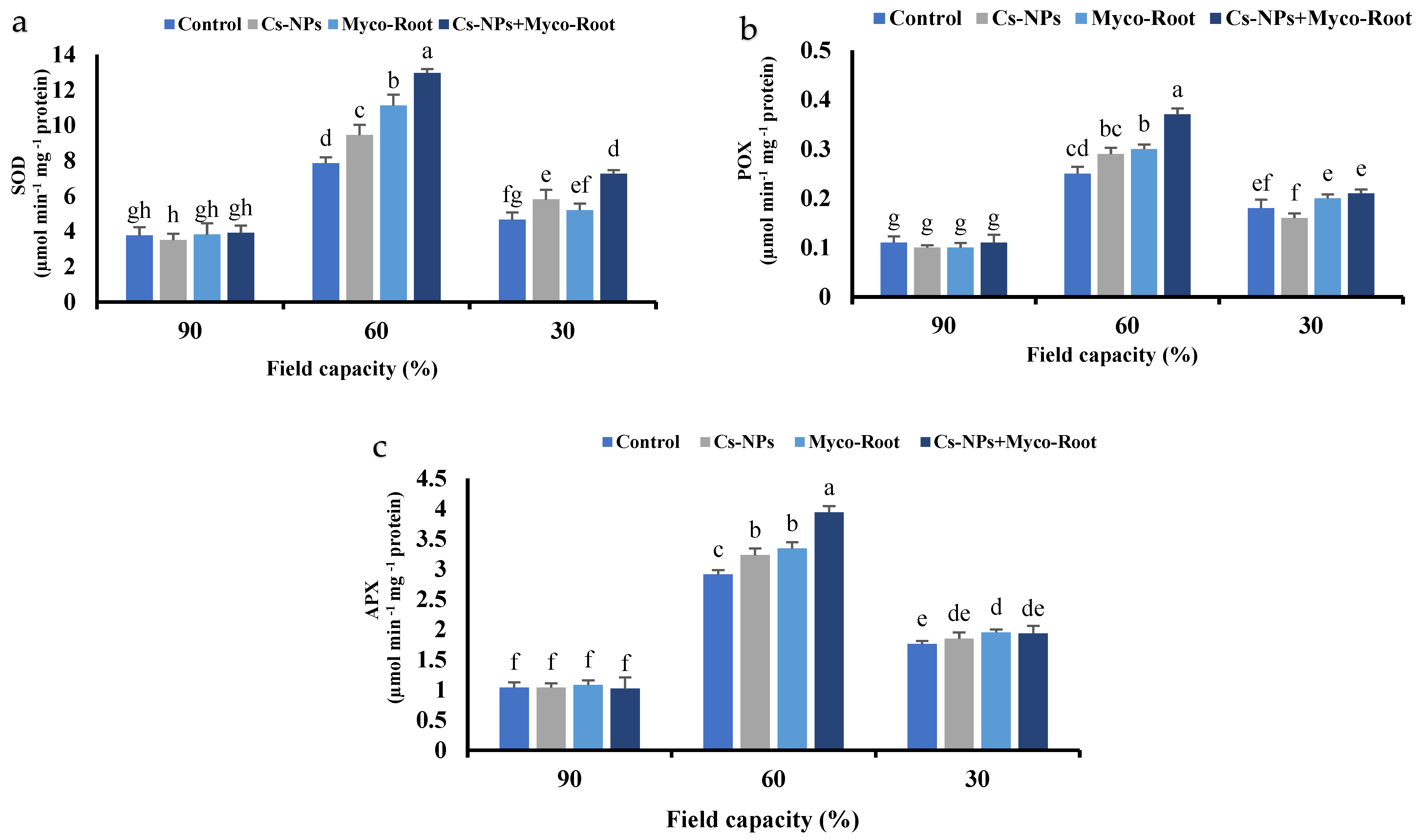
3.8. Superoxide Dismutase (SOD), Ascorbate Peroxidase (APX) and Peroxidase (POX) Activity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ursu, B.; Borcean, I. Researches Concerning the Sowing Technology at Lallemantia iberica F.et M. Res. J. Agric. Sci. 2012, 44, 168–171. [Google Scholar]
- Paravar, A.; Maleki Farahani, S.; Rezazadeh, A.R. Lallemantia species response to drought stress and Arbuscular mycorrhizal fungi application. Ind. Crops Prod. 2021, 172, 114002. [Google Scholar] [CrossRef]
- Ghasemi, V.M.; Moghaddam, S.S.; Rahimi, A.; Pourakbar, L.; Popović-Djordjević, J. Winter Cultivation and Nano Fertilizers Improve Yield Components and Antioxidant Traits of Dragon’s Head (Lallemantia iberica (m.b.) Fischer & Meyer). Plants 2020, 9, 252. [Google Scholar] [CrossRef] [Green Version]
- Omidi, H.; Shams, H.; Seif Sahandi, M.; Rajabian, T. Balangu (Lallemantia Sp.) Growth and Physiology under Field Drought Conditions Affecting Plant Medicinal Content. Plant Physiol. Biochem. 2018, 130, 641–646. [Google Scholar] [CrossRef] [PubMed]
- Nasrollahzadeh, S.; Aghaei-Gharachorlou, P. Effect of irrigation frequency and planting density on herbage biomass, essential oil production and mucilage yield of dragon’s head (Lallemantia iberica fish. Et mey.). IJAB 2014, 3, 89–94. [Google Scholar]
- Razavi, S.M.A.; Mohammadi, M.T. Influence of different substitution levels of Balangu seed gum on textural characteristics of selected hydrocolloids. Electron. J. Environ. Agric. Food Chem. 2011, 10, 2826–2837. [Google Scholar]
- Yuce, E.; Bagci, E. Study of the essential oil composition of Lallemantia iberica (M. Bieb.) Fisch. and C.A. Mey. (Lamiaceae) from Turkey. Asian J. Chem. 2012, 24, 4817–4818. [Google Scholar]
- Sachdev, S.; Ansari, S.A.; Ansari, M.I.; Fujita, M.; Hasanuzzaman, M. Abiotic stress and reactive oxygen species: Generation, signaling, and defense mechanisms. Antioxidants 2021, 10, 277. [Google Scholar] [CrossRef]
- Xu, J.; Jin, J.; Zhao, H. Drought stress tolerance analysis of Populus ussuriensis clones with different ploidies. J. Res. 2019, 30, 1267–1275. [Google Scholar] [CrossRef]
- Flexas, J.; Medrano, H.J. Drought-inhibition of photosynthesis in C3 plants: Stomatal and non-stomatal limitations revisited. Ann. Bot. 2002, 89, 183–189. [Google Scholar] [CrossRef] [Green Version]
- Karimi, S.; Abbaspour, H.; Sinaki, J.M.; Makarian, H. Effects of water deficit and chitosan spraying on osmotic adjustment and soluble protein of cultivars castor bean (Ricinus communis L.). J. Stress Physiol. Biochem. 2012, 28, 160–169. [Google Scholar]
- Dacosta, M.; Huang, B. Changes in antioxidant enzyme activities and lipid peroxidation for bent grass species in response to drought stress. J. Am. Soc. Hortic. Sci. 2007, 132, 319–326. [Google Scholar] [CrossRef] [Green Version]
- Ostadi, A.; Javanmard, A.; Amani Machiani, M.; Sadeghpour, A.; Maggi, F.; Nouraein, M.; Morshedloo, M.R.; Hano, C.; Lorenzo, J.M. Co-Application of TiO2 Nanoparticles and Arbuscular Mycorrhizal Fungi Improves Essential Oil Quantity and Quality of Sage (Salvia officinalis L.) in Drought Stress Conditions. Plants 2022, 11, 1659. [Google Scholar] [CrossRef]
- Amani Machiani, M.; Javanmard, A.; Morshedloo, M.R.; Aghaee, A.; Maggi, F. Funneliformis mosseae inoculation under water deficit stress improves the yield and phytochemical characteristics of thyme in intercropping with soybean. Sci. Rep. 2021, 11, 15279. [Google Scholar] [CrossRef]
- Strzemski, M.; Dzida, K.; Dresler, S.; Sowa, I.; Kurzepa, J.; Szymczak, G.; Wójciak, M. Nitrogen fertilisation decreases the yield of bioactive compounds in Carlina acaulis L. grown in the field. Ind. Crops Prod. 2021, 170, 113698. [Google Scholar] [CrossRef]
- Rasouli, F.; Amini, T.; Asadi, M.; Hassanpouraghdam, M.B.; Aazami, M.A.; Ercisli, S.; Skrovankova, S.; Mlcek, J. Growth and Antioxidant Responses of Lettuce (Lactuca sativa L.) to Arbuscular Mycorrhiza Inoculation and Seaweed Extract Foliar Application. Agronomy 2022, 12, 401. [Google Scholar] [CrossRef]
- Palacio-Márquez, A.; Ramírez-Estrada, C.A.; Gutiérrez-Ruelas, N.J.; Sánchez, E.; Ojeda-Barrios, D.L.; Chávez-Mendoza, C.; Sida-Arreola, J.P. Efficiency of foliar application of zinc oxide nanoparticles versus zinc nitrate complexed with chitosan on nitrogen assimilation, photosynthetic activity, and production of green beans (Phaseolus vulgaris L.). Sci. Hortic. 2021, 288, 110297. [Google Scholar] [CrossRef]
- Golkar, P.; Taghizadeh, M.; Yousefian, Z. The effects of chitosan and salicylic acid on elicitation of secondary metabolites and antioxidant activity of safflower under in vitro salinity stress. Plant Cell Tissue Organ Cult. 2019, 137, 575–585. [Google Scholar] [CrossRef]
- Farouk, S.; Amany, R. Improving growth and yield of cowpea by foliar application of chitosan under water stress. Egypt. J. Biol. Pest. Control 2012, 14, 15–27. [Google Scholar] [CrossRef] [Green Version]
- Hassan, F.A.S.; Ali, E.; Gaber, A.; Fetouh, M.I.; Mazrou, R. Chitosan Nanoparticles Effectively Combat Salinity Stress by Enhancing Antioxidant Activity and Alkaloid Biosynthesis in Catharanthus roseus (L.) G. Don. Plant Physiol. Biochem. 2021, 162, 291–300. [Google Scholar] [CrossRef]
- Tourian, N.; Sinaki, J.M.; Hasani, N.; Madani, H. Change in photosynthetic pigment concentration of wheat grass (Agropyron repens) cultivars response to drought stress and foliar application with chitosan. Int. J. Agron. Plant Prod. 2013, 4, 1084–1091. [Google Scholar]
- El-Serafy, R.S. Phenotypic Plasticity, Biomass allocation, and biochemical analysis of cordyline seedlings in response to oligo-chitosan foliar spray. Soil Sci. Plant Nutr. 2020, 20, 1503–1514. [Google Scholar] [CrossRef]
- Emami Bistgani, Z.; Siadat, S.A.; Bakhshandeh, A.; Ghasemi Pirbalouti, A.; Hashemi, M. Interactive effects of drought stress and chitosan application on physiological characteristics and essential oil yield of Thymus daenesis Celak. Crop J. 2017, 5, 407–415. [Google Scholar] [CrossRef]
- Yin, H.; Frette, X.C.; Christensen, L.P.; Grevsen, K. Chitosan oligosaccharides promote the content of polyphenols in Greek oregano (Origanum vulgare ssp. hirtum). J. Agric. Food Chem. 2012, 60, 136–143. [Google Scholar] [CrossRef]
- Alizadeh, A.; Moghaddam, M.; Asgharzade, A.; Mahmoodi Sourestani, M. Phytochemical and physiological response of Satureja hortensis L. to different irrigation regimes and chitosan application. Ind. Crops Prod. 2020, 158, 112990. [Google Scholar] [CrossRef]
- Panwar, J.; Tarafdar, J.C. Arbuscular mycorrhizal fungal dynamics under Mitragyna parvifolia (Roxb.) Korth. in thar desert. Appl. Soil Ecol. 2006, 34, 200–208. [Google Scholar] [CrossRef]
- Abdollahi, A.A.; Feizian, M.; Mehdipourian, G.; Khojasteh, D.N. Arbuscular mycorrhizal fungi inoculation improve essential oil and physiological parameters and nutritional values of Thymus daenensis Celak and Thymus vulgaris L. under normal and drought stress conditions. Eur. J. Soil Biol. 2020, 100, 103217. [Google Scholar] [CrossRef]
- Hashem, A.; Kumar, A.; Al-Dbass, A.M.; Alqarawi, A.A.; Al-Arjani, A.B.F.; Singh, G.; Farooq, M.; Abd_Allah, E.F. Arbuscular mycorrhizal fungi and biochar improves drought tolerance in chickpea. Saudi J. Biol. Sci. 2019, 26, 614–624. [Google Scholar] [CrossRef]
- Silva, V.C.; Casaes Alves, F.A.; Oliveira, R.A.; Jesus, R.M.; Bomfim Costa, L.C.; Gross, E. Influence of arbuscular mycorrhizal fungi on growth, mineral composition and production of essential oil in Mentha × piperita L. var. citrata (Ehrh.) Briq. under two phosphorus levels. J. Med. Plant Res. 2014, 8, 1321–1332. [Google Scholar] [CrossRef]
- Benaffari, W.; Boutasknit, A.; Anli, M.; Ait-El-Mokhtar, M.; Ait-Rahou, Y.; Ben-Laouane, R.; Ahmed, H.; Mitsui, T.; Baslam, M.; Meddich, A. The native arbuscular mycorrhizal fungi and vermicompost-based organic amendments enhance soil fertility, growth performance, and the drought stress tolerance of quinoa. Plants 2022, 11, 393. [Google Scholar] [CrossRef]
- Mathur, S.; Tomar, R.S.; Jajoo, A. Arbuscular mycorrhizal fungi (AMF) protects photosynthetic apparatus of wheat under drought stress. Photosynth. Res. 2019, 139, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Pirzad, A.; Mohammadzadeh, S.J. Water use efficiency of three mycorrhizal Lamiaceae species (Lavandula officinalis, Ros-ma-rinus officinalis and Thymus vulgaris). Agric. Water Manag. 2018, 204, 1–10. [Google Scholar] [CrossRef]
- Khorasaninejad, S.; Mousavi, A.; Soltanloo, H.; Hemmati, K.; Khalighi, A. The effect drought stress on growth parameters, essential oil yield and constituent of pepprmint (Mentha piperita L.). J. Med. Plant Res. 2011, 22, 5360–5365. [Google Scholar]
- Arnon, A. Method of extraction of chlorophyll in the plants. Agron. J. 1967, 23, 112–121. [Google Scholar] [CrossRef]
- Morshedloo, M.R.; Craker, L.E.; Salami, A.; Nazeri, V.; Sang, H.; Maggi, F. Effect of prolonged water stress on essential oil content, compositions and gene expression patterns of mono-and sesquiterpene synthesis in two oreganos (Origanum vulgare L.) subspecies. Plant Physiol. Biochem. 2017, 111, 119–128. [Google Scholar] [CrossRef]
- Safaei, L.; Sharifi Ashorabadi, E.; Afyouni, D. The effects npk, chemical and manure fertilizers investigation on the phenolic yield and essential oil components in Thymus daenensis L. EJMP 2017, 17, 1–15. [Google Scholar]
- Poshtdar, A.; Mashhadie, A.R.A.; Moradi, F.; Siadat, S.A.; Bakhshandeh, A. Effect of source and rate of nitrogen fertilizer on yield and water and nitrogen use efficiency of peppermint (Mentha piperita L.). Iran. J. Crop Sci. 2016, 18, 14–31. [Google Scholar]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Quadrupole Mass Spectrometry; Allured Publishing Corporation: Carroll Stream, IL, USA, 2007; Volume 456, pp. 544–545. [Google Scholar]
- Morshedloo, M.R.; Maggi, F.; Neko, H.T.; Aghdam, M.S. Sumac (Rhus coriaria L.) fruit: Essential oil variability in iranian populations. Ind. Crops Prod. 2018, 111, 1–7. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Beauchamp, C.; Fridovich, I. Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels. Anal. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar] [CrossRef]
- Kumar, K.B.; Khan, P.A. Peroxidase & Polyphenol Oxidase in Excised Ragi (Eleusine Corocana Cv PR 202) Leaves during Senescence. Indian J. Exp. Biol. 1982, 20, 412–416. [Google Scholar]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence—A practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef]
- Vargas-Ortiz, E.; Ramírez-Tobias, H.M.; González-Escobar, J.L.; Gutiérrez-García, A.K.; Bojórquez-Velázquez, E.; Espitia-Rangel, E.; Barba de la Rosa, A.P. Biomass, chlorophyll fluorescence, and osmoregulation traits let differentiation of wild and cultivated Amaranthus under water stress. J. Photochem. Photobiol. B Biol. 2021, 220, 112210. [Google Scholar] [CrossRef]
- Albouchi, A.; Bejaoui, Z.; El Aouni, M.H. Influence of moderate or severe water stress on the growth of Casuarina glauca Sieb. Seedlings. Sécheresse 2003, 14, 137–142. [Google Scholar]
- Bahreininejad, B.; Razmjoo, J.; Mirza, M. Influence of water stress on morphophysiological and phytochemical traits in (Thymus daenensis). Int. J. Plant Prod. 2013, 7, 152–166. [Google Scholar]
- Tabrizi, L.; Koocheki, A.; Rezvani Moghaddam, P.; Nasiri Mahallati, M.; Bannayan, M. Effect of irrigation and organic manure on Khorasan thyme (Thymus transcaspicus Klokov). Arch. Agron. Soil Sci. 2011, 57, 317–326. [Google Scholar] [CrossRef]
- Abd El-Azeim, M.M.; Sherif, M.A.; Hussien, M.S.; Tantawy, I.A.A.; Bashandy, S.O. Impacts of nano- and non-nanofertilizers on potato quality and productivity. Acta Ecol. Sin. 2020, 40, 388–397. [Google Scholar] [CrossRef]
- Ghasemi Pirbalouti, A.; Malekpoor, F.; Salimi, A.; Golparvar, A. Exogenous application of chitosan on biochemical and physiological characteristics, phenolic content and antioxidant activity of two species of basil (Ocimum ciliatum and Ocimum basilicum) under reduced irrigation. Sci. Hortic. 2017, 217, 114–122. [Google Scholar] [CrossRef]
- Zhao, Y.; Cartabia, A.; Lalaymia, I.; Declerck, S. Arbuscular mycorrhizal fungi and production of secondary metabolites in medicinal plants. Mycorrhiza 2022, 32, 221–256. [Google Scholar] [CrossRef]
- Minaei, A.; Hassani, A.; Nazemiyeh, H.; Besharat, S. Effect of drought stress on some morphophysiological and phytochemical characteristics of oregano (Origanum vulgare L. ssp. gracile). IJMAPR 2019, 35, 252–265. [Google Scholar]
- Gholinezhad, E.; Darvishzadeh, R. Influence of arbuscular mycorrhiza fungi and drought stress on fatty acids profile of sesame (Sesamum indicum L.). Field Crops Res. 2021, 262, 108035. [Google Scholar] [CrossRef]
- Amani Machiani, M.; Javanmard, A.; Morshedloo, M.R.; Janmohammadi, M.; Maggi, F. Funneliformis mosseae Application Improves the Oil Quantity and Quality and Ecophysiological Characteristics of Soybean (Glycine max L.) Under Water Stress Conditions. J. Plant Nutr. Soil Sci. 2021, 21, 3076–3090. [Google Scholar] [CrossRef]
- Limpanavech, P.; Chaiyasuta, S.; Vongpromek, R.; Pichyangkura, R.; Khunwasi, C.; Chadchawan, S. Chitosan effects on floral production, gene expression, and anatomical changes in the Dendrobium orchid. Sci. Hortic. 2008, 116, 65–72. [Google Scholar] [CrossRef]
- Ali, E.F.; El-Shehawi, A.M.; Ibrahim, O.H.M.; Abdul-Hafeez, E.Y.; Moussa, M.M.; Hassan, F.A.S. A vital role of chitosan nanoparticles in improvisation the drought stress tolerance in Catharanthus roseus (L.) through biochemical and gene expression modulation. Plant Physiol. Biochem. 2021, 161, 166–175. [Google Scholar] [CrossRef]
- Van, S.; Dinh Minh, H.; Nguyen Anh, D. Study on chitosan nanoparticles on biophysical characteristics and growth of Robusta coffee in greenhouse. Biocatal. Agric. Biotechnol. 2013, 2, 289–294. [Google Scholar] [CrossRef]
- Baker, N.R. Chlorophyll fluorescence: A probe of photosynthesis in vivo. Annul. Rev. Plant Biol. 2008, 59, 89–113. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.; Tian, Y.; Zhang, B.; Hassan, M.J.; Li, Z.; Zhu, Y. Chitosan (CTS) Alleviates Heat-Induced Leaf Senescence in Creeping Bentgrass by Regulating Chlorophyll Metabolism, Antioxidant Defense, and the Heat Shock Pathway. Molecules 2021, 26, 5337. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Zhang, C.; Shi, S.; Wang, B.; Zhao, J. Physiological and biochemical changes in different drought-tolerant alfalfa (Medicago sativa L.) varieties under PEG-induced drought stress. Acta Physiol. Plant 2018, 40, 25. [Google Scholar] [CrossRef]
- Dutta, T.; Neelapu, N.R.; Wani, S.H.; Challa, S. Compatible Solute Engineering of Crop Plants for Improved Tolerance toward Abiotic Stresses. Biochemical, Physiological and Molecular Avenues for Combating Abiotic Stress Tolerance in Plants; Academic Press: Cambridge, MA, USA, 2018; pp. 221–254. [Google Scholar]
- Pryor, W.A.; Stanley, J.P. A suggested mechanism for the production of malonaldehyde during the antioxidation of polyunsaturated fatty acids, nonenzymatic production of prostaglandin endoperoxides during autoxidation. Organells 1975, 40, 3615–3617. [Google Scholar] [CrossRef]
- Guo, Y.Y.; Yu, H.Y.; Yang, M.M.; Kong, D.S.; Zhang, Y.J. Effect of Drought Stress on Lipid Peroxidation, Osmotic Adjustment and Antioxidant Enzyme Activity of Leaves and Roots of Lycium ruthenicum Murr. Seedling. Russ. J. Plant Physiol. 2018, 65, 244–250. [Google Scholar] [CrossRef]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S.M.A. Plant drought stress: Effects, mechanisms and management. Agron. Sustain. Dev. 2009, 29, 185–212. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
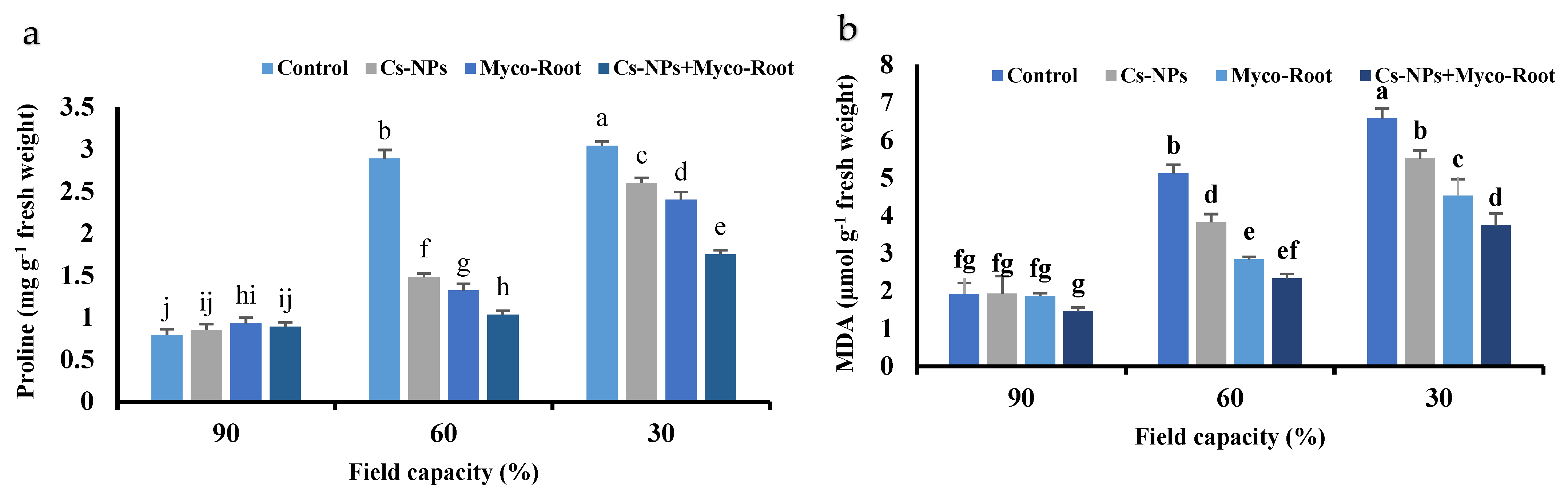{kind=link}
{kind=link}
| Treatments | Fresh Weight (g pot−1) | Dry Weight (g pot−1) | Chlorophyll a (mg g−1 Fresh Weight) | Chlorophyll b (mg g−1 Fresh Weight) | Total Chlorophyll (mg g−1 Fresh Weight) | Carotenoid (mg g−1 Fresh Weight) | Essential Oil Content (%) | Essential Oil Yield (g pot−1) | |
|---|---|---|---|---|---|---|---|---|---|
| 90% FC | Control | 42.67 ± 1.69 d | 15.06 ± 0.52 d | 10.03 ± 0.61 d | 6.35 ± 0.34 d | 16.38 ± 0.77 d | 4.03 ± 0.06 c | 0.16 ± 0.02 j | 0.02 ± 0.001 j |
| Cs-NPs | 51.33 ± 1.24 c | 17.45 ± 0.21 c | 11.59 ± 0.28 c | 7.52 ± 0.05 c | 19.11 ± 0.26 c | 4.19 ± 0.02 c | 0.27 ± 0.03 i | 0.04 ± 0.002 i | |
| Myco-Root | 59.00 ± 133 b | 20.65 ± 0.31 b | 12.07 ± 0.12 b | 8.09 ± 0.07 b | 20.16 ± 0.17 b | 4.49 ± 0.06 b | 0.32 ± 0.03 i | 0.06 ± 0.003 g,h | |
| Cs-NPs+ Myco-Root | 62.33 ± 0.47 a | 21.63 ± 0.09 a | 13.31 ± 0.07 a | 8.87 ± 0.04 a | 22.19 ± 0.13 a | 5.00 ± 0.18 a | 0.42 ± 0.08 h | 0.09 ± 0.001 e,f | |
| 60% FC | Control | 38.67 ± 1.81 e | 12.87 ± 0.15 e | 6.66 ± 0.12 g | 4.44 ± 0.05 h | 11.11 ± 0.14 h | 2.12 ± 0.04 i | 0.78 ± 0.04 e | 0.10 ± 0.006 e |
| Cs-NPs | 44.00 ± 1.94 d | 15.15 ± 0.45 d | 7.05 ± 0.19 g | 4.70 ± 0.05 g | 11.77 ± 0.12 g | 3.24 ± 0.02 e | 1.05 ± 0.06 c | 0.15 ± 0.003 c | |
| Myco-Root | 49.33 ± 1.47 c | 17.27 ± 0.41 c | 7.68 ± 0.11 f | 5.12 ± 0.07 f | 12.81 ± 0.19 f | 3.55 ± 0.03 d | 1.18 ± 0.07 b | 0.20 ± 0.001 b | |
| Cs-NPs+ Myco-Root | 51.33 ± 1.65 c | 18.07 ± 0.22 c | 8.37 ± 0.22 e | 5.64 ± 0.09 e | 14.02 ± 0.30 e | 3.70 ± 0.04 d | 1.43 ± 0.04 a | 0.25 ± 0.004 a | |
| 30% FC | Control | 27.33 ± 1.92 g | 9.60 ± 0.63 f | 4.11 ± 0.05 i | 2.74 ± 0.03 k | 6.85 ± 0.09 k | 1.68 ± 0.11 j | 0.56 ± 0.03 g | 0.05 ± 0.002 h,i |
| Cs-NPs | 34.33 ± 2.04 f | 12.22 ± 0.47 e | 5.31 ± 0.03 h | 3.54 ± 0.02 j | 8.86 ± 0.15 j | 2.31 ± 0.31 h | 0.66 ± 0.04 f | 0.08 ± 0.004 f,g | |
| Myco-Root | 38.00 ± 2.16 e | 13.05 ± 0.58 e | 5.68 ± 0.16 h | 3.790.16 i | 9.47 ± 0.27i | 2.64 ± 0.07 g | 0.66 ± 0.03 f | 0.08 ± 0.005 e,f | |
| Cs-NPs+ Myco-Root | 42.67 ± 0.74 d | 14.33 ± 0.36 d | 6.78 ± 0.04 g | 4.520.02 g,h | 11.31 ± 0.06 g,h | 3.04 ± 0.12 f | 0.88 ± 0.06 d | 0.12 ± 0.007 d | |
| Source of Variation | Significance Levels | ||||||||
| Irrigation | ** | ** | ** | ** | ** | ** | ** | ** | |
| Fertilizer sources | ** | ** | ** | ** | ** | ** | ** | ** | |
| Irrigation × Fertilizer sources | ** | ** | ** | ** | ** | ** | ** | ** | |
| Constituents | RI | 90% FC Control | 90% FC Cs-NPs | 90% FC Myco-Root | 90% FC Cs-NPs+ Myco-Root | 60% FC Control | 60% FC Cs-NPs | 60% FC Myco-Root | 60% FC Cs-NPs+ Myco-Root | 30% FC Control | 30% FC Cs-NPs | 30% FC Myco-Root | 30% FC Cs-NPs+ Myco-Root | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | α-Pinene | 932 | 0.78 | 0.81 | 1.03 | 1.09 | 0.87 | 1.22 | 1.14 | 1.06 | 0.92 | 0.91 | 1.12 | 1.1 |
| 2 | Sabinene | 969 | 1.66 | 1.19 | 1.52 | 1.71 | 1.44 | 1.51 | 1.55 | 1.14 | 1.23 | 1.44 | 1.52 | 1.38 |
| 3 | β-Myrcene | 988 | 1.63 | 2.32 | 3.01 | 2.88 | 1.83 | 1.91 | 2.98 | 2.63 | 3.06 | 3.11 | 2.96 | 3.02 |
| 4 | Limonene | 1024 | 0.53 | 0.46 | 0.12 | 0.25 | 0 | 0.28 | 0.22 | 0.55 | 0.69 | 0.3 | 0.53 | 0 |
| 5 | Linalool | 1096 | 0.31 | 0.33 | 0.37 | 0.43 | 0.81 | 0.23 | 0.97 | 0.32 | 0 | 0.41 | 0 | 0 |
| 6 | n-Dodecane | 1200 | 0.17 | 0.28 | 0.29 | 0.18 | 0.36 | 0.7 | 0.21 | 0.4 | 0.41 | 0.36 | 0.17 | 0.29 |
| 7 | Neral | 1235 | 2.28 | 0.14 | 0.1 | 2.66 | 1.36 | 0 | 0.14 | 0.66 | 0 | 0.16 | 0 | 0.12 |
| 8 | Menthyl acetate | 1294 | 2.26 | 3.15 | 3.56 | 3.78 | 2.41 | 2.04 | 0.92 | 1.29 | 2.04 | 1.55 | 1.18 | 0.75 |
| 9 | Unknown | - | 1.72 | 1.4 | 1.7 | 1.58 | 1.71 | 1.24 | 1.59 | 1.29 | 1.66 | 1.46 | 1.71 | 2.26 |
| 10 | β-Elemene | 1389 | 1.42 | 1.06 | 0.88 | 1.46 | 1.22 | 0.71 | 1.56 | 1.13 | 0.41 | 0.69 | 0.77 | 1.08 |
| 11 | (E)-Caryophyllene | 1417 | 16.28 | 17.15 | 18.39 | 19.01 | 17.23 | 18.73 | 18.04 | 19.82 | 16.54 | 17.22 | 17.76 | 18.61 |
| 12 | α-Humulene | 1452 | 3.43 | 3.28 | 3.74 | 3.66 | 2.56 | 2.1 | 3.29 | 2.17 | 3.55 | 2.86 | 0 | 3.11 |
| 13 | Germacrene D | 1484 | 31.22 | 35.25 | 34.95 | 33.61 | 32.14 | 39.16 | 34.53 | 39.77 | 34.47 | 38.83 | 37.75 | 36.2 |
| 14 | Valencene | 1496 | 2.49 | 2.12 | 2.58 | 2.2 | 2.71 | 1.18 | 2.07 | 0.87 | 1.96 | 1.73 | 2.21 | 3.07 |
| 15 | Bicyclogermacrene | 1500 | 8.23 | 7.1 | 8.5 | 7.86 | 8.2 | 8.59 | 8.83 | 9.07 | 8.84 | 8.26 | 9.04 | 9.22 |
| 16 | δ-cadinene | 1522 | 0.94 | 1.23 | 0.28 | 0.24 | 0.71 | 0.08 | 0 | 0 | 0.39 | 0 | 0.27 | 0.68 |
| 17 | β-Bisabolene | 1505 | 1.86 | 2.22 | 1.31 | 1.89 | 2.06 | 1.14 | 1.63 | 1.36 | 1.44 | 1.12 | 1.2 | 1.51 |
| 18 | Spathulenol | 1577 | 2.43 | 1.78 | 2.06 | 2.38 | 1.07 | 1.66 | 3.34 | 1.93 | 2.23 | 2.04 | 2.27 | 2.38 |
| 19 | Caryophyllene oxide | 1582 | 4.23 | 4 | 3.85 | 4.02 | 4.91 | 4.14 | 4.02 | 6.15 | 5.78 | 4.19 | 6.96 | 5.63 |
| 20 | Phytol | 1942 | 1.61 | 0.74 | 0.98 | 1.88 | 1.74 | 0.45 | 0.57 | 0.84 | 1.23 | 0.49 | 0.64 | 0.8 |
| Total identified (%) | 85.48 | 86.01 | 89.22 | 92.77 | 85.34 | 87.07 | 87.6 | 92.45 | 86.85 | 87.13 | 88.06 | 91.21 | ||
| Sources of Variation | df | Leaf Greenness (SPAD Index) | F0 | Fm | Fv | Fv/Fm | Proline | MDA | SOD | POX | APX |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Irrigation | 2 | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** |
| Fertilizer | 3 | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** |
| Irrigation regimes × Fertilizer | 6 | * | Ns | ** | ** | * | ** | ** | ** | ** | ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Javanmard, A.; Ashrafi, M.; Morshedloo, M.R.; Machiani, M.A.; Rasouli, F.; Maggi, F. Optimizing Phytochemical and Physiological Characteristics of Balangu (Lallemantia iberica) by Foliar Application of Chitosan Nanoparticles and Myco-Root Inoculation under Water Supply Restrictions. Horticulturae 2022, 8, 695. https://doi.org/10.3390/horticulturae8080695
Javanmard A, Ashrafi M, Morshedloo MR, Machiani MA, Rasouli F, Maggi F. Optimizing Phytochemical and Physiological Characteristics of Balangu (Lallemantia iberica) by Foliar Application of Chitosan Nanoparticles and Myco-Root Inoculation under Water Supply Restrictions. Horticulturae. 2022; 8(8):695. https://doi.org/10.3390/horticulturae8080695
Chicago/Turabian StyleJavanmard, Abdollah, Mahsa Ashrafi, Mohammad Reza Morshedloo, Mostafa Amani Machiani, Farzad Rasouli, and Filippo Maggi. 2022. "Optimizing Phytochemical and Physiological Characteristics of Balangu (Lallemantia iberica) by Foliar Application of Chitosan Nanoparticles and Myco-Root Inoculation under Water Supply Restrictions" Horticulturae 8, no. 8: 695. https://doi.org/10.3390/horticulturae8080695