
The Link between Mineral Elements Variation and Internal Flesh Breakdown of ‘Keitt’ Mango in a Steep Slope Mountain Area, Southwest China

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area Description
2.2. Fruit, Leaf and Soil Sampling
2.3. Analytical Methods
2.4. Statistical Analysis
3. Results
3.1. IFB Incidence in Mango Orchards
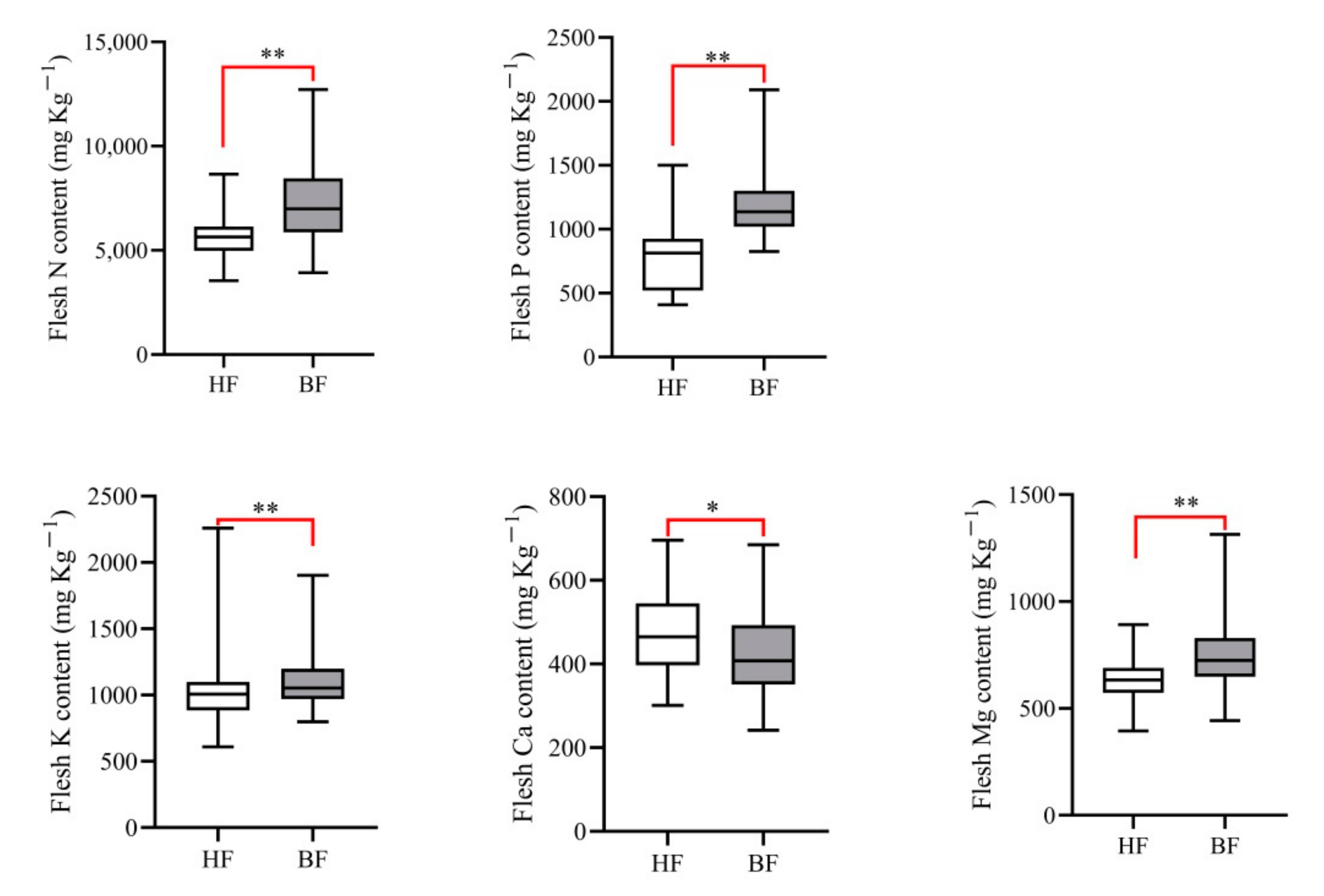
3.2. Variation of Mineral Element Concentrations between the Flesh of Healthy Fruit and Fruit with IFB
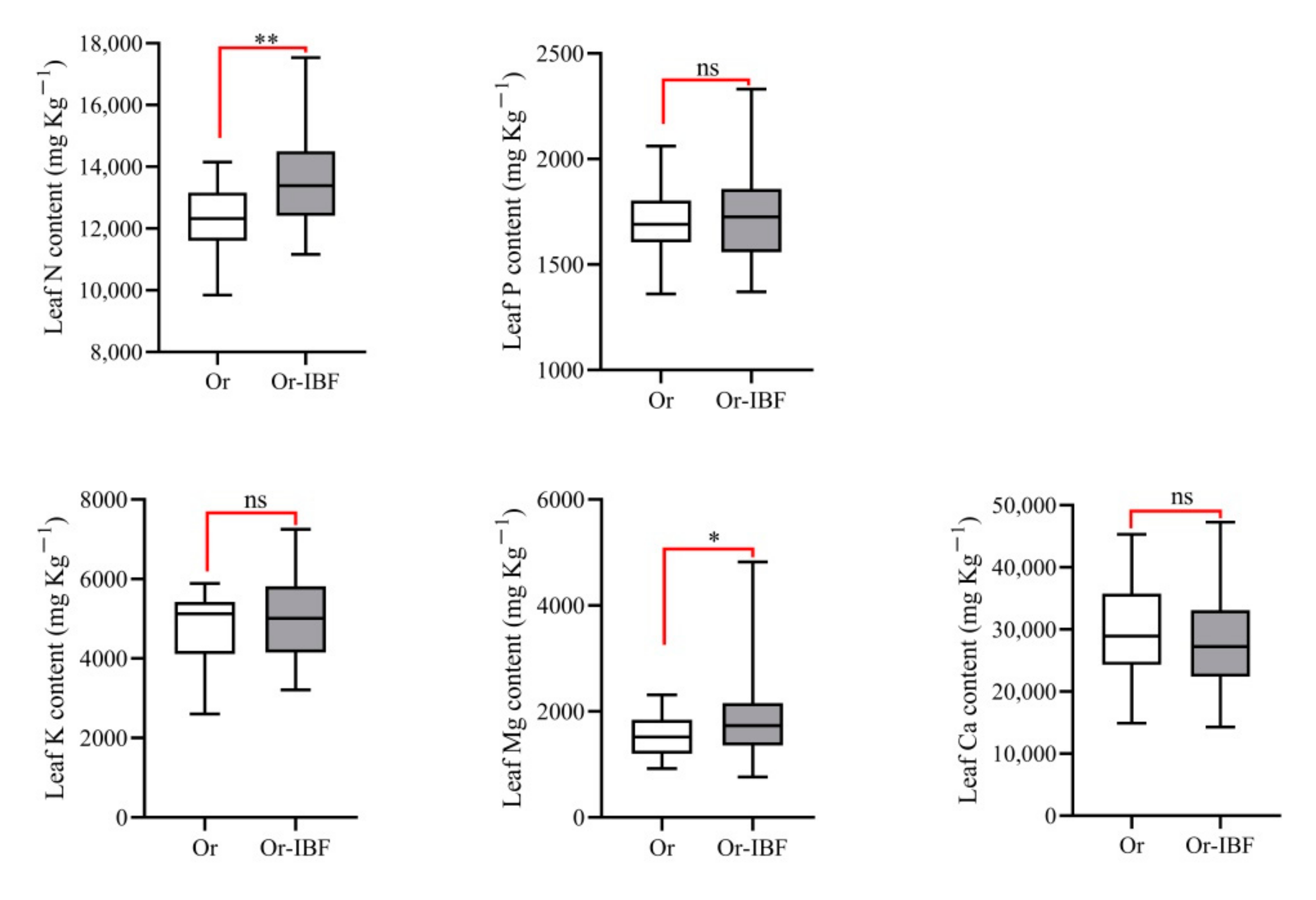
3.3. Variation in Leaf Mineral Element Concentrations between Orchards with and without IFB Fruit
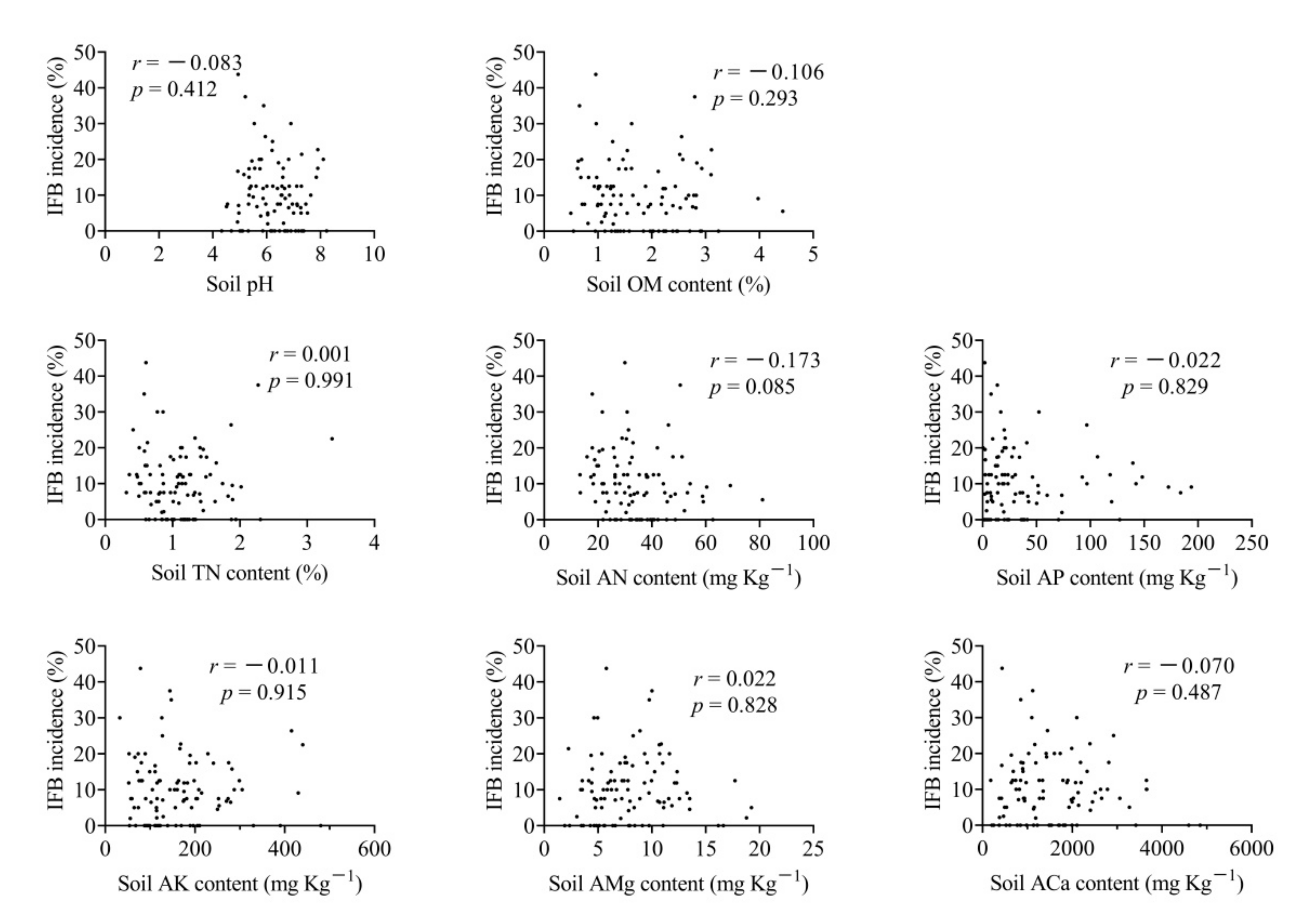
3.4. Variation of Soil pH and Nutrient Contents between Orchard with and without IFB Fruit
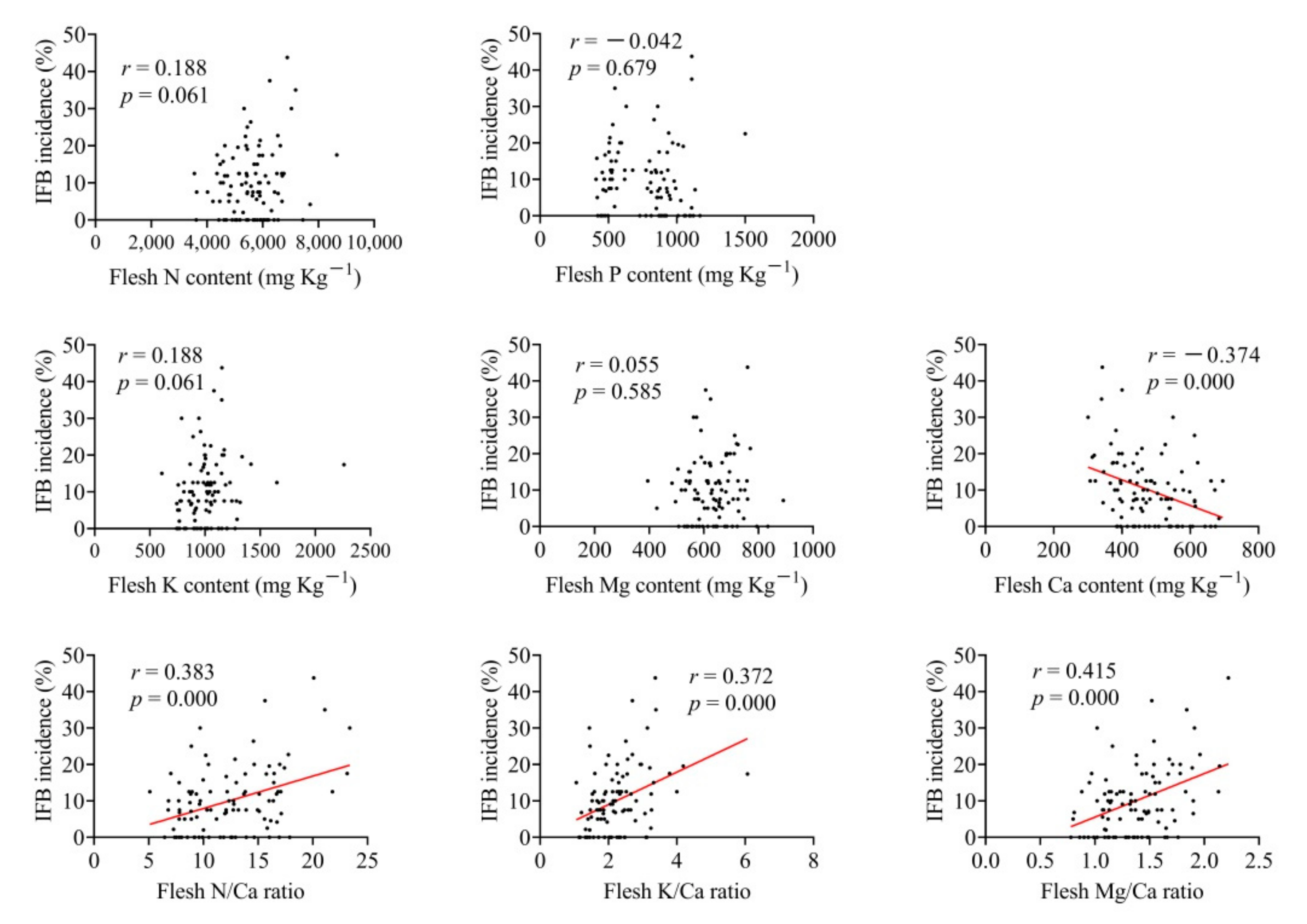
3.5. Correlation Analysis of Nutrient Concentration in Fruit Flesh, Leaf and Soil
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lagunes-Delgado, C.; Agama-Acevedo, E.; Patiño-Rodríguez, O.; Martinez, M.M.; Bello-Pérez, L.A. Recovery of mango starch from unripe mango juice. LWT 2022, 153, 112514. [Google Scholar] [CrossRef]
- Raymond, L.; Schaffer, B.; Brecht, B.J.; Jonathan, H. Internal breakdown in mango fruit: Symptomology and histology of jelly seed, soft nose and stem-end cavity. Postharvest Biol. Technol. 1998, 13, 59–70. [Google Scholar] [CrossRef]
- Mogollón-Lancheros, M.R.; Freitas, S.T.D.; Zoffoli, J.P.; Contreras, C.; Neta, M.L.D.S. Non-destructive prediction and detection of internal physiological disorders in ‘Keitt’ mango using a hand-held Vis-NIR spectrometer. Postharvest Biol. Technol. 2020, 167, 11251. [Google Scholar]
- Machhindra, T.; Sharma, J.A. Spongy tissue development in Alphonso mango: Association with Staphylococcus xylosus. Eur. J. Plant Pathol. 2008, 122, 355. [Google Scholar]
- Muhammad, A.; Ahmad, I.; Malik, A.U.; Nawab, U.; Jabbar, A. Mango soft nose disorder and fruit quality in relation to pre and postharvest treatments. Life Sci. Int. J. 2007, 1, 455–462. [Google Scholar]
- Shivashankara, K.S.; Mathai, C.K. Relationship of leaf and fruit transpiration rates to the incidence of spongy tissue disorder in two mango (Mangifera indica L.) cultivars. Sci. Hortic. 1999, 82, 317–323. [Google Scholar] [CrossRef]
- Young, T.W. ‘Soft-nose’, a physiological disorder in mango fruits. Proc. Fla. State Hortic. Soc. 1957, 23, 259–261. [Google Scholar]
- Subraman, H.; Krishnam, S.; Subhadra, N.V.; Dalal, V.B.; Randhawa, G.S.; Chacko, E.K. Studies on internal breakdown, a physiological ripening disorder in Alphonso mangoes (Mangifera indica L.). Trop. Sci. 1971, 13, 203–210. [Google Scholar]
- Raymond, L.; Schaffer, B.; Brecht, J.K.; Hanlone, E.A. Internal breakdown, mineral element concentration, and weight of mango fruit. J. Plant Nutr. 1998, 21, 871–889. [Google Scholar] [CrossRef]
- Ma, X.W.; Yao, Q.S.; Ma, H.Y.; Wu, H.X.; Zhou, Y.G.; Wang, S.B. Relationship between internal breakdown and mineral nutrition in the flesh of ‘Keitt’ mango. Acta Hortic. 2018, 1217, 351–356. [Google Scholar] [CrossRef]
- Liu, X.G.; Zhang, Y.Y.; Leng, X.X.; Yang, Q.L.; Chen, H.Q.; Wang, X.K.; Cui, N.B. Exploring the optimisation of mulching and irrigation management practices for mango production in a dry hot environment based on the entropy weight method. Sci. Hortic. 2022, 291, 110564. [Google Scholar] [CrossRef]
- Sun, G.Z.; Hu, T.T.; Liu, X.G.; Peng, Y.L.; Leng, X.X.; Li, Y.L.; Yang, Q.L. Optimizing irrigation and fertilization at various growth stages to improve mango yield, fruit quality an water-fertilizer use efficiency in xerothermic regions. Agric. Water Manag. 2022, 260, 107296. [Google Scholar] [CrossRef]
- Bao, S.D. Soil Agrochemical Analysis, 3rd ed.; China Agriculture Press: Beijing, China, 2000; pp. 432–470. [Google Scholar]
- Wang, Y.L.; Zhang, X.F.; Wang, Y.Z.; Yang, S.L.; Qu, H.Y. The changes of intracellular calcium concentration and distribution in the hard end pear (Pyrus pyrifolia cv. ‘Whangkeumbae’) fruit. Cell Calcium 2018, 71, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Chen, J.Z. Citrus Fruit-Cracking: Causes and Occurrence. Hortic. Plant J. 2017, 3, 255–260. [Google Scholar] [CrossRef]
- Torres, E.; Recasens, I.; Àvila, G.; Lordan, J.; Alegre, S. Early stage fruit analysis to detect a high risk of bitter pit in ‘Golden Smoothee’. Sci. Hortic. 2017, 219, 98–106. [Google Scholar] [CrossRef] [Green Version]
- De Bang, T.C.; Husted, S.; Laursen, K.H.; Persson, D.P.; Schjoerring, J.K. The molecular–physiological functions of mineral macronutrients and their consequences for deficiency symptoms in plants. New Phytol. 2021, 229, 2446–2469. [Google Scholar] [CrossRef]
- Li, Q.; Cheng, C.X.; Zhang, X.F.; Wang, C.H.; Yang, S.L. Preharvest bagging and postharvest calcium treatment affects superficial scald incidence and calcium nutrition during storage of ‘Chili’ pear (Pyrus bretschneideri) fruit. Postharvest Biol. Technol. 2020, 163, 111149. [Google Scholar] [CrossRef]
- Zhang, Y.H.; Su, M.Q.; Liu, B.; Wang, S.B.; Wang, J.F.; Ma, X.W. Analysis of relationship between internal flesh breakdown and calcium in mango ‘Keitt’ fruit. South Chin. Fruits 2021, 50, 82–86. [Google Scholar]
- Tao, S.H. Effects of Calcium Treatment on Soft Nose and Storability of Pre-Bagging ‘Keitt’ Mango Fruit during Storage; Zhejiang Gongshang University: Hangzhou, China, 2018. [Google Scholar]
- Huang, X.M.; Wang, H.C.; Zhong, W.L.; Yuan, W.Q.; Lu, J.M.; Li, J.G. Spraying calcium is not an effective way to increase structural calcium in litchi pericarp. Sci. Hortic. 2008, 117, 39–44. [Google Scholar] [CrossRef]
- Bangerth, F. Calcium-related physiological disorder in plants. Annu. Rev. Phytopathol. 1979, 17, 97–122. [Google Scholar] [CrossRef]
- Hocking, B.; Tyerman, S.D.; Burton, R.A.; Gilliham, M. Fruit calcium: Transport and physiology. Front. Plant Sci. 2016, 29, 569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.; Kim, J.; Choi, J.P.; Lee, M.; Kim, M.K.; Lee, Y.H.; Hur, Y.; Nou, S.; Park, S.U.; Min, S.R.; et al. Intracellular Ca2+ and K+ concentration in Brassica oleracea leaf nduces differential expression of transporter and stress-related genes. BMC Genom. 2016, 17, 211. [Google Scholar] [CrossRef] [Green Version]
- Song, W.P.; Yi, J.W.; Kurniadinata, O.F.; Wang, H.C.; Huang, X.M. Linking fruit Ca uptake capacity to fruit growth and pedicel anatomy, a cross-species study. Front. Plant Sci. 2018, 9, 575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Djangsou, H.; Enrico, F.; Domenico, R.; Matteo, B. Blossom end-rot in tomato (Solanum lycopersicum L.): A multi-disciplinary overview of inducing factors and control strategies. Sci. Hortic. 2019, 249, 49–58. [Google Scholar]
- Sete, P.B.; Comin, J.J.; Ciotta, M.C.; Salume, J.A.; Thewes, F.; Brackman, A.; Toselli, M.; Nava, G.; Rozane, D.E.; Loss, A.; et al. Nitrogen fertilization affects yield and fruit quality in pear. Sci. Hortic. 2019, 258, 108782. [Google Scholar] [CrossRef]
- Ho, L.C.; White, P.J. A cellular hypothesis for the induction of blossom-end rot in tomato fruit. Ann. Bot. 2005, 95, 571–581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, Z.H.; Jiao, Q.Y.; Wang, R.; Ma, C.H. Investigation and analysis of relationship between mineral elements alteration and cork spot physiological disorder of Chinese pear ‘Chili’ (Pyrus bretschneideri Rehd.). Sci. Hortic. 2020, 260, 108883. [Google Scholar] [CrossRef]
- Saquet, A.A.; Streifc, J.; Alemidda, D.P.F. Mineral composition and distribution within ‘Rocha’ pear in relation to internal storage disorders. Postharvest Biol. Technol. 2019, 158, 111002. [Google Scholar] [CrossRef]
- De Freitas, S.T.; do Amarante, C.V.T.; Labavitch, J.M.; Mitcham, E.J. Cellular approach to understand bitter pit development in apple fruit. Postharvest Biol. Technol. 2010, 57, 6–13. [Google Scholar] [CrossRef]
- De Freitas, S.T.; Handa, A.K.; Wu, Q.Y.; Park, S.H.; Mitcham, E.J. Role of pectin methylesterases in cellular calcium distribution and blossom-end rot development in tomato fruit. Plant J. 2012, 71, 824–835. [Google Scholar] [CrossRef]
- Jiang, F.L.; Lopez, A.; Jeon, S.; de Freitas, S.T.; Yu, Q.H.; Zhen, W.; Labavitch, J.M.; Tian, S.; Powell, A.L.T.; Mitcham, E. Disassembly of the fruit cell wall by the ripening-associated polygalacturonase and expansin influences tomato cracking. Hortic. Res. 2019, 6, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ge, S.F.; Zhu, Z.L.; Peng, L.; Chen, Q.; Jiang, Y.M. Soil Nutrient Status and Leaf Nutrient Diagnosis in the Main Apple Producing Regions in China. Hortic. Plant J. 2018, 4, 89–93. [Google Scholar] [CrossRef]
- Polverigiani, S.; Franzina, M.; Neri, D. Effect of soil condition on apple root development and plant resilience in intensive orchards. Appl. Soil Ecol. 2018, 123, 787–792. [Google Scholar] [CrossRef]
- Li, Z.G.; Zhang, R.H.; Xia, S.J.; Wang, L.; Liu, C.; Zhang, R.Q.; Fan, Z.H.; Chen, F.; Liu, Y. Interactions between N, P and K fertilizers affect the environment and the yield and quality of satsumas. Glob. Ecol. Conserv. 2019, 19, e00663. [Google Scholar] [CrossRef]
- Wang, Y.D.; Liu, C.; Cui, P.F.; Su, D.R. Effects of partial root-zone drying on alfalfa growth, yield and quality under subsurface drip irrigation. Agric. Water Manag. 2021, 245, 106608. [Google Scholar] [CrossRef]
- Elhani, S.; Haddai, M.; Csákvári, E.; Zantar, S.; Hamim, A.; Villányi, V.; Douaik, A.; Bánfalvi, V. Effects of partial roo-zone drying and deficit irrigation water-use efficiency and some potato (Solanum tuberosum L.) quality traits under glasshouse conditions. Agric. Water Manag. 2019, 224, 105745. [Google Scholar] [CrossRef]
- Ghafari, H.; Hassanpou, H.; Jafari, M.; Besharat, S. Physiological, biochemical and gene-expressional responses to water deficit in apple subjected to partial-zone drying (PRD). Plant Physiol. Biochem. 2020, 148, 333–346. [Google Scholar] [CrossRef]
- Wang, L.; de Kroon, H.; Smits, A.J.M. Combined effects of partial root drying and patchy fertilizer placement on nutrient acquisition and growth of oilseed rape. Plant Soil. 2007, 295, 207–216. [Google Scholar] [CrossRef]














Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, X.; Wang, J.; Su, M.; Liu, B.; Du, B.; Zhang, Y.; He, L.; Wang, S.; Wu, H. The Link between Mineral Elements Variation and Internal Flesh Breakdown of ‘Keitt’ Mango in a Steep Slope Mountain Area, Southwest China. Horticulturae 2022, 8, 533. https://doi.org/10.3390/horticulturae8060533
Ma X, Wang J, Su M, Liu B, Du B, Zhang Y, He L, Wang S, Wu H. The Link between Mineral Elements Variation and Internal Flesh Breakdown of ‘Keitt’ Mango in a Steep Slope Mountain Area, Southwest China. Horticulturae. 2022; 8(6):533. https://doi.org/10.3390/horticulturae8060533
Chicago/Turabian StyleMa, Xiaowei, Jianfang Wang, Muqing Su, Bin Liu, Bang Du, Yuehua Zhang, Liping He, Songbiao Wang, and Hongxia Wu. 2022. "The Link between Mineral Elements Variation and Internal Flesh Breakdown of ‘Keitt’ Mango in a Steep Slope Mountain Area, Southwest China" Horticulturae 8, no. 6: 533. https://doi.org/10.3390/horticulturae8060533