
Physiological Study of the Efficacy of Archer® Eclipse in the Protection against Sunburn in Cucumber Plants

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
2.2. Description of the Treatments and Experimental Design
2.3. Plant Sampling
2.4. Leaf Temperature and Thermographic Images
2.5. Gas Exchange Measurements
2.6. Chl a Fluorescence Analysis
2.7. Concentration of Chls and Carotenoids
2.8. Electrolyte Leakage
2.9. Determination of the Concentration of Oxidative Indicators (Malondialdehyde (MDA), H2O2, and O2−)
2.10. Determination of Total Phenol Concentration
2.11. Statistical Analysis
3. Results
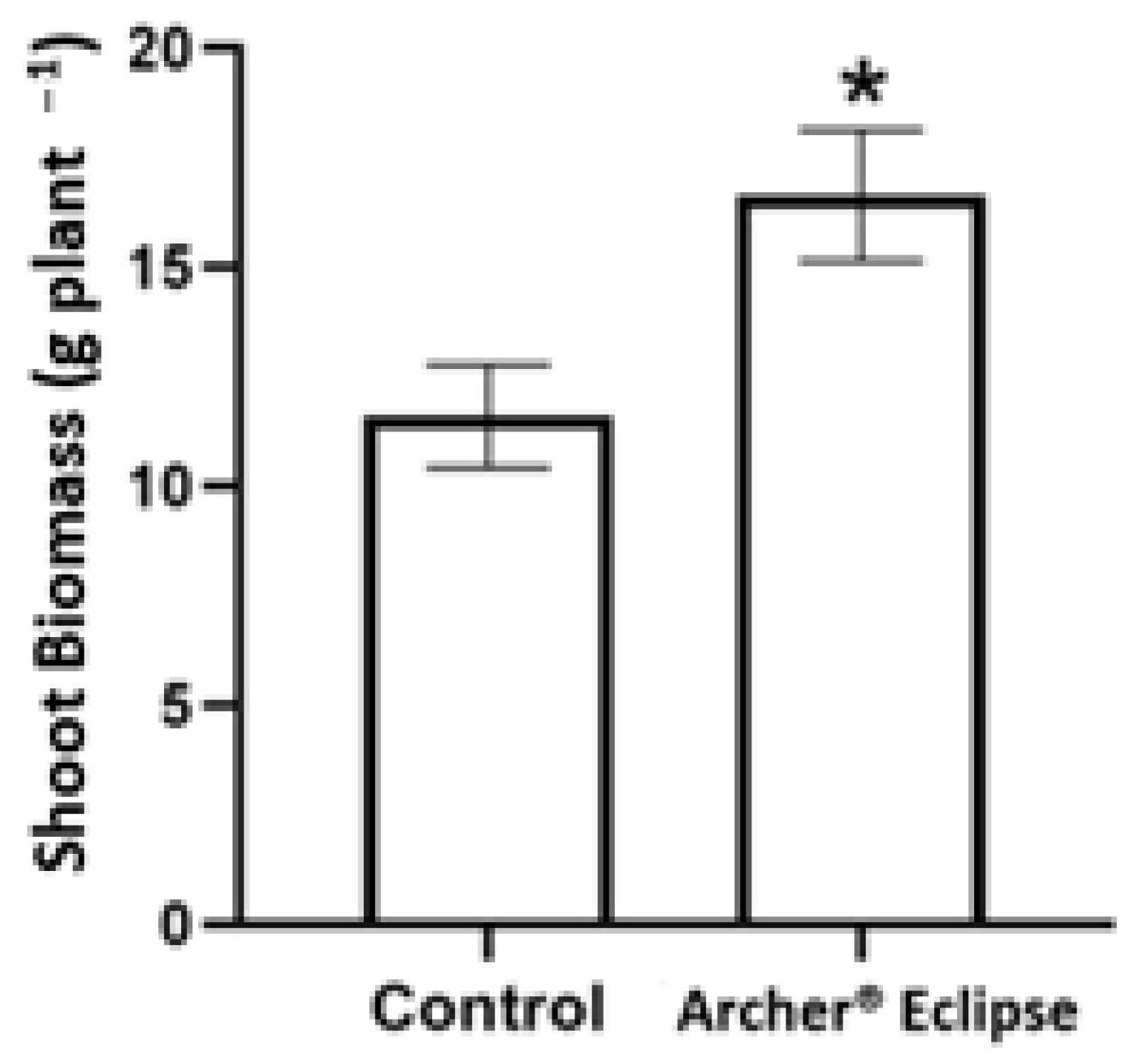
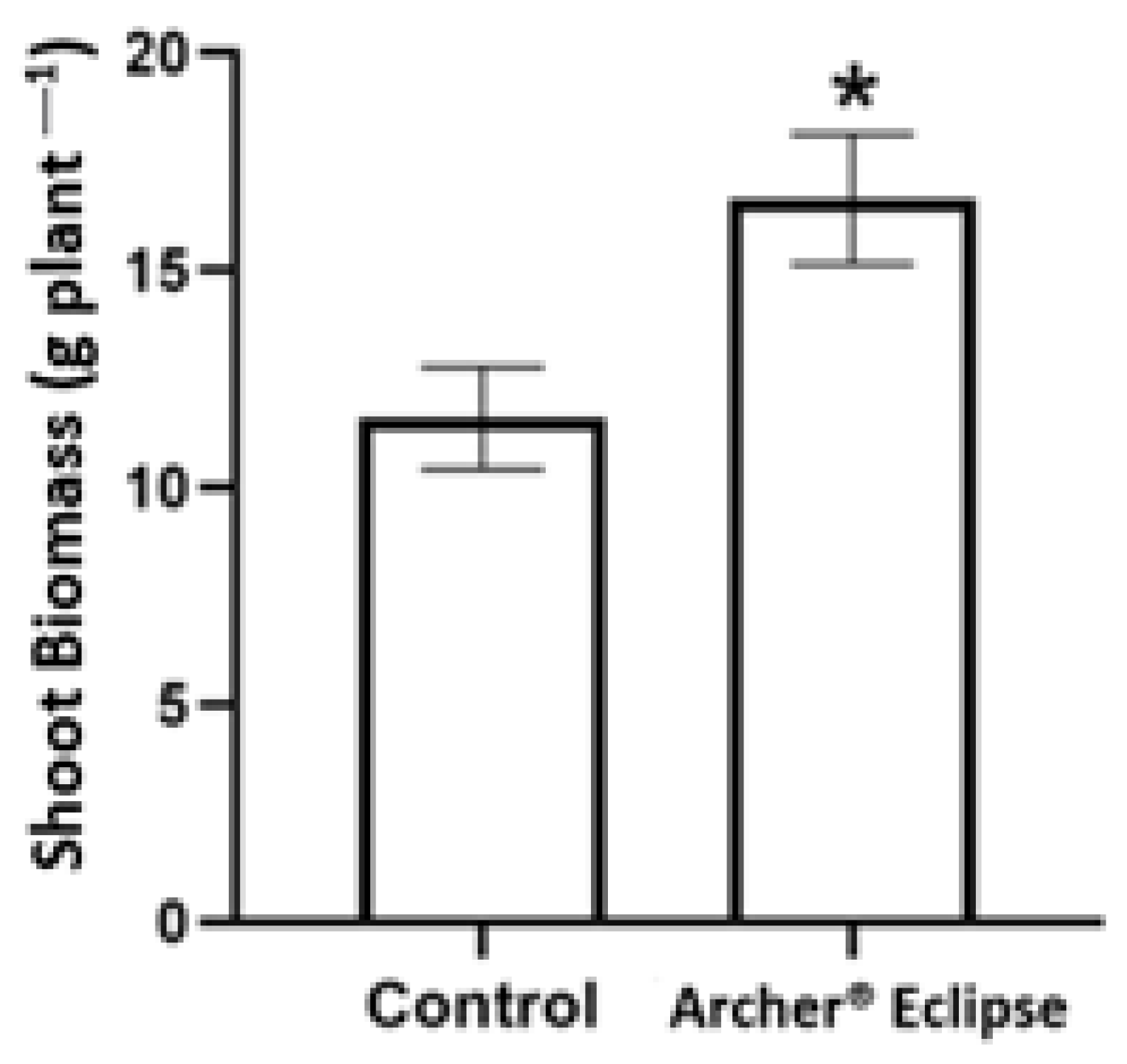
3.1. Visual Symptoms and Biomass
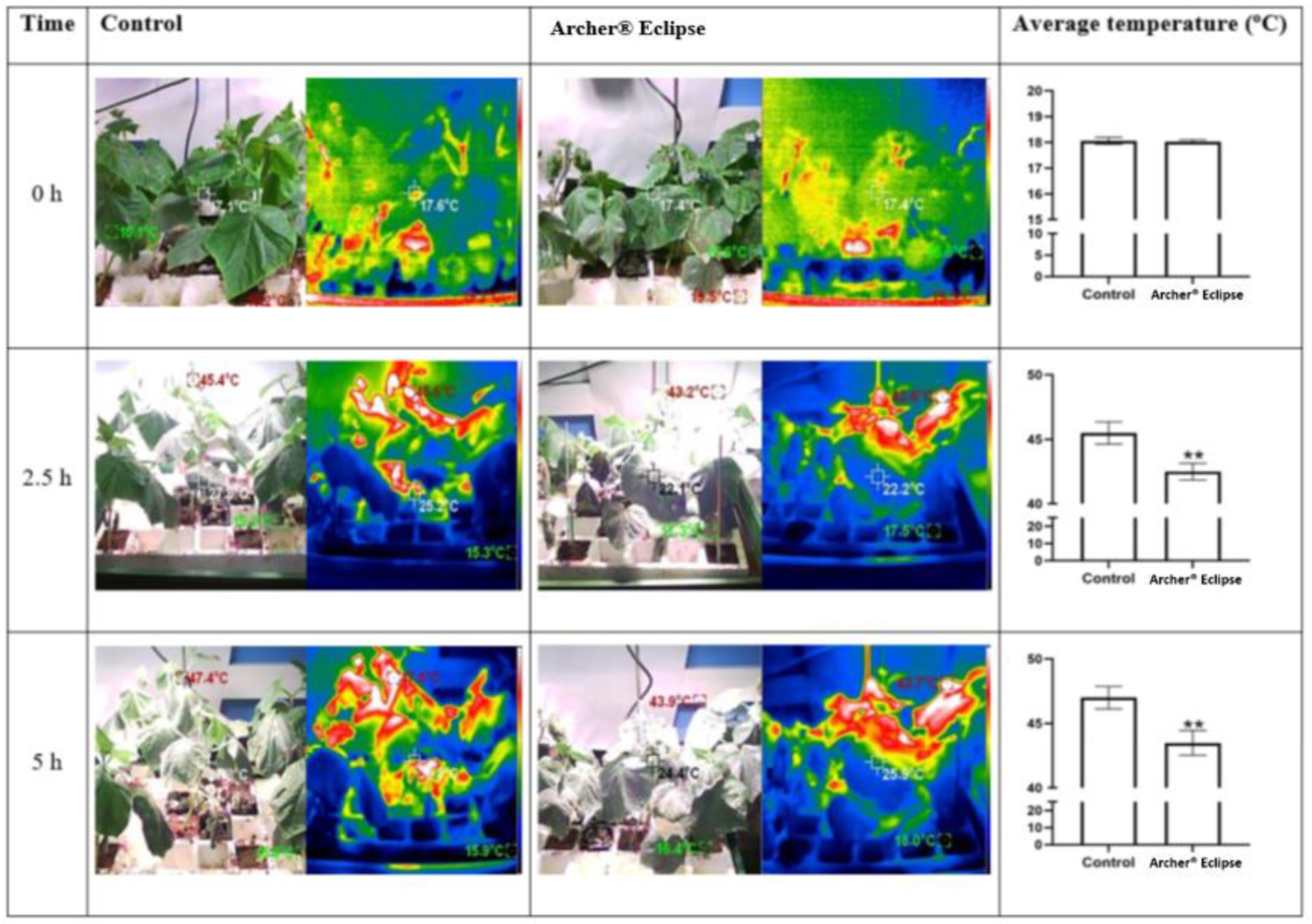
3.2. Leaf Temperature
3.3. Photosynthetic Parameters
3.4. Oxidative Stress Indicators and Total Phenol Concentration
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gambetta, J.M.; Holzapfel, B.P.; Stoll, M.; Friedel, M. Sunburn in Grapes: A Review. Front. Plant Sci. 2021, 11, 604691. [Google Scholar] [CrossRef]
- Ayangbenro, A.S.; Babalola, O.O. Reclamation of Arid and Semi-Arid Soils: The Role of Plant Growth-Promoting Archaea and Bacteria. Curr. Plant Biol. 2021, 25, 100173. [Google Scholar] [CrossRef]
- Dawood, M.F.A.; Tahjib-Ul-Arif, M.; Sohag, A.A.M.; Abdel Latef, A.A.H.; Ragaey, M.M. Mechanistic Insight of Allantoin in Protecting Tomato Plants Against Ultraviolet C Stress. Plants 2020, 10, 11. [Google Scholar] [CrossRef] [PubMed]
- Katano, K.; Honda, K.; Suzuki, N. Integration between ROS Regulatory Systems and Other Signals in the Regulation of Various Types of Heat Responses in Plants. IJMS 2018, 19, 3370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lal, N.; Sahu, N. Management Strategies of Sun Burn in Fruit Crops-A Review. Int. J. Curr. Microbiol. App. Sci. 2017, 6, 1126–1138. [Google Scholar] [CrossRef]
- Manja, K.; Aoun, M. The Use of Nets for Tree Fruit Crops and Their Impact on the Production: A Review. Sci. Hortic. 2019, 246, 110–122. [Google Scholar] [CrossRef]
- Liu, Z.; Jiao, X.; Zhu, C.; Katul, G.G.; Ma, J.; Guo, W. Micro-Climatic and Crop Responses to Micro-Sprinkler Irrigation. Agric. Water Manag. 2021, 243, 106498. [Google Scholar] [CrossRef]
- Han, S.G.; Selvakumar, G.; Yi, P.H.; Lee, S.E. Effect of Sprinkler, Surface Drip and Subsurface Drip Irrigation Methods on ‘Fuji’/M9 and ‘Fuji’/M26 Apple Orchards Growth, Soil Properties, and Water Consumption. Korean J. Soil. Sci. Fert. 2018, 51, 608–615. [Google Scholar] [CrossRef]
- Glenn, D.M. Particle Film Mechanisms of Action That Reduce the Effect of Environmental Stress in ‘Empire’ Apple. J. Am. Soc. Hortic. Sci. 2009, 134, 314–321. [Google Scholar] [CrossRef] [Green Version]
- Gharaghani, A.; Mohammadi Javarzari, A.; Vahdati, K. Kaolin Particle Film Alleviates Adverse Effects of Light and Heat Stresses and Improves Nut and Kernel Quality in Persian Walnut. Sci. Hortic. 2018, 239, 35–40. [Google Scholar] [CrossRef]
- Brito, C.; Dinis, L.-T.; Moutinho-Pereira, J.; Correia, C. Kaolin, an Emerging Tool to Alleviate the Effects of Abiotic Stresses on Crop Performance. Sci. Hortic. 2019, 250, 310–316. [Google Scholar] [CrossRef]
- Mahmoudian, M.; Rahemi, M.; Karimi, S.; Yazdani, N.; Tajdini, Z.; Sarikhani, S.; Vahdati, K. Role of Kaolin on Drought Tolerance and Nut Quality of Persian Walnut. J. Saudi Soc. Agric. Sci. 2021, 20, 409–416. [Google Scholar] [CrossRef]
- Chandel, A.K.; Khot, L.R.; Osroosh, Y.; Peters, T.R. Thermal-RGB Imager Derived in-Field Apple Surface Temperature Estimates for Sunburn Management. Agric. For. Meteorol. 2018, 253–254, 132–140. [Google Scholar] [CrossRef]
- Ko, S.-S.; Jhong, C.-M.; Lin, Y.-J.; Wei, C.-Y.; Lee, J.-Y.; Shih, M.-C. Blue Light Mediates Chloroplast Avoidance and Enhances Photoprotection of Vanilla Orchid. IJMS 2020, 21, 8022. [Google Scholar] [CrossRef] [PubMed]
- Padhi, B.; Chauhan, G.; Kandoi, D.; Stirbet, A.; Tripathy, B.C.; Govindjee, G. A Comparison of Chlorophyll Fluorescence Transient Measurements, Using Handy PEA and FluorPen Fluorometers. Photosynthetica 2021, 59, 399–408. [Google Scholar] [CrossRef]
- Andrianto, H.; Suhardi; Faizal, A. Detection of Chlorophyll Content Based on Spectral Properties at Leaf Level: A Meta-Analysis. In Proceedings of the 2018 International Conference on Information Technology Systems and Innovation (ICITSI), Bandung-Padang, Indonesia, 22–26 October 2018; pp. 364–369. [Google Scholar]
- Soloklui, A.A.G.; Ershadi, A.; Fallahi, E. Evaluation of Cold Hardiness in Seven Iranian Commercial Pomegranate (Punica Granatum L.) Cultivars. HortScience 2012, 47, 1821–1825. [Google Scholar] [CrossRef] [Green Version]
- Morales, M.; Munné-Bosch, S. Malondialdehyde: Facts and Artifacts. Plant Physiol. 2019, 180, 1246–1250. [Google Scholar] [CrossRef] [Green Version]
- Junglee, S.; Urban, L.; Sallanon, H.; Lopez-Lauri, F. Optimized Assay for Hydrogen Peroxide Determination in Plant Tissue Using Potassium Iodide. AJAC 2014, 5, 730–736. [Google Scholar] [CrossRef] [Green Version]
- Barrameda-Medina, Y.; Montesinos-Pereira, D.; Romero, L.; Blasco, B.; Ruiz, J.M. Role of GSH Homeostasis under Zn Toxicity in Plants with Different Zn Tolerance. Plant Sci. 2014, 227, 110–121. [Google Scholar] [CrossRef]
- Alam, M.K.; Rana, Z.H.; Islam, S.N.; Akhtaruzzaman, M. Total Phenolic Content and Antioxidant Activity of Methanolic Extract of Selected Wild Leafy Vegetables Grown in Bangladesh: A Cheapest Source of Antioxidants. Potr. S. J. F. Sci. 2019, 13, 287–293. [Google Scholar] [CrossRef] [Green Version]
- Del Amor, F.M.; Gálvez, A.; Romero, M.; Otalora, G.; López-Marín, J. Preliminary Study of the Behaviour of a Courgette Crop Grown under Photoselective Shade Nets. Acta Hortic. 2020, 341–345. [Google Scholar] [CrossRef]
- De la Torre-González, A.; Navarro-León, E.; Albacete, A.; Blasco, B.; Ruiz, J.M. Study of Phytohormone Profile and Oxidative Metabolism as Key Process to Identification of Salinity Response in Tomato Commercial Genotypes. J. Plant Physiol. 2017, 216, 164–173. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.R.; Datta, S.C.; Varghese, E. Effect of Surround WP®, a Kaolin-Based Particle Film on Sunburn, Fruit Cracking and Postharvest Quality of ‘Kandhari’ Pomegranates. Crop Prot. 2018, 114, 18–22. [Google Scholar] [CrossRef]
- Nyongesah, M.J.; Wang, Q.; Li, P. Effectiveness of Photochemical Reflectance Index to Trace Vertical and Seasonal Chlorophyll a/b Ratio in Haloxylon Ammodendron. Acta Physiol. Plant 2015, 37, 2. [Google Scholar] [CrossRef]
- Liu, Y.; Dong, S.; Wei, S.; Wang, W.; Miao, H.; Bo, K.; Gu, X.; Zhang, S. QTL Mapping of Heat Tolerance in Cucumber (Cucumis sativus L.) at Adult Stage. Plants 2021, 10, 324. [Google Scholar] [CrossRef] [PubMed]
- Takshak, S.; Agrawal, S.B. Exogenous Application of IAA Alleviates Effects of Supplemental Ultraviolet-B Radiation in the Medicinal Plant Withania Somnifera Dunal. Plant Biol. J. 2017, 19, 904–916. [Google Scholar] [CrossRef]
- Tossi, V.E.; Regalado, J.J.; Iannicelli, J.; Laino, L.E.; Burrieza, H.P.; Escandón, A.S.; Pitta-Álvarez, S.I. Beyond Arabidopsis: Differential UV-B Response Mediated by UVR8 in Diverse Species. Front. Plant Sci. 2019, 10, 780. [Google Scholar] [CrossRef]
- Sgroppo, S.C.; Sosa, C.A. Zapallo anco (Cucurbita moschata, D.) fresco cortado tratado con luz UV-C. FaCENA 2021, 25, 7–19. [Google Scholar]
- Khan, N.; Ali, S.; Zandi, P.; Mehmood, A.; Ullah, S.; Ikram, M.; Ismail, I.; Shahid, M.A.; Babar, M.A. Role of Sugars, Amino Acids and Organic Acids in Improving Plant Abiotic Stress Tolerance. Pak. J. Bot. 2020, 52, 355–363. [Google Scholar] [CrossRef]
- Talanova, V.V.; Topchieva, L.V.; Titov, A.F. Effect of Abscisic Acid on the Resistance of Cucumber Seedlings to Combined Exposure to High Temperature and Chloride. Biol. Bull. Russ. Acad. Sci. 2006, 33, 619–622. [Google Scholar] [CrossRef]
- Yu, B.; Ming, F.; Liang, Y.; Wang, Y.; Gan, Y.; Qiu, Z.; Yan, S.; Cao, B. Heat Stress Resistance Mechanisms of Two Cucumber Varieties from Different Regions. IJMS 2022, 23, 1817. [Google Scholar] [CrossRef] [PubMed]
- Badmus, U.O.; Ač, A.; Klem, K.; Urban, O.; Jansen, M.A.K. A Meta-Analysis of the Effects of UV Radiation on the Plant Carotenoid Pool. Plant Physiol. Biochem. 2022, 183, 36–45. [Google Scholar] [CrossRef] [PubMed]
- Köhler, H.; Contreras, R.A.; Pizarro, M.; Cortés-Antíquera, R.; Zúñiga, G.E. Antioxidant Responses Induced by UVB Radiation in Deschampsia Antarctica Desv. Front. Plant Sci. 2017, 8, 921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sahin, U.; Ekinci, M.; Ors, S.; Turan, M.; Yildiz, S.; Yildirim, E. Effects of Individual and Combined Effects of Salinity and Drought on Physiological, Nutritional and Biochemical Properties of Cabbage (Brassica Oleracea Var. Capitata). Sci. Hortic. 2018, 240, 196–204. [Google Scholar] [CrossRef]
- Tarkowski, Ł.P.; Signorelli, S.; Höfte, M. Γ-Aminobutyric Acid and Related Amino Acids in Plant Immune Responses: Emerging Mechanisms of Action. Plant Cell Environ. 2020, 43, 1103–1116. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gs (mol m−2 s−¹) | Eleaf (mmol m−2 s−¹) | Ci (µmol mol−¹) | ACO2 (µmol m−2 s−¹) | WUE (µmol mol−¹) | |
|---|---|---|---|---|---|
| Control | 0.016 ± 0.002 | 0.56 ± 0.07 | 330.16 ± 62.52 | 0.61 ± 0.12 | 1.09 ± 0.09 |
| Treatment | 0.010 ± 0.004 | 0.33 ± 0.11 | 312.99 ± 47.32 | 0.67 ± 0.17 | 2.05 ± 0.24 |
| p-value | * | * | NS | NS | ** |
| Fv/Fm | PIABS | RC/ABS | ΨEo | |
|---|---|---|---|---|
| Control | 0.34 ± 0.11 | 2.52 ± 1.20 | 0.38 ± 0.22 | 0.47 ± 0.08 |
| Treatment | 0.83 ± 0.01 | 10.23 ± 1.09 | 1.00 ± 0.04 | 0.67 ± 0.01 |
| p-value | *** | *** | *** | *** |
| Total Chls (mg g−1 FW) | Chl a/b Ratio (-) | Carotenoids (mg g−1 PF) | |
|---|---|---|---|
| Control | 0.373 ± 0.007 | 1.40 ± 0.03 | 0.169 ± 0.003 |
| Treatment | 0.454 ± 0.008 | 1.78 ± 0.01 | 0.159 ± 0.002 |
| p-value | *** | *** | ** |
| EC (%) | MDA (μM g−1 FW) | O2− (μg g−1 FW) | H2O2 (μg g−1 FW) | |
|---|---|---|---|---|
| Control | 31.59 ± 0.46 | 19.46 ± 0.92 | 31.44 ± 0.27 | 53.83 ± 11.11 |
| Treatment | 17.26 ± 0.57 | 5.61 ± 0.38 | 16.27 ± 0.44 | 10.97 ± 4.35 |
| p-value | *** | *** | *** | *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Navarro-Morillo, I.; Garcia-Sanchez, F.; Camara-Zapata, J.M.; Navarro-León, E.; Izquierdo-Ramos, M.J.; Blasco, B.; Ruiz, J.M. Physiological Study of the Efficacy of Archer® Eclipse in the Protection against Sunburn in Cucumber Plants. Horticulturae 2022, 8, 500. https://doi.org/10.3390/horticulturae8060500
Navarro-Morillo I, Garcia-Sanchez F, Camara-Zapata JM, Navarro-León E, Izquierdo-Ramos MJ, Blasco B, Ruiz JM. Physiological Study of the Efficacy of Archer® Eclipse in the Protection against Sunburn in Cucumber Plants. Horticulturae. 2022; 8(6):500. https://doi.org/10.3390/horticulturae8060500
Chicago/Turabian StyleNavarro-Morillo, Iván, Francisco Garcia-Sanchez, Jose Maria Camara-Zapata, Eloy Navarro-León, María José Izquierdo-Ramos, Begoña Blasco, and Juan Manuel Ruiz. 2022. "Physiological Study of the Efficacy of Archer® Eclipse in the Protection against Sunburn in Cucumber Plants" Horticulturae 8, no. 6: 500. https://doi.org/10.3390/horticulturae8060500