
Effects of Chromium Toxicity on Physiological Performance and Nutrient Uptake in Two Grapevine Cultivars (Vitis vinifera L.) Growing on Own Roots or Grafted onto Different Rootstocks

 ,
,  ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Experimental Conditions
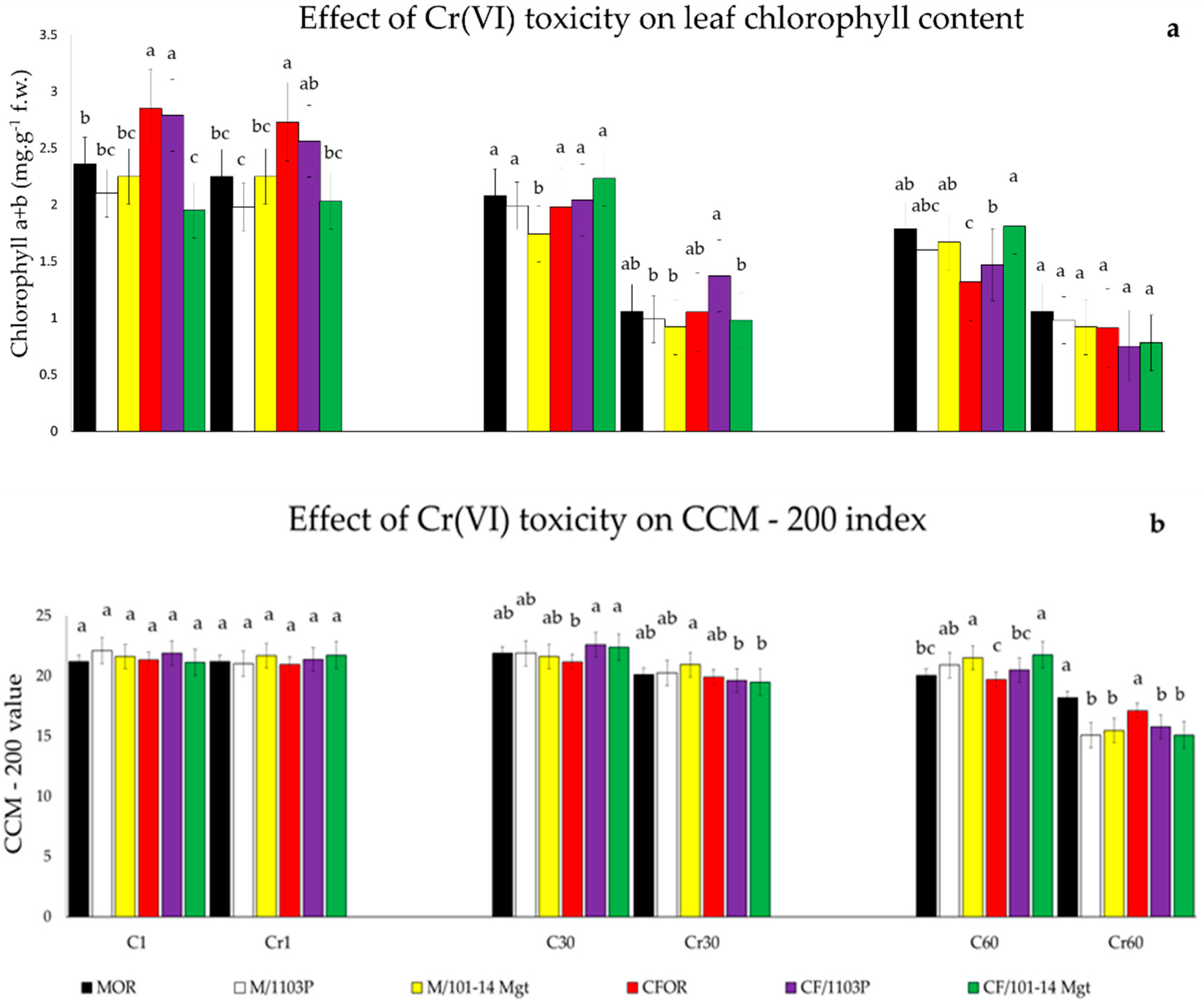
2.2. Chlorophyll Content
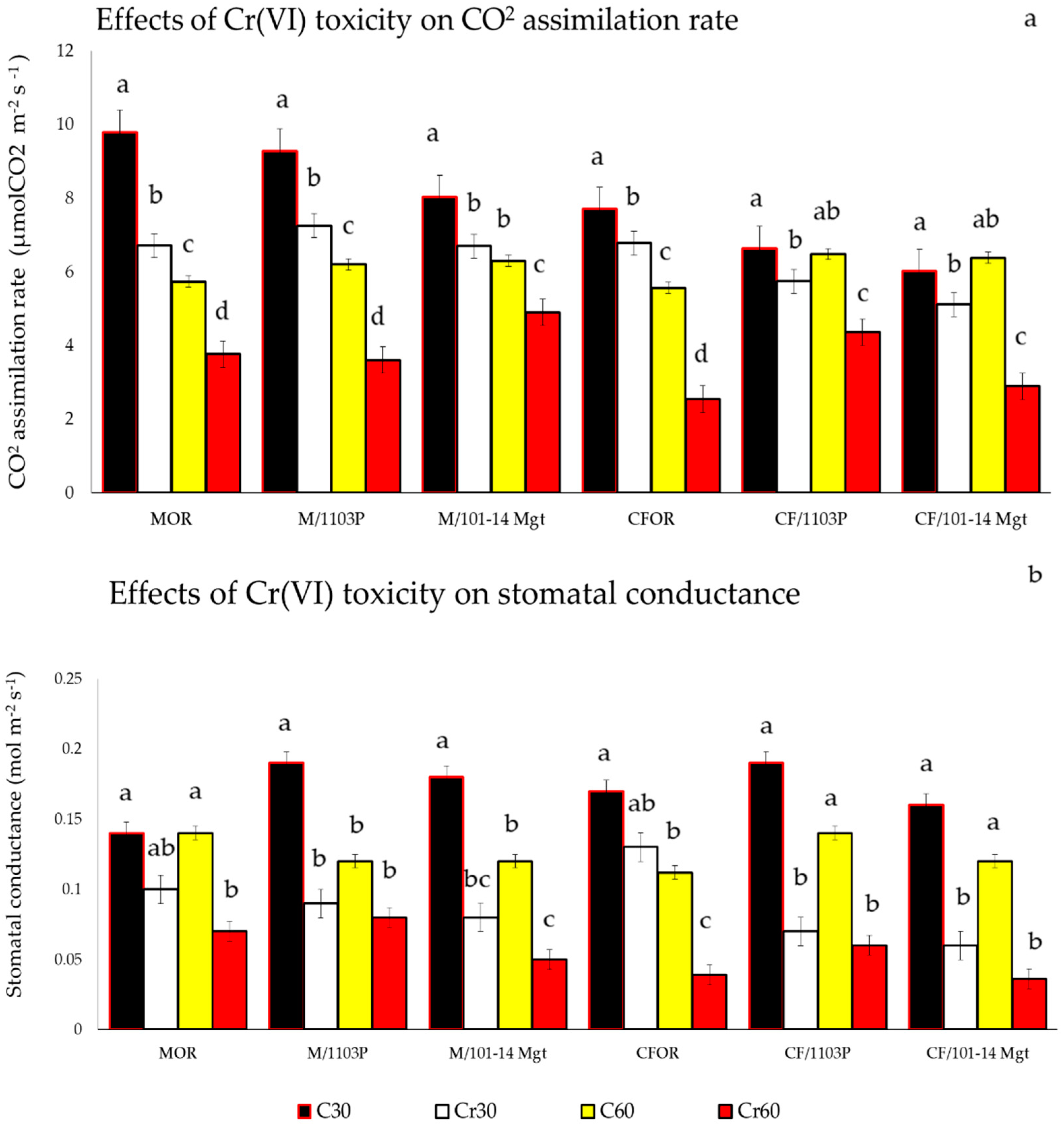
2.3. Stem Water Potential and Gas Exchange
2.4. Tissue Nutrient Concentrations and Growth Parameters
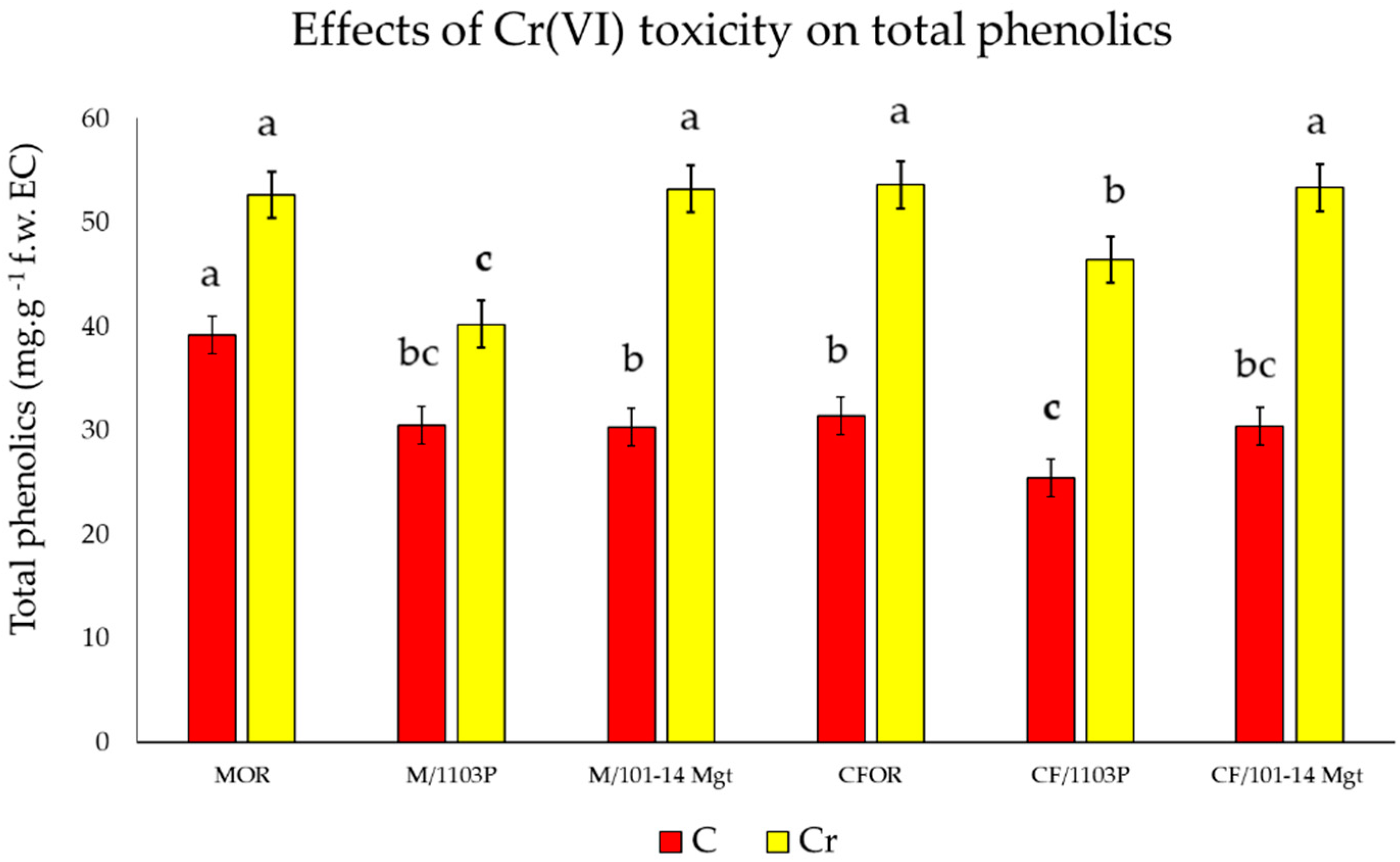
2.5. Total Phenolics
2.6. Statistical Analysis
3. Results
3.1. Plant Growth Parameters
3.2. Tissue Nutrient Concentrations
3.3. Chlorophyll Content
3.4. Total Phenolic Content
3.5. Water Status and Photosynthetic Activity
4. Discussion
4.1. Nutrient Concentrations in Plant Tissues and Vine Growth
4.2. Chlorophyll Pigments, Water Status, and Photosynthetic Activity
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sangwan, P.; Kumar, V.; Joshi, U.N. Effect of Chromium(VI) Toxicity on Enzymes of Nitrogen Metabolism in Clusterbean (Cyamopsis tetragonoloba L.). Enzym. Res. 2014, 2014, 784036. [Google Scholar] [CrossRef] [Green Version]
- Linos, A.; Petralias, A.; Christophi, C.A.; Christoforidou, E.; Kouroutou, P.; Stoltidis, M.; Veloudaki, A.; Tzala, E.; Makris, K.C.; Karagas, M.R. Oral Ingestion of Hexavalent Chromium through Drinking Water and Cancer Mortality in an Industrial Area of Greece-An Ecological Study. Environ. Health 2011, 10, 50. [Google Scholar] [CrossRef] [Green Version]
- Shanker, A.K.; Cervantes, C.; Loza-Tavera, H.; Avudainayagam, S. Chromium Toxicity in Plants. Environ. Int. 2005, 31, 739–753. [Google Scholar] [CrossRef]
- Amin, H.; Arain, B.A.; Amin, F.; Surhio, M.A. Phytotoxicity of Chromium on Germination, Growth and Biochemical Attributes of Hibiscus esculentus L. AJPS 2013, 4, 2431–2439. [Google Scholar] [CrossRef] [Green Version]
- Nagajyoti, P.C.; Lee, K.D.; Sreekanth, T.V.M. Heavy Metals, Occurrence and Toxicity for Plants: A Review. Environ. Chem. Lett. 2010, 8, 199–216. [Google Scholar] [CrossRef]
- Chung, J.-B.; Burau, R.G.; Zasoski, R.J. Chromate Generation by Chromate depleted Subsurface Materials. Water Air Soil Pollut. 2001, 128, 407–417. [Google Scholar] [CrossRef]
- Fantoni, D.; Brozzo, G.; Canepa, M.; Cipolli, F.; Marini, L.; Ottonello, G.; Zuccolini, M. Natural Hexavalent Chromium in Groundwaters Interacting with Ophiolitic Rocks. Environ. Geol. 2002, 42, 871–882. [Google Scholar] [CrossRef]
- Morrison, J.M.; Goldhaber, M.B.; Lee, L.; Holloway, J.M.; Wanty, R.B.; Wolf, R.E.; Ranville, J.F. A Regional-Scale Study of Chromium and Nickel in Soils of Northern California, USA. Appl. Geochem. 2009, 24, 1500–1511. [Google Scholar] [CrossRef]
- Zayed, A.M.; Terry, N. Chromium in the Environment: Factors Affecting Biological Remediation. Plant Soil 2003, 249, 139–156. [Google Scholar] [CrossRef]
- Gupta, V.K.; Rastogi, A.; Nayak, A. Adsorption Studies on the Removal of Hexavalent Chromium from Aqueous Solution using a low cost Fertilizer Industry Waste Material. J Colloid Interface Sci. 2010, 342, 135–141. [Google Scholar] [CrossRef] [Green Version]
- Vasilatos, C.; Megremi, I.; Economou-Eliopoulos, M.; Mitsis, I. Hexavalent Chromium and Other Toxic Elements in Natural Waters in the Thiva–Tanagra–Malakasa Basin, Greece. Hell. J. Geosci. 2012, 43, 57–66. [Google Scholar]
- Economou-Eliopoulos, M.; Antivachi, D.; Vasilatos, C.; Megremi, I. Evaluation of the Cr (VI) and Other Toxic Element Contamination and Their Potential Sources: The Case of the Thiva Basin (Greece). Geosci. Front. 2012, 3, 523–539. [Google Scholar] [CrossRef] [Green Version]
- Kaprara, E.; Kazakis, N.; Simeonidis, K.; Coles, S.; Zouboulis, A.I.; Samaras, P.; Mitrakas, M. Occurrence of Cr (VI) in Drinking Water of Greece and Relation to the Geological Background. J. Hazard. Mater. 2015, 281, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Licina, V.; Antic-Mladenovic, S.; Kresovic, M.; Rinklebe, J. Effect of High Nickel and Chromium Background Levels in Serpentine Soil on Their Accumulation in Organs of a Perennial Plant. Commun. Soil Sci. Plant Anal. 2010, 41, 482–496. [Google Scholar] [CrossRef]
- Gąstol, M.; Domagała-Świątkiewicz, I. Trace Element Partitioning in ‘Sibera’ Grapevines as Affected by Nitrogen Fertilisation. SAJEV 2016, 35, 217–225. [Google Scholar] [CrossRef]
- Sinha, S.; Singh, S.; Mallick, S.; Sinam, G. Role of Antioxidants in Cr Tolerance of Three Crop Plants: Metal Accumulation in Seeds. Ecotoxicol. Environ. Saf. 2009, 72, 1111–1121. [Google Scholar] [CrossRef]
- Shahid, M.; Shamshad, S.; Rafiq, M.; Khalid, S.; Bibi, I.; Niazi, N.K.; Dumat, C.; Rashid, M.I. Chromium Speciation, Bioavailability, Uptake, Toxicity and Detoxification in Soil-Plant System: A Review. Chemosphere 2017, 178, 513–533. [Google Scholar] [CrossRef]
- Hoagland, D.R.; Arnon, D.I. The Water-Culture Method for Growing Plants without Soil. Circ. Calif. Agric. Exp. Stn. 1950, 347, 32. [Google Scholar]
- Wintermans, J.F.G.M.; De Mots, A. Spectrophotometric Characteristics of Chlorophylls a and b and Their Phenophytins in Ethanol. Biochim. Biophys. Acta (BBA)-Biophys. Incl. Photosynth. 1965, 109, 448–453. [Google Scholar] [CrossRef]
- Choné, X. Stem Water Potential Is a Sensitive Indicator of Grapevine Water Status. Ann. Bot. 2001, 87, 477–483. [Google Scholar] [CrossRef] [Green Version]
- Chyla, M.A.; Zyrnicki, W. Determination of Metal Concentrations in Animal Hair by the ICP Method. BTER 2000, 75, 187–194. [Google Scholar] [CrossRef]
- Chapman, H.D.; Pratt, P.F. Methods of Analysis for Soils, Plants and Waters; Division of Agricultural Sciences, University of California: Riverside, CA, USA, 1961; p. 309. [Google Scholar]
- Gaines, T.P.; Mitchell, G.A. Boron Determination in Plant Tissue by the Azomethine H Method. Commun. Soil Sci. Plant Anal. 1979, 10, 1099–1108. [Google Scholar] [CrossRef]
- Di Stefano, R.; Cravero, M.C.; Gentilini, M. Methodi per lo studio dei polifenoli dei vini. L’Enotecnico Maggio 1989, 19, 83–89. [Google Scholar]
- Shanker, A.; Djanaguiraman, M.; Sudhagar, R.; Chandrashekar, C.; Pathmanabhan, G. Differential Antioxidative Response of Ascorbate Glutathione Pathway Enzymes and Metabolites to Chromium Speciation Stress in Green Gram ((Vigna radiata L.) R. Wilczek. Cv CO4) Roots. Plant Sci. 2004, 166, 1035–1043. [Google Scholar] [CrossRef]
- Mangabeira, P.A.; Ferreira, A.S.; de Almeida, A.-A.F.; Fernandes, V.F.; Lucena, E.; Souza, V.L.; dos Santos Júnior, A.J.; Oliveira, A.H.; Grenier-Loustalot, M.F.; Barbier, F.; et al. Compartmentalization and Ultrastructural Alterations Induced by Chromium in Aquatic Macrophytes. Biometals 2011, 24, 1017–1026. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.F.; Wang, Z.W.; Dong, F.; Lei, G.J.; Shi, Y.Z.; Li, G.X.; Zheng, S.J. Exogenous Auxin Alleviates Cadmium Toxicity in Arabidopsis Thaliana by Stimulating Synthesis of Hemicellulose 1 and Increasing the Cadmium Fixation Capacity of Root Cell Walls. J. Hazard. Mater. 2013, 263, 398–403. [Google Scholar] [CrossRef] [PubMed]
- Caldelas, C.; Bort, J.; Febrero, A. Ultrastructure and Subcellular Distribution of Cr in Iris pseudacorus L. Using TEM and X-Ray Microanalysis. Cell Biol. Toxicol. 2012, 28, 57–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Do Nascimento, J.L.; de Almeida, A.-A.F.; Barroso, J.P.; Mangabeira, P.A.O.; Ahnert, D.; Sousa, A.G.R.; Silva, J.V.S.; Baligar, V.C. Physiological, Ultrastructural, Biochemical and Molecular Responses of Young Cocoa Plants to the Toxicity of Cr (III) in Soil. Ecotoxicol. Environ. Saf. 2018, 159, 272–283. [Google Scholar] [CrossRef]
- Santana, K.B.; de Almeida, A.-A.F.; Souza, V.L.; Mangabeira, P.A.O.; da Silva, D.C.; Gomes, F.P.; Dutruch, L.; Loguercio, L.L. Physiological Analyses of Genipa Americana L. Reveals a Tree with Ability as Phytostabilizer and Rhizofilterer of Chromium Ions for Phytoremediation of Polluted Watersheds. Environ. Exp. Bot. 2012, 80, 35–42. [Google Scholar] [CrossRef]
- Sharma, R.K.; Agrawal, M.; Marshall, F. Heavy Metal Contamination in Vegetables Grown in Wastewater Irrigated Areas of Varanasi, India. Bull. Environ. Contam. Toxicol. 2006, 77, 312–318. [Google Scholar] [CrossRef]
- Gardea-Torresdey, J.L.; Peralta-Videa, J.R.; Montes, M.; de la Rosa, G.; Corral-Diaz, B. Bioaccumulation of Cadmium, Chromium and Copper by Convolvulus Arvensis L.: Impact on Plant Growth and Uptake of Nutritional Elements. Bioresour. Technol. 2004, 92, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Handa, N.; Kohli, S.K.; Thukral, A.K.; Bhardwaj, R.; Alyemeni, M.N.; Wijaya, L.; Ahmad, P. Protective Role of Selenium against Chromium Stress Involving Metabolites and Essential Elements in Brassica juncea L. Seedlings. 3 Biotech 2018, 8, 66. [Google Scholar] [CrossRef]
- Oliveira, H. Chromium as an Environmental Pollutant: Insights on Induced Plant Toxicity. J. Bot. 2012, 2012, 375843. [Google Scholar] [CrossRef]
- Kabata-Pendias, A. Trace Elements in Soils and Plants, 3rd ed.; CRC Press: Boca Raton, FL, USA, 2000. [Google Scholar] [CrossRef]
- Barbosa, R.; Dealmeida, A.; Mielke, M.; Loguercio, L.; Mangabeira, P.; Gomes, F. A Physiological Analysis of Genipa americana L.: A Potential Phytoremediator Tree for Chromium Polluted Watersheds. Environ. Exp. Bot. 2007, 61, 264–271. [Google Scholar] [CrossRef]
- Papenbrock, J.; Mock, H.-P.; Tanaka, R.; Kruse, E.; Grimm, B. Role of Magnesium Chelatase Activity in the Early Steps of the Tetrapyrrole Biosynthetic Pathway. Plant Physiol. 2000, 122, 1161–1170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganesh, K.S.; Baskaran, L.; Rajasekaran, S.; Sumathi, K.; Chidambaram, A.L.A.; Sundaramoorthy, P. Chromium Stress Induced Alterations in Biochemical and Enzyme Metabolism in Aquatic and Terrestrial Plants. Colloids Surf. B Biointerfaces 2008, 63, 159–163. [Google Scholar] [CrossRef]
- Ilag, L.L.; Kumar, A.M.; Söll, D. Light Regulation of Chlorophyll Biosynthesis at the Level of 5-Aminolevulinate Formation in Arabidopsis. Plant Cell 1994, 6, 265–275. [Google Scholar] [CrossRef] [Green Version]
- Islam, F.; Yasmeen, T.; Arif, M.S.; Riaz, M.; Shahzad, S.M.; Imran, Q.; Ali, I. Combined Ability of Chromium (Cr) Tolerant Plant Growth Promoting Bacteria (PGPB) and Salicylic Acid (SA) in Attenuation of Chromium Stress in Maize Plants. Plant Physiol. Biochem. 2016, 108, 456–467. [Google Scholar] [CrossRef]
- Qureshi, F.F.; Ashraf, M.A.; Rasheed, R.; Ali, S.; Hussain, I.; Ahmed, A.; Iqbal, M. Organic Chelates Decrease Phytotoxic Effects and Enhance Chromium Uptake by Regulating Chromium-Speciation in Castor Bean (Ricinus communis L.). Sci. Total Environ. 2020, 716, 137061. [Google Scholar] [CrossRef]
- Wong, P.K.; Chang, L. Effects of Copper, Chromium and Nickel on Growth, Photosynthesis and Chlorophyll a Synthesis of Chlorella Pyrenoidosa 251. Environ. Pollut. 1991, 72, 127–139. [Google Scholar] [CrossRef]
- Ouzounidou, G.; Moustakas, M.; Eleftheriou, E.P. Physiological and Ultrastructural Effects of Cadmium on Wheat (Triticum aestivum L.) Leaves. Arch. Environ. Contam. Toxicol. 1997, 32, 154–160. [Google Scholar] [CrossRef] [PubMed]
- Sundaramoorthy, P.; Chidambaram, A.; Ganesh, K.S.; Unnikannan, P.; Baskaran, L. Chromium Stress in Paddy: (I) Nutrient Status of Paddy under Chromium Stress; (Ii) Phytoremediation of Chromium by Aquatic and Terrestrial Weeds. Comptes Rendus Biol. 2010, 333, 597–607. [Google Scholar] [CrossRef] [PubMed]
- Marschner, H.; Marschner, P. Mineral Nutrition of Higher Plants; Gulf Professional Publishing: London, UK; Academic Press: San Diego, CA, USA, 1995. [Google Scholar]
- Janas, K.M.; Zielińska-Tomaszewska, J.; Rybaczek, D.; Maszewski, J.; Posmyk, M.M.; Amarowicz, R.; Kosińska, A. The Impact of Copper Ions on Growth, Lipid Peroxidation, and Phenolic Compound Accumulation and Localization in Lentil (Lens culinaris Medic.) Seedlings. J. Plant Physiol. 2010, 167, 270–276. [Google Scholar] [CrossRef]
- Singh, H.P.; Mahajan, P.; Kaur, S.; Batish, D.R.; Kohli, R.K. Chromium Toxicity and Tolerance in Plants. Environ. Chem. Lett. 2013, 11, 229–254. [Google Scholar] [CrossRef]
- Mallick, S.; Sinam, G.; Kumar Mishra, R.; Sinha, S. Interactive Effects of Cr and Fe Treatments on Plants Growth, Nutrition and Oxidative Status in Zea mays L. Ecotoxicol. Environ. Saf. 2010, 73, 987–995. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Jiang, W.; Li, M. Effects of Trivalent and Hexavalent Chromium on Root Growth and Cell Division of Allium cepa. Hereditas 2008, 117, 23–29. [Google Scholar] [CrossRef]
- Bagchi, D.; Stohs, S.J.; Downs, B.W.; Bagchi, M.; Preuss, H.G. Cytotoxicity and Oxidative Mechanisms of Different Forms of Chromium. Toxicology 2002, 180, 5–22. [Google Scholar] [CrossRef]
- Gill, R.A.; Zang, L.; Ali, B.; Farooq, M.A.; Cui, P.; Yang, S.; Ali, S.; Zhou, W. Chromium-Induced Physio-Chemical and Ultrastructural Changes in Four Cultivars of Brassica napus L. Chemosphere 2015, 120, 154–164. [Google Scholar] [CrossRef]
- Afshan, S.; Ali, S.; Bharwana, S.A.; Rizwan, M.; Farid, M.; Abbas, F.; Ibrahim, M.; Mehmood, M.A.; Abbasi, G.H. Citric Acid Enhances the Phytoextraction of Chromium, Plant Growth, and Photosynthesis by Alleviating the Oxidative Damages in Brassica napus L. Environ. Sci Pollut Res 2015, 22, 11679–11689. [Google Scholar] [CrossRef]
- Mohanty, M.; Patra, H. Tolerance Potential and Physiological Responses of Helianthus annuus L. Exposed to Varying Doses of Hexavalent Chromium. J. Mater. Environ. Sci. 2016, 7, 2221–2228. [Google Scholar]
- Sytar, O.; Kumar, A.; Latowski, D.; Kuczynska, P.; Strzałka, K.; Prasad, M.N.V. Heavy Metal-Induced Oxidative Damage, Defense Reactions, and Detoxification Mechanisms in Plants. Acta Physiol. Plant. 2013, 35, 985–999. [Google Scholar] [CrossRef]
- Kiani-Pouya, A.; Rasouli, F. The Potential of Leaf Chlorophyll Content to Screen Bread-Wheat Genotypes in Saline Condition. Photosynthetica 2014, 52, 288–300. [Google Scholar] [CrossRef]
- Chandra, P.; Kulshreshtha, K. Chromium Accumulation and Toxicity in Aquatic Vascular Plants. Bot. Rev. 2004, 70, 313–327. [Google Scholar] [CrossRef]
- Panda, S.K.; Choudhury, S. Chromium Stress in Plants. Braz. J. Plant Physiol. 2005, 17, 95–102. [Google Scholar] [CrossRef]
- Sinha, S.; Saxena, R.; Singh, S. Chromium Induced Lipid Peroxidation in the Plants of Pistia Stratiotes L.: Role of Antioxidants and Antioxidant Enzymes. Chemosphere 2005, 58, 595–604. [Google Scholar] [CrossRef]
- Balal, R.M.; Shahid, M.A.; Vincent, C.; Zotarelli, L.; Liu, G.; Mattson, N.S.; Rathinasabapathi, B.; Martínez-Nicolas, J.J.; Garcia-Sanchez, F. Kinnow Mandarin Plants Grafted on Tetraploid Rootstocks Are More Tolerant to Cr-Toxicity than Those Grafted on Its Diploids One. Environ. Exp. Bot. 2017, 140, 8–18. [Google Scholar] [CrossRef]
- Jaffe, E.K.; Ali, S.; Mitchell, L.W.; Taylor, K.M.; Volin, M.; Markham, G.D. Characterization of the Role of the Stimulatory Magnesium of Escherichia Coli Porphobilinogen Synthase. Biochemistry 1995, 34, 244–251. [Google Scholar] [CrossRef]
- Buchanan, B.B.; Gruissem, W.; Jones, R.L. Biochemistry and Molecular Biology of Plants; John Wiley & Sons: Hoboken, NJ, USA, 2015. [Google Scholar]
- Breinig, S.; Kervinen, J.; Stith, L.; Wasson, A.S.; Fairman, R.; Wlodawer, A.; Zdanov, A.; Jaffe, E.K. Control of Tetrapyrrole Biosynthesis by Alternate Quaternary Forms of Porphobilinogen Synthase. Nat. Struct. Mol. Biol. 2003, 10, 757–763. [Google Scholar] [CrossRef]
- Pandey, N.; Sharma, C.P. Chromium Interference in Iron Nutrition and Water Relations of Cabbage. Environ. Exp. Bot. 2003, 49, 195–200. [Google Scholar] [CrossRef]
- Hayat, S.; Khalique, G.; Irfan, M.; Wani, A.S.; Tripathi, B.N.; Ahmad, A. Physiological Changes Induced by Chromium Stress in Plants: An Overview. Protoplasma 2012, 249, 599–611. [Google Scholar] [CrossRef]
- Ali, S.; Farooq, M.A.; Yasmeen, T.; Hussain, S.; Arif, M.S.; Abbas, F.; Bharwana, S.A.; Zhang, G. The Influence of Silicon on Barley Growth, Photosynthesis and Ultra-Structure under Chromium Stress. Ecotoxicol. Environ. Saf. 2013, 89, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Assche, F.; Clijsters, H. Effects of Metals on Enzyme Activity in Plants. Plant Cell Environ. 1990, 13, 195–206. [Google Scholar] [CrossRef]
- Davies, F.T.; Puryear, J.D.; Newton, R.J.; Egilla, J.N.; Saraiva Grossi, J.A. Mycorrhizal fungi increase chromium uptake by sunflower plants: Influence on tissue mineral concentration, growth, and gas exchange. J. Plant Nutr. 2002, 25, 2389–2407. [Google Scholar] [CrossRef] [Green Version]
- Dan, T.V.; KrishnaRaj, S.; Saxena, P.K. Metal Tolerance of Scented Geranium (Pelargonium Sp. ‘Frensham’): Effects of Cadmium and Nickel on Chlorophyll Fluorescence Kinetics. Int. J. Phytoremediation 2000, 2, 91–104. [Google Scholar] [CrossRef]
- Subrahmanyam, D. Effects of Chromium Toxicity on Leaf Photosynthetic Characteristics and Oxidative Changes in Wheat (Triticum aestivum L.). Photosynthetica 2008, 46, 339. [Google Scholar] [CrossRef]
- DalCorso, G.; Fasani, E.; Furini, A. Recent Advances in the Analysis of Metal Hyperaccumulation and Hypertolerance in Plants Using Proteomics. Front. Plant Sci. 2013, 4, 280. [Google Scholar] [CrossRef] [Green Version]
- You, J.; Chan, Z. ROS Regulation During Abiotic Stress Responses in Crop Plants. Front. Plant Sci. 2015, 6, 1092. [Google Scholar] [CrossRef] [Green Version]
- Toor, R.K.; Savage, G.P. Changes in Major Antioxidant Components of Tomatoes during Post-Harvest Storage. Food Chem. 2006, 99, 724–727. [Google Scholar] [CrossRef]
- Kısa, D.; Elmastaş, M.; Öztürk, L.; Kayır, Ö. Responses of the Phenolic Compounds of Zea Mays under Heavy Metal Stress. Appl. Biol. Chem. 2016, 59, 813–820. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Merlot | Cabernet Franc | ||||||
|---|---|---|---|---|---|---|---|
| [g] | Shoots | Trunks | Roots | Shoots | Trunks | Roots | |
| Control | Own roots | 49.78 a | 62.33 a | 52.83 b | 42.34 ab | 54.61 c | 44.54 b |
| 1103P | 50.66 a | 59.09 a | 62.43 a | 43.12 a | 71.53 a | 51.32 a | |
| 101-14 Mgt | 43.63 bc | 45.11 b | 51.16 b | 38.78 bc | 62.35 b | 42.82 b | |
| 120 μM Cr(VI) | Own roots | 46.16 b | 61.61 a | 44.87 c | 39.56 ab | 55.16 c | 37.16 c |
| 1103P | 43.13 bc | 58.86 a | 32.37 d | 38.67 bc | 72.36 a | 41.34 b | |
| 101-14 Mgt | 41.66 c | 46.50 b | 31.67 d | 35.33 c | 60.94 bc | 31.53 d | |
| F: | 5.281 | 11.665 | 15.439 | 5.281 | 11.665 | 15.439 | |
| Leaves | |||||||||||
| Merlot | Cabernet Franc | ||||||||||
| [% d.w.] | N | P | K | Ca | Mg | N | P | K | Ca | Mg | |
| Control | Own roots | 2.13 a | 0.31 a | 2.12 a | 3.07 a | 0.57 a | 2.56 b | 0.30 abc | 2.31 a | 3.16 a | 0.55 a |
| 1103P | 2.09 a | 0.30 a | 1.47 cd | 2.58 b | 0.46 ab | 2.95 a | 0.35 a | 1.46 b | 2.98 ab 2.98 ab | 0.47 abc | |
| 101-14 Mgt | 1.79 b | 0.29 a | 1.83 b | 2.62 b | 0.52 ab | 1.81 cd | 0.32 ab | 2.03 a | 2.89 bc | 0.54 ab | |
| 120 μM Cr(VI) | Own roots | 1.79 b 1. | 0.21 b | 0.75 e | 2.74 b | 0.39 bc | 1.58 d | 0.29 bc | 1.30 b | 2.73 cd | 0.41 bc |
| 1103P | 1.80 b | 0.25 ab | 1.40 d | 2.36 c | 0.41 bc | 1.92 c | 0.27 bc | 1.21 b | 2.62 d | 0.38 c | |
| 101-14 Mgt | 1.81 ab | 0.20 b | 1.59 c | 2.62 b | 0.32 c | 1.73 cd | 0.26 c | 1.42 b | 2.67 d | 0.41 bc | |
| F | 8.344 | 6.502 | 44.952 | 9.699 | 6.149 | 8.344 | 6.502 | 44.952 | 9.699 | 6.194 | |
| Shoots | |||||||||||
| Merlot | Cabernet Franc | ||||||||||
| N | P | K | Ca | Mg | N | P | K | Ca | Mg | ||
| Control | Own roots | 0.32 e | 0.20 bc | 0.65 a | 0.78 a | 0.17 b | 0.87 a | 0.24 ab | 0.61 a | 0.46 c | 0.16 b |
| 1103P | 0.71 bc | 0.24 ab | 0.63 ab | 0.79 a | 0.15 c | 0.85 ab | 0.29 a | 0.67 a | 0.77 a | 0.17 b | |
| 101-14 Mgt | 0.76 b | 0.30 a | 0.58 ab | 0.75 ab | 0.65 a | 0.84 abc | 0.23 abc | 0.63 a | 0.73 a | 0.26 a | |
| 120 μM Cr(VI) | Own roots | 0.87 a | 0.18 bc | 0.38 ab | 0.68 bc | 0.14 c | 0.77 c | 0.16 cd | 0.42 ab | 0.57 b | 0.13 c |
| 1103P | 0.68 cd | 0.19 bc | 0.36 b | 0.69 bc | 0.15 c | 0.79 bc | 0.21 bc | 0.25 b | 0.59 b | 0.14 c | |
| 101-14 Mgt | 0.62 d | 0.16 c | 0.43 ab | 0.65 c | 0.14 c | 0.64 d | 0.12 d | 0.29 b | 0.53 bc | 0.13 c | |
| F | 7.571 | 1.466 | 15.131 | 1.860 | 44.395 | 7.571 | 1.466 | 15.131 | 1.860 | 44.395 | |
| Trunks | |||||||||||
| Merlot | Cabernet Franc | ||||||||||
| N | P | K | Ca | Mg | N | P | K | Ca | Mg | ||
| Control | Own roots | 0.61 c | 0.28 a | 0.35 b | 0.90 a | 0.15 b | 0.63 b | 0.26 a | 0.24 bc | 0.94 a | 0.18 a |
| 1103P | 0.71 a | 0.19 b | 0.27 c | 0.89 a | 0.16 ab | 0.64 b | 0.20 b | 0.25 ab | 0.91 ab | 0.16 b | |
| 101-14 Mgt | 0.687 ab | 0.17 b | 0.43 a | 0.83 a | 0.17 a | 0.71 a | 0.23 ab | 0.28 a | 0.82 bc | 0.14 c | |
| 120 μM Cr(VI) | Own roots | 0.51 d | 0.08 c | 0.28 c | 0.62 b | 0.12 c | 0.55 c | 0.16 c | 0.23 bc | 0.74 cd | 0.12 d |
| 1103P | 0.56 cd | 0.11 c | 0.25 cd | 0.81 a | 0.13 c | 0.65 ab | 0.13 cd | 0.19 d | 0.76 cd | 0.13 cd | |
| 101-14 Mgt | 0.63 bc | 0.09 c | 0.22 d | 0.56 b | 0.15 b | 0.60 bc | 0.11 d | 0.21 cd | 0.68 d | 0.13 cd | |
| F | 2.550 | 16.739 | 5.185 | 4.937 | 6.972 | 2.550 | 16.739 | 5.186 | 4.937 | 6.972 | |
| Roots | |||||||||||
| Merlot | Cabernet Franc | ||||||||||
| N | P | K | Ca | Mg | N | P | K | Ca | Mg | ||
| Control | Own roots | 1.05 | 0.26 b | 0.60 a | 1.18 | 0.24 b | 1.12 | 0.30 a | 0.65 a | 0.94 | 0.27 a |
| 1103P | 0.99 | 0.28 a | 0.50 a | 1.39 | 0.29 a | 1.06 | 0.28 b | 0.48 b | 0.90 | 0.25 ab | |
| 101-14 Mgt | 1.05 | 0.23 c | 0.53 a | 1.03 | 0.24 b | 1.10 | 0.27 b | 0.42 bc | 1.05 | 0.26 a | |
| 120 μM Cr(VI) | Own roots | 0.83 | 0.22 c | 0.38 b | 1.24 | 0.22 bc | 0.98 | 0.22 cd | 0.39 bc | 1.16 | 0.27 a |
| 1103P | 0.85 | 0.18 d | 0.27 c | 0.96 | 0.20 c | 0.87 | 0.21 d | 0.24 d | 1.24 | 0.25 ab | |
| 101-14 Mgt | 0.88 | 0.22 c | 0.50 a | 0.96 | 0.19 c | 0.89 | 0.23 c | 0.36 c | 1.05 | 0.22 b | |
| F | 12.907 | 8.597 | 2.663 | 12.907 | 8.597 | 2.663 | |||||
| Leaves | |||||||||||||
| Merlot | Cabernet Franc | ||||||||||||
| [mg kg−1] | Fe | Mn | Zn | B | Cu | Cr | Fe | Mn | Zn | B | Cu | Cr | |
| Control | Own roots | 107.89 ab | 68.23 b | 18.26 a | 56.84 ab | 9.89 a | 0.47 c | 112.37 bc | 62.69 a | 13.42 b | 46.03 ab | 8.35 a | 0.63 c |
| 1103P | 121.29 a | 96.04 a | 12.86 c | 62.17 a | 7.71 b | 0.55 c | 136.27 a | 63.88 a | 12.73 bc | 51.92 a | 6.02 bc | 0.16 c | |
| 101-14 Mgt | 100.56 b | 52.90 c | 13.00 c | 51.06 b | 6.12 bc | 0.69 c | 122.76 ab | 48.20 bc | 15.50 a | 45.91 ab | 6.45 ab | 0.62 c | |
| 120 μM Cr(VI) | Own roots | 96.81 b | 48.42 cd | 14.05 b | 46.37 b | 5.02 c | 3.49 a | 102.53 cd | 56.55 ab | 11.80 c | 41.84 b | 5.34 bc | 3.01 ab |
| 1103P | 94.74 b | 39.26 de | 11.47 d | 44.85 b | 4.11 c | 2.01 b | 92.03 d | 48.37 bc | 8.75 d | 45.33 ab | 4.21 c | 2.15 b | |
| 101-14 Mgt | 105.47 b | 36.69 e | 11.85 d | 47.35 b | 4.81 c | 3.22 a | 90.89 d | 39.97 c | 9.21 d | 40.37 b | 4.32 c | 3.28 a | |
| F | 10.024 | 23.921 | 25.634 | 24.486 | 11.889 | 28.305 | 10.024 | 23.921 | 25.634 | 24.486 | 11.889 | 28.305 | |
| Shoots | |||||||||||||
| Merlot | Cabernet Franc | ||||||||||||
| Fe | Mn | Zn | B | Cu | Cr | Fe | Mn | Zn | B | Cu | Cr | ||
| Control | Own roots | 24.27 a | 44.84 ab | 30.79 a | 15.78 a | 10.46 a | 0.66 b | 25.90 a | 30.42 b | 24.90 b | 13.43 b | 7.62 bc | 0.75 b |
| 1103P | 21.13 a | 43.27 b | 22.42 b | 15.00 a | 7.99 a | 0.54 b | 23.77 ab | 46.70 a | 31.67 a | 14.24 b | 10.71 a | 0.59 b | |
| 101-14 Mgt | 24.69 a | 50.03 a | 31.35 a | 14.96 a | 10.27 a | 0.48 b | 24.08 ab | 40.83 a | 16.25 cd | 16.55 a | 8.31 ab | 0.52 b | |
| 120 μM Cr(VI) | Own roots | 15.92 b | 24.74 d | 14.79 d | 11.03 c | 7.54 a | 1.90 ab | 21.70 b | 29.71 b | 14.86 d | 9.39 c | 5.30 c | 2.98 a |
| 1103P | 15.07 b | 34.83 c | 14.02 d | 13.18 b | 6.64 b | 2.47 a | 15.47 c | 28.55 b | 17.36 c | 9.10 c | 5.13 c | 2.06 ab | |
| 101-14 Mgt | 17.09 b | 44.22 ab | 20.33 c | 12.85 b | 7.63 a | 2.60 a | 16.95 c | 21.72 c | 10.29 e | 9.77 c | 5.64 bc | 2.11 ab | |
| F | 6.661 | 15.88 | 33.063 | 15.281 | 27.921 | 37.616 | 6.661 | 15.88 | 33.063 | 15.281 | 27.921 | 37.616 | |
| Trunks | |||||||||||||
| Merlot | Cabernet Franc | ||||||||||||
| Fe | Mn | Zn | B | Cu | Cr | Fe | Mn | Zn | B | Cu | Cr | ||
| Control | Own roots | 110.03 a | 24.40 b | 18.95 ab | 11.55 a | 8.29 abc | 3.18 b | 101.49 a | 18.57 b | 21.10 a | 11.48 a | 8.96 abc | 4.50 c |
| 1103P | 107.37 ab | 27.02 a | 15.72 abc | 10.53 b | 9.92 a | 4.19 b | 109.38 a | 24.97 a | 18.92 ab | 11.01 ab | 9.82 a | 3.34 c | |
| 101-14 Mgt | 99.24 ab | 25.85 ab | 20.07 a | 10.46 b | 9.49 ab | 5.24 b | 97.67 ab | 23.81 a | 19.30 ab | 10.58 a | 9.54 ab | 3.64 c | |
| 120 μM Cr(VI) | Own roots | 58.75 c | 18.19 d | 10.26 bc | 9.14 c | 5.47 c | 95.28 a | 73.13 abc | 19.21 b | 9.52 c | 8.41 c | 5.98 bc | 102.24 a |
| 1103P | 78.40 abc | 23.63 b | 10.58 bc | 8.96 c | 6.01 bc | 58.76 ab | 54.27 c | 19.61 b | 10.52 bc | 8.92 c | 5.55 c | 40.95 bc | |
| 101-14 Mgt | 70.11 bc | 20.75 c | 9.89 c | 9.01 c | 5.89 bc | 81.28 a | 60.80 bc | 18.57 b | 9.24 c | 8.96 c | 5.46 c | 62.23 ab | |
| F | 14.320 | 6.983 | 25.058 | 7.903 | 10.334 | 16.902 | 14.32 | 6.983 | 25.058 | 7.903 | 10.334 | 16.902 | |
| Roots | |||||||||||||
| Merlot | Cabernet Franc | ||||||||||||
| Fe | Mn | Zn | B | Cu | Cr | Fe | Mn | Zn | B | Cu | Cr | ||
| Control | Own roots | 213.24 ab | 32.22 a | 26.72 b | 20.88 c | 26.23 a | 11.3 c | 230.50 a | 35.27 a | 22.34 c | 24.99 b | 31.76 ab | 11.94 d |
| 1103P | 202.96 abc | 27.79 b | 30.61 a | 22.20 b | 25.84 a | 6.22 c | 204.52 ab | 26.19 b | 29.23 a | 29.82 a | 24.31 abc | 9.71 d | |
| 101-14 Mgt | 245.38 a | 35.90 a | 30.37 a | 23.24 a | 30.90 a | 11.63 c | 231.23 a | 32.41 ab | 24.81 b | 23.70 c | 33.14 a | 11.85 d | |
| 120 μM Cr(VI) | Own roots | 160.63 bcd | 22.83 bc | 15.69 c | 19.34 d | 19.96 ab | 658.81 a | 163.76 b | 27.66 b | 15.17 d | 24.77 b | 17.89 c | 868.34 a |
| 1103P | 136.24 d | 29.04 b | 16.97 c | 19.19 d | 12.95 b | 396.05 b | 177.60 ab | 30.01 ab | 16.81 d | 23.21 c | 18.56 c | 570.76 c | |
| 101-14 Mgt | 151.13 cd | 20.23 c | 17.15 c | 16.95 e | 18.70 ab | 427.19 b | 175.52 ab | 28.16 b | 17.19 d | 23.11 c | 19.64 bc | 718.45 b | |
| F | 5.262 | 2.535 | 16.015 | 5.532 | 11.928 | 117.731 | 5.262 | 2.535 | 16.015 | 5.532 | 11.928 | 117.73 | |
| Merlot | Cabernet Franc | ||||
|---|---|---|---|---|---|
| Thirty Days | Sixty Days | Thirty Days | Sixty Days | ||
| Control | Own roots | 0.783 bc | 0.721 b | 0.779 b | 0.721 a |
| 1103P | 0.820 ab | 0.732 ab | 0.791 ab | 0.744 a | |
| 101-14 Mgt | 0.837 a | 0.771 a | 0.821 a | 0.716 a | |
| 120 μM Cr(VI) | Own roots | 0.761 c | 0.675 c | 0.718 d | 0.576 b |
| 1103P | 0.800 abc | 0.501 d | 0.771 bc | 0.579 b | |
| 101-14 Mgt | 0.773 c | 0.494 d | 0.736 cd | 0.482 c | |
| F: 19.015 | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nikolaou, K.-E.; Chatzistathis, T.; Theocharis, S.; Argiriou, A.; Koundouras, S.; Zioziou, E. Effects of Chromium Toxicity on Physiological Performance and Nutrient Uptake in Two Grapevine Cultivars (Vitis vinifera L.) Growing on Own Roots or Grafted onto Different Rootstocks. Horticulturae 2022, 8, 493. https://doi.org/10.3390/horticulturae8060493
Nikolaou K-E, Chatzistathis T, Theocharis S, Argiriou A, Koundouras S, Zioziou E. Effects of Chromium Toxicity on Physiological Performance and Nutrient Uptake in Two Grapevine Cultivars (Vitis vinifera L.) Growing on Own Roots or Grafted onto Different Rootstocks. Horticulturae. 2022; 8(6):493. https://doi.org/10.3390/horticulturae8060493
Chicago/Turabian StyleNikolaou, Kleopatra-Eleni, Theocharis Chatzistathis, Serafeim Theocharis, Anagnostis Argiriou, Stefanos Koundouras, and Elefteria Zioziou. 2022. "Effects of Chromium Toxicity on Physiological Performance and Nutrient Uptake in Two Grapevine Cultivars (Vitis vinifera L.) Growing on Own Roots or Grafted onto Different Rootstocks" Horticulturae 8, no. 6: 493. https://doi.org/10.3390/horticulturae8060493