
Genetic Diversity and Streptomycin Sensitivity in Xanthomonas axonopodis pv. punicae Causing Oily Spot Disease in Pomegranates

 , , ,
, , ,
Abstract
:1. Introduction
2. Material and Methods
2.1. Collection of Xanthomonas axonopodis pv. punicae (Xap) Isolates
2.2. Pathogenicity Assay
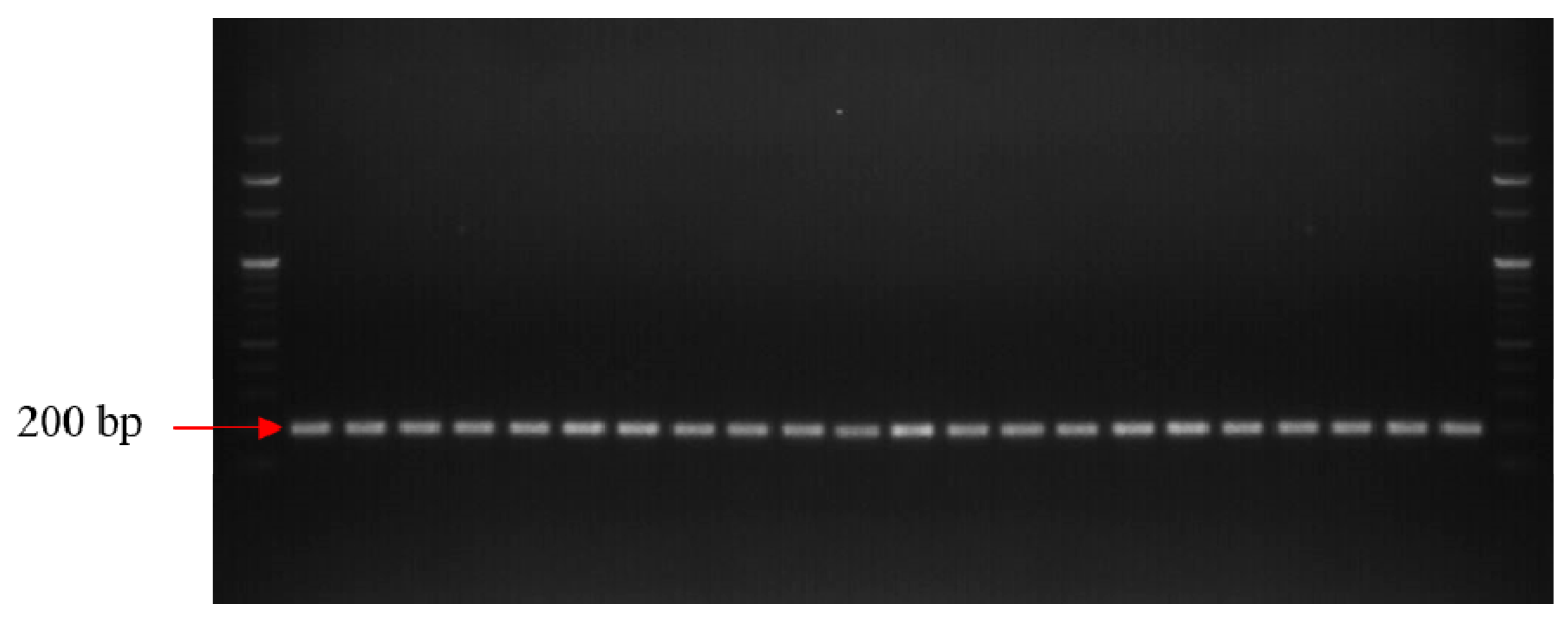
2.3. Confirmation of Identity of Xap Isolates and Detection of Variability Amongst Isolates Using SSR Markers
2.4. Data Analysis
2.5. Screening for Streptomycin Sensitivity In Vitro
2.6. Molecular Confirmation of Streptomycin Resistance Genes
2.7. Molecular Characterization of rpsL Gene
3. Results
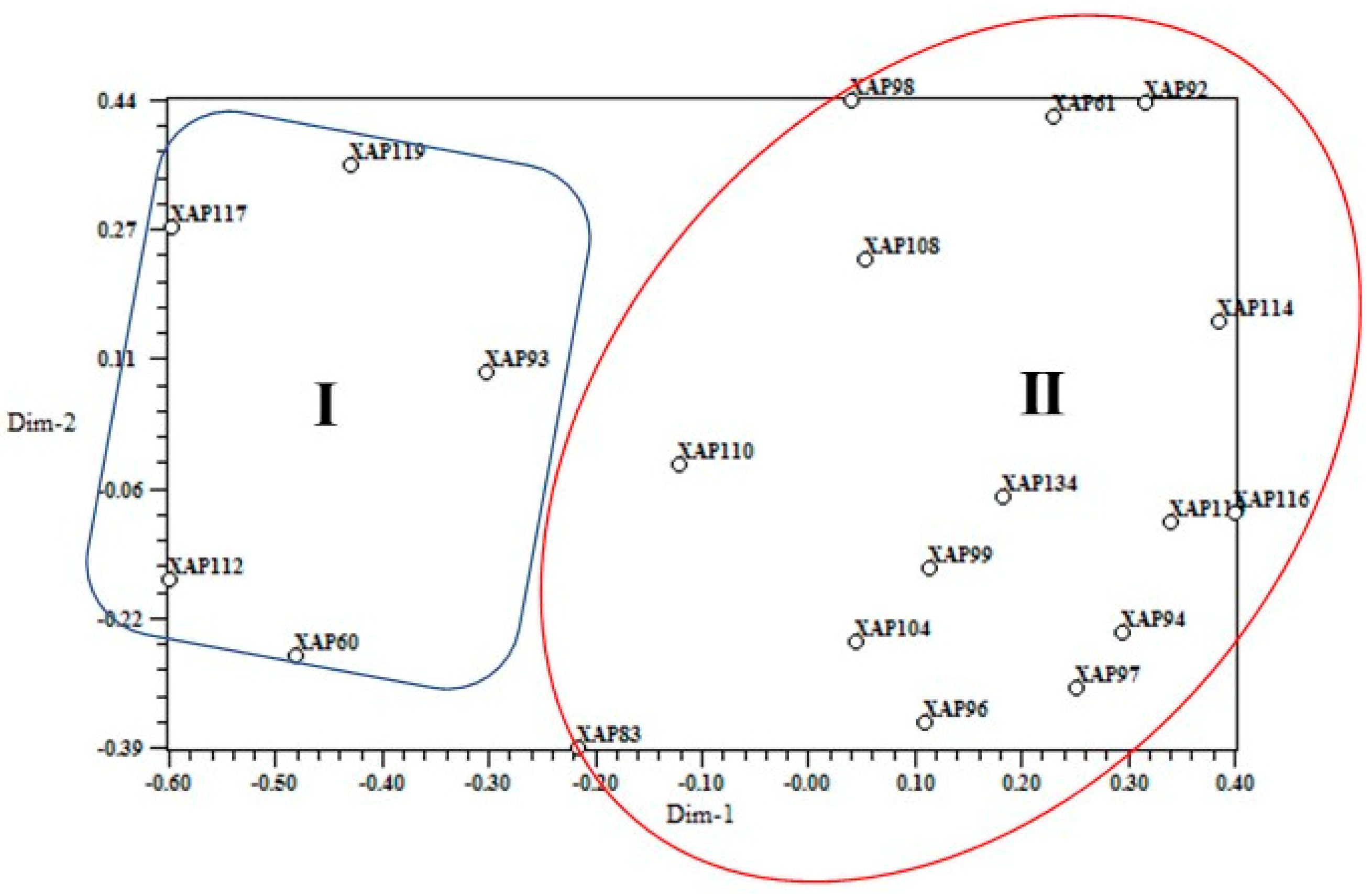
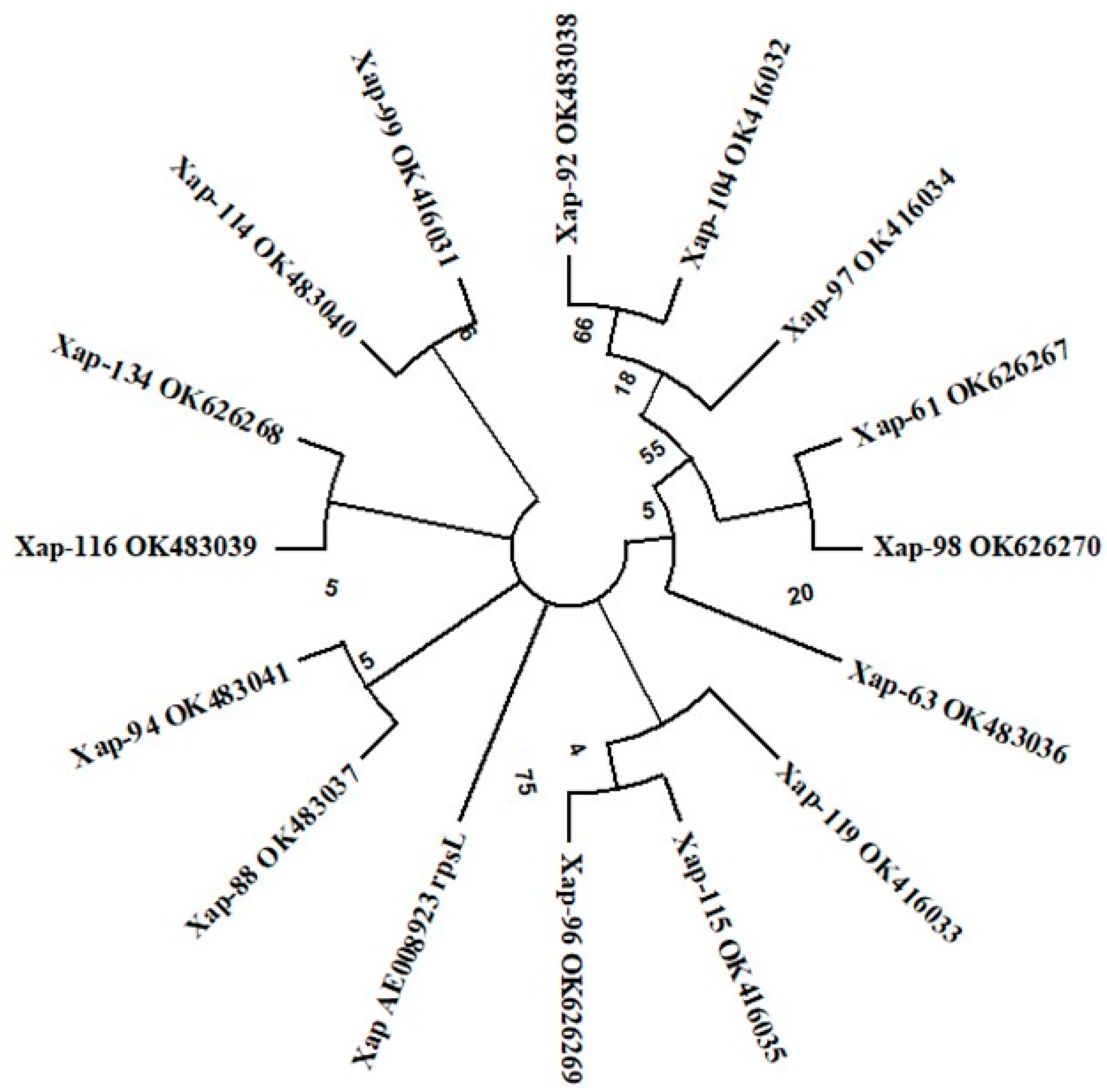
3.1. Identity Confirmation and Genetic Diversity Analysis
3.2. Sensitivity to Streptomycin and Its Molecular Basis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Department of Agriculture, Cooperation & Farmers Welfare. Agricultural-Statistics at a Glance 2020; Directorate of Economics and Statistics, Government of India: New Delhi, India, 2020.
- Pal, R.K.; Sharma, J.; Babu, D.; Singh, N.V.; Gaikwad, N. Pomegranate Revolution in India-A Success Story of ICAR.; ICAR-NRC on Pomegranate Solapur: Solapur, India, 2017. [Google Scholar]
- Sharma, J.; Manjunath, G.; Xavier, K.V.; Vallad, G.E. Diseases and Management. In The Pomegranate: Botany, Production and Uses; Sarkhosh, A., Yavari, A.M., Zamani, Z., Eds.; Centre for Agriculture and Bioscience International: Wallingford, UK, 2021; pp. 357–391. [Google Scholar] [CrossRef]
- Sharma, J.; Maithy, A.; Singh, N.V.; Mallikarjun Manjunatha, N.; Pokhare, S. Bimonthly Pomegranate Advisory for Bearing Orchards; NRC-P: Solapur, India, 2022. [Google Scholar]
- Benagi, V.L.; RaviKumar, M.R.; Nargund, V.B. Threat of Bacterial blight on Pomegranate in India—Mitigation by an Integrated Approach. II International Symposium on the Pomegranate 113–116. 2012. Available online: http://om.ciheam.org/article.php?IDPDF=6917 (accessed on 10 March 2022).
- Schatz, A.; Bugie, E.; Waksman, S.A. Streptomycin, a substance exhibiting antibiotic activity against gram-positive and gram-negative bacteria. Proc. Soc. Exp. Biol. Med. 1944, 55, 66–69. [Google Scholar] [CrossRef]
- Rani, U.; Verma, K.S. Perpetuation and spread of Xanthomonas axonopodis pv. punicae causing black spot of pomegranate. Plant Dis. Res. Ludhiana 2002, 17, 46–50. [Google Scholar]
- Sharma, J.; Sharma, K.K.; Jadhav, V.T. Diseases of Pomegranate. In Diseases of Fruit Crops; Misra, A.K., Chowdappa, P., Sharma, P., Khetrapal, R.K., Eds.; Indian Phytopathological Society: New Delhi, India, 2012; pp. 181–224. [Google Scholar]
- Sundin, G.W.; Wang, N. Antibiotic resistance in plant-pathogenic bacteria. Annu. Rev. Phytopathol. 2018, 56, 161–180. [Google Scholar] [CrossRef] [PubMed]
- Pezzella, C.; Ricci, A.; DiGiannatale, E.; Luzzi, I.; Carattoli, A. Tetracycline and streptomycin resistance genes, transposons, and plasmids in Salmonella enterica isolates from animals in Italy. Antimicrob. Agents Chemother. 2004, 48, 903–908. [Google Scholar] [CrossRef] [Green Version]
- Araújo, E.R.; Pereira, R.C.; Ferreira, M.A.S.V.; Quezado-Duval, A.M.; Café-Filho, A.C. Sensitivity of xanthomonads causing tomato bacterial spot to copper and streptomycin and in vivo infra-specific competitive ability in Xanthomonas perforans resistant and sensitive to copper. J. Plant Pathol. 2012, 94, 79–87. Available online: https://www.jstor.org/stable/45156011 (accessed on 10 March 2022).
- Förster, H.; McGhee, G.C.; Sundin, G.W.; Adaskaveg, J.E. Characterization of streptomycin resistance in isolates of Erwinia amylovora in California. Phytopathology 2015, 105, 1302–1310. [Google Scholar] [CrossRef] [Green Version]
- Wiener, P.; Egan, S.; Wellington, E.M.H. Evidence for transfer of antibiotic-resistance genes in soil populations of streptomy-cetes. Mol. Ecol. 1998, 7, 1205–1216. [Google Scholar] [CrossRef]
- Chiou, C.-S.; Jones, A.L. Nucleotide sequence analysis of a transposon (Tn5393) carrying streptomycin resistance genes in Erwinia amylovora and other gram-negative bacteria. J. Bacteriol. 1993, 175, 732–740. [Google Scholar] [CrossRef] [Green Version]
- Hall, R.M.; Collis, C.M. Mobile gene cassettes and integrons: Capture and spread of genes by site specific recombination. Mol. Microbiol. 1995, 15, 593–600. [Google Scholar] [CrossRef]
- Ozaki, M.; Mizushima, S.; Nomura, M. Identification and functional characterization of the protein controlled by the strep-tomycin-resistant locus in E. coli. Nature 1969, 222, 333–339. [Google Scholar] [CrossRef]
- Chiou, C.-S.; Jones, A.L. Molecular analysis of high-level streptomycin resistance in Erwinia amylovora. Phytopathology 1995, 85, 324–328. [Google Scholar] [CrossRef]
- Behlau, F.; Jones, J.B.; Myers, M.E.; Graham, J.H. Monitoring for resistant populations of Xanthomonas citri subsp. citri and epiphytic bacteria on citrus trees treated with copper or streptomycin using a new semi-selective medium. Eur. J. Plant Pathol. 2012, 132, 259–270. [Google Scholar] [CrossRef]
- Xu, Y.; Zhu, X.F.; Zhou, M.G.; Kuang, J.; Zhang, Y.; Shang, Y.; Wang, J.X. Status of streptomycin resistance development in Xanthomonas oryzae pv. oryzae and Xanthomonas oryzae pv. oryzicola in China and their resistance characters. J. Phytopathol. 2010, 158, 601–608. [Google Scholar] [CrossRef]
- Krishna, P.; Prasanna Kumar, M.K.; Channappa, M.; Devanna, P.; Singh, K.; Eeregowda, P.M.; Mahesh, H.B.; Chandrashekar, B.S.; Babu, V.; Desai, R.U.; et al. Antibiotic resilience in Xanthomonas axonopodis pv. punicae causing bacterial blight of pome-granate. Curr. Sci. 2020, 119, 1564–1569. [Google Scholar] [CrossRef]
- Sharma, J.; Sharma, K.K.; Kumar, A.; Mondal, K.K.; Thalor, S.; Maity, A.; Gharate, R.; Chinchure, S.; Jadhav, V.T. Pomegranate bacterial blight: Symptomatology and rapid inoculation technique for Xanthomonas axonopodis pv. punicae. J. Plant Pathol. 2017, 99, 109–119. Available online: https://www.jstor.org/stable/44280579 (accessed on 10 March 2022).
- Kumar, A.; Sharma, J.; Munjal, V.; Sakthivel, K.; Thalor, S.K.; Mondal, K.K.; Chinchure, S.; Gharate, R. Polyphasic phenotypic and genetic analysis reveals clonal nature of Xanthomonas axonopodis pv. punicae causing pomegranate bacterial blight. Plant Pathol. 2020, 69, 347–359. [Google Scholar] [CrossRef]
- Doddaraju, P.; Kumar, P.; Gunnaiah, R.; Gowda, A.A.; Lokesh, V.; Pujer, P.; Manjunatha, G. Reliable and early diagnosis of bacterial blight in pomegranate caused by Xanthomonas axonopodis pv. punicae using sensitive PCR techniques. Sci. Rep. 2019, 9, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Liu, K.; Muse, S.V. PowerMarker: An integrated analysis environment for genetic marker analysis. Bioinformatics 2005, 21, 2128–2129. [Google Scholar] [CrossRef] [Green Version]
- Jaccard, P. Nouvelles recherches sur la distri-bution fiorale. Bull. Soc. Vaud. Sci. Nat. 1908, 44, 223–270. [Google Scholar]
- Rohlf, F.J. NTSYS-pc: Numerical Taxonomy and Multivariate Analysis System; Version 2.0; Exeter Software: Setauket, NY, USA, 1997. [Google Scholar]
- Hall, T. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Hingorani, M.K.; Mehta, P.P. Bacterial leaf spot of pomegranate. Indian Phytopathol. 1952, 5, 55–56. [Google Scholar]
- Hingorani, M.K.; Singh, N.J. Xanthomonas punicae sp. now on Punica granatum. Indian J. Agric. Sci. 1959, 29, 45–48. [Google Scholar]
- Sohi, H.S.; Jain, S.S.; Sharma, S.L.; Verma, B.R. New records of plant diseases from H.P. Indian Phytopathol. 1964, 17, 42–45. [Google Scholar]
- Kanwar, Z.S. A note on bacterial disease of pomegranate (Punica granatum L.) in Haryana. Haryana J. Hort. Sci. 1976, 5, 177–180. [Google Scholar]
- Dhandar, D.G.; Nallathambi, P.; Rawat, R.D.; Sawant, D.M. Bacterial leaf and fruit spot-A new threat to pomegranate or-chards in Maharashtra. J. Mycol. Plant Pathol. 2004, 34, 971. [Google Scholar]
- Akhtar, M.A.; Bhatti, M.H.R. Occurrence of bacterial leaf spot of pomegranate in Pakistan. Pak. J. Agric. Sci. 1992, 13, 95–97. [Google Scholar]
- Petersen, Y.; Mansvelt, E.L.; Venter, E.; Langenhoven, W.E. Detection of Xanthomonas axonopodis pv. punicae causing bacterial blight on pomegranate in South Africa. Australas. Plant Pathol. 2010, 39, 544–546. [Google Scholar] [CrossRef]
- Icoz, S.M.; Polat, I.; Sulu, G.; Yilmaz, M.; Unlu, A.; Soylu, S.; Bozkurt, I.A.; Baysal, O. First report of bacterial blight of pome-granate caused by Xanthomonas axonopodis pv. punicae in Turkey. Plant Dis. 2014, 98, 427. [Google Scholar] [CrossRef]
- Mondal, K.K.; Rajendran, T.P.; Phaneendra, C.; Mani, C.; Sharma, J.; Shukla, R.; Verma, G.; Kumar, R.; Singh, D.; Kumar, A.; et al. The reliable and rapid polymerase chain reaction (PCR) diagnosis for Xanthomonas axonopodis pv. punicae in pomegranate. Afr. J. Microbiol. Res. 2012, 6, 5950–5956. [Google Scholar]
- Prasannakumar, M.K.; Buela Parivallal, P.; Manjunatha, C.; Mahesh, H.B.; Pramesh, D.; Narayan, K.S.; Gopal, V.B.; Priyanka, K.; Puneeth, M.E.; Rangaswamy, K.T. Loop-mediated isothermal amplification assay for pre-symptomatic stage de-tection of Xanthomonas axonopodis pv. punicae infection in pomegranate. Australas. Plant Pathol. 2020, 49, 467–473. [Google Scholar] [CrossRef]
- Alvarez-Martinez, C.E.; Sgro, G.G.; Araujo, G.G.; Paiva, M.R.; Matsuyama, B.Y.; Guzzo, C.R.; Andrade, M.O.; Farah, C.S. Secrete or perish: The role of secretion systems in Xanthomonas biology. Comput. Struct. Biotechnol. J. 2021, 19, 279–302. [Google Scholar] [CrossRef] [PubMed]
- Mahfooz, S.; Srivastava, A.; Yadav, M.C.; Tahoor, A. Comparative genomics in phytopathogenic prokaryotes reveals the higher relative abundance and density of long-SSRs in the smallest prokaryotic genome. 3 Biotech 2019, 9, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Moxon, R.; Bayliss, C.; Hood, D. Bacterial contingency loci: The role of simple sequence DNA repeats in bacterial adaptation. Annu. Rev. Phytopathol. 2006, 40, 307–333. [Google Scholar] [CrossRef] [PubMed]
- Saurabh, B.; Sneha, S.; Suvidya, R.; Pramod, K.; Shailesh, B. Analysis of distribution and significance of simple sequence repeats in enteric bacteria Shigella dysenteriae SD197. Bioinformation 2011, 6, 348–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Field, D.; Wills, C. Abundant microsatellite polymorphism in Saccharomyces cerevisiae, and the different distributions of mi-crosatellites in eight prokaryotes and S. cerevisiae, result from strong mutation pressures and a variety of selective forces. Proc. Natl. Acad. Sci. USA 1998, 95, 1647–1652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tardiani, A.C.; Perecin, D.; Peixoto-Junior, R.F.; Sanguino, A.; Landell, M.M.; Beriam, L.O.; Nunes, D.S.; Camargo, L.E.; Creste, S. Molecular and pathogenic diversity among Brazilian isolates of Xanthomonas albilineans assessed with SSR marker loci. Plant Dis. 2014, 98, 540–546. [Google Scholar] [CrossRef] [Green Version]
- Anand, G.; Kapoor, R. Population structure and virulence analysis of Alternaria carthami isolates of India using ISSR and SSR markers. World J. Microbiol. Biotechnol. 2018, 34, 140. [Google Scholar] [CrossRef]
- McGhee, G.C.; Guasco, J.; Bellomo, L.M.; Blumer-Schuette, S.E.; Shane, W.W.; Irish-Brown, A.; Sundin, G.W. Genetic analysis of streptomycin-resistant (SmR) strains of Erwinia amylovora suggests that dissemination of two genotypes is responsible for the current distribution of SmR E. amylovora in Michigan. Phytopathology 2011, 192, 182–191. [Google Scholar] [CrossRef] [Green Version]
- Sundin, G.W.; Bender, C.L. Expression of the strA-strB streptomycin resistance genes in Pseudomonas syringae and Xanthomonas campestris and characterization of IS6100 in X. campestris. Appl. Environ. Microbiol. 1995, 61, 2891–2897. [Google Scholar] [CrossRef] [Green Version]
- Hyun, J.-W.; Kim, H.-J.; Yi, P.-H.; Hwang, R.-Y.; Park, E.-W. Mode of action of streptomycin resistance in the citrus canker pathogen (Xanthomonas smithii subsp. citri) in Jeju Island. Plant Pathol. J. 2012, 28, 207–211. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Li, Q.; Gao, H.; Zhang, Z.; Liu, Y.; Lu, J.; Dai, E. The roles of rpsL, rrs, and gidB mutations in predicting streptomy-cin-resistant drugs used on clinical Mycobacterium tuberculosis isolates from Hebei Province, China. Int. J. Clin. Exp. Pathol. 2019, 12, 2713–2721. [Google Scholar] [PubMed]
- Dai, R.; He, J.; Zha, X.; Wang, Y.; Zhang, X.; Gao, H.; Yang, X.; Li, J.; Xin, Y.; Wang, Y.; et al. A novel mechanism of streptomycin resistance in Yersinia pestis: Mutation in the rpsL gene. PLoS Negl. Trop. Dis. 2021, 15, e0009324. [Google Scholar] [CrossRef] [PubMed]
- Escursell, M.M.; Roschi, A.; Smits, T.H.; Rezzonico, F. Characterization and direct molecular discrimination of rpsL mutations leading to high streptomycin resistance in Erwinia amylovora. J. Plant Pathol. 2021, 103, 99–108. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sr. No. | Isolate Code | State in India | GPS Coordinates |
|---|---|---|---|
| 1. | XAP-61 | Maharashtra | 18°14′00″ N 75°41′34″ E |
| 2. | XAP-63 | Maharashtra | 17°50′00″ N 75°51′00″ E |
| 3. | XAP-88 | Maharashtra | 18°17′21″ N 75°41′15″ E |
| 4. | XAP-99 | Maharashtra | 17°43′46”N 75°17′19″ E |
| 5. | XAP-104 | Maharashtra | 17°40′30″ N 75°19′36″ E |
| 6. | XAP-114 | Maharashtra | 17°30′39″ N 75°27′07″ E |
| 7. | XAP-119 | Maharashtra | 17°23′52″ N 75°48′03″ E |
| 8. | XAP-134 | Maharashtra | 17°31′27″ N 76°12′19″ E |
| 9. | XAP-93 | Maharashtra | 17°29′04″ N 75°14′05″ E |
| 10. | XAP-117 | Maharashtra | 18°34′48″ N 73°52′25″ E |
| 11. | XAP-96 | Maharashtra | 18°37′35″ N 75°04′43″ E |
| 12. | XAP-98 | Maharashtra | 17°43′08″ N 75°50′38″ E |
| 13. | XAP-108 | Maharashtra | 21°33′18″ N 74°28′12″ E |
| 14. | XAP-83 | Maharashtra | 17°43′09″ N 76°01′26″ E |
| 15. | XAP-92 | Himachal Pradesh | 31°57′33″ N 77°06′32″ E |
| 16. | XAP-60 | Karnataka | 13°08′05″ N 77°29′45″ E |
| 17. | XAP-97 | Karnataka | 16°10′08″ N 75°39′41″ E |
| 18. | XAP-112 | Karnataka | 17°19′47″ N 76°50′03″ E |
| 19. | XAP-94 | Telangana | 16°36′12″ N 77°43′35″ E |
| 20. | XAP-110 | Tamil Nadu | 13°52′26″ N 75°34′01″ E |
| 21. | XAP-115 | Uttar Pradesh | 26°31′36″ N 79°49′46″ E |
| 22. | XAP-116 | Rajasthan | 25°45′07″ N 71°23′48″ E |
| Name | Forward (5′-3′) | Reverse (5′-3′) | Amplicon Size (bp) |
|---|---|---|---|
| XopQ | GCGAGGAACTTGGAATGCTC | AGGTCGAAGGCTTTTTGCG | 190 |
| strA | CCAAGTCAGAGGGTCCAATC | TGACTGGTTGCCTGTCAGAG | 760 |
| StrB | TAGATCGCGTTGCTCCTCTT | ACGTTTCGCAACCTGTTCTC | 758 |
| rpsL | CAAGCGACCACCTACAAGAGT | GTACTTGGAACGGCCTTGAC | 315 |
| Sr. No. | Primer | Motif | Primer Sequence F/R (5′-3′) | MAF | Allele No. | Gene Diversity | PIC | Expected Amplicon Size (bp) |
|---|---|---|---|---|---|---|---|---|
| 1 | XAP_SSR42 | (CG)7 | GACACAGACCAACGGATTC/GGTGACATTGACCTGCTC | 0.70 | 2 | 0.42 | 0.33 | 253 |
| 2 | XAP_SSR38 | (CGG)6 | CACCGGCACACTCAATAC/ATGCATCCTGCTGCTGAT | 0.65 | 2 | 0.46 | 0.35 | 160 |
| 3 | XAP_SSR59 | (CG)6 | GAGCGGTTCACAGGAAAT/GTCGACGAACAACAGCAT | 0.75 | 2 | 0.38 | 0.30 | 223 |
| 4 | XAP_SSR60 | (CG)7 | AGGGTTGAGCGTGTTTCT/CACTGGATCTGACCATCAAG | 0.75 | 2 | 0.38 | 0.30 | 249 |
| 5 | Xap_SSR28 | (ACC)5 | AGCGACCAGTCCATCTATC/CTCCAGCAGGTAGCTCAG | 0.56 | 2 | 0.49 | 0.37 | 291 |
| 6 | XAP_SSR6 | (TGC)4 | CTTCCTCGATCATCACCTC/CGCAGTGATCTTCGTCAT | 0.61 | 2 | 0.48 | 0.36 | 212 |
| 7 | XAP_SSR22 | (CCA)5 | ATGCGAGGTACCATGCTC/CTCTGCCTCAAGGTCTCTTT | 0.78 | 2 | 0.35 | 0.29 | 278 |
| 8 | XAP_SSR27 | (CT)7 | GTTGCTTCCTACGACAGG/GCGCTTCACAGATCGTAT | 0.55 | 2 | 0.50 | 0.37 | 254 |
| 9 | XAP_SSR24 | (GCA)4 | CAGCGATGTGGTGATTTC/GAGCATGCTGGTCAGAAC | 0.60 | 2 | 0.48 | 0.36 | 228 |
| 10 | XAP_SSR43 | (CAT)4 | CTGAACATCGTGGTGCTG/CTGGCTGTTCTGGATCAC | 0.53 | 2 | 0.50 | 0.37 | 252 |
| 11 | XAP_SSR33 | (CG)6 | CCTGATCAGCCGGTATTC/CACACCACACGGTCCATC | 0.71 | 2 | 0.42 | 0.33 | 300 |
| 12 | XAP_SSR53 | (CCA)4 | AGATCCAGACCTGTTCGAC/CTGAACCATCTGCGTACC | 0.53 | 2 | 0.50 | 0.37 | 247 |
| 13 | XAP_SSR13 | (TGG)4 | GTCGATCTGGTGGAAGAAC/AGGAACGAGGAAAACTGC | 0.88 | 2 | 0.21 | 0.19 | 254 |
| 14 | XAP_SSR17 | (CGC)4 | GACATGATCGACGTCTCC/CAGTTGTCCTTGACCTCGT | 0.53 | 2 | 0.50 | 0.37 | 233 |
| 15 | XAP_SSR10 | (GCG)4 | CTGACCATCGTCAATTCC/CATAGGCGATGTTGTTGG | 0.68 | 2 | 0.43 | 0.34 | 283 |
| 16 | XAP_SSR35 | (AGC)4 | GTACAGCTCGGATCAACG/AGGCTTACCAGATCACCAC | 0.80 | 2 | 0.32 | 0.27 | 227 |
| 17 | XAP_SSR65 | (GCC)4 | CTTGGGAAGTAACCACCAG/CGGTGCTGTCAATAGGTT | 0.78 | 2 | 0.35 | 0.29 | 252 |
| 18 | XAP_SSR23 | (GTC)4 | GTCAGTTGCAGTGCGTAAC/GGCTGATTACCTCGTGACT | 1.00 | 1 | 0.00 | 0.00 | 283 |
| Mean | 0.69 | 1.94 | 0.40 | 0.31 |
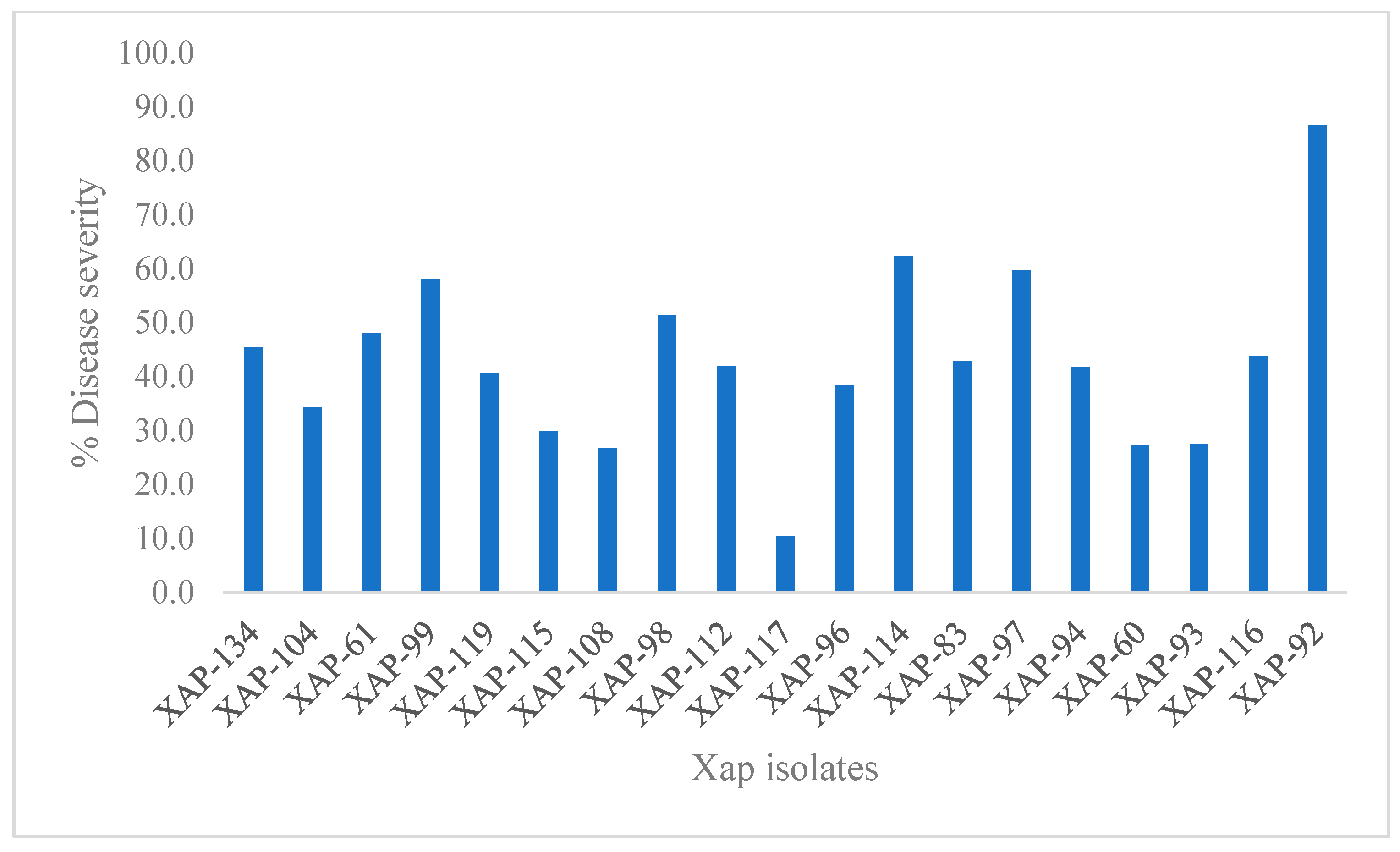
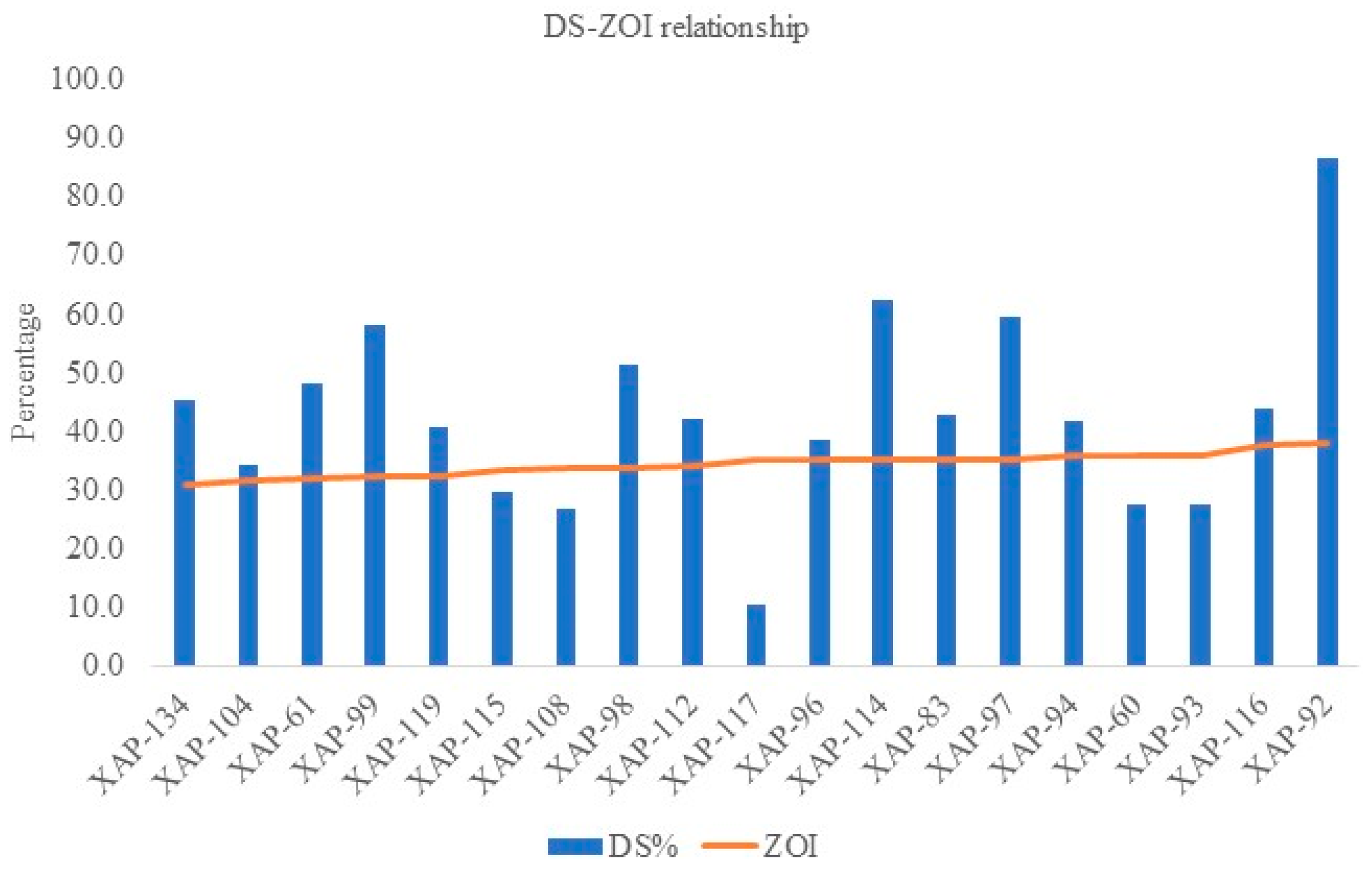
| S. No. | Isolate Code | Location | rpsL Mutation | strA/B | ZOI @ 1500 µg/mL (Non-Transformed) | Antibiotic Resilience/Sensitivity | % DS | Days Taken for Symptom Manifestation | Pathogenicity |
|---|---|---|---|---|---|---|---|---|---|
| Cluster 1 | |||||||||
| 1. | XAP-60 | KA | NA | - + | 34.07 | S | 27.3 | 3 weeks | low |
| 2. | XAP-112 | KA | NA | - + | 31.19 | S | 42.0 | 2 weeks | high |
| 3. | XAP-93 | MH | NA | + + | 34.44 | S | 27.5 | 3 weeks | low |
| 4. | XAP-117 | MH | NA | + - | 32.96 | S | 10.4 | 3 weeks | low |
| 5. | XAP-119 | MH | No | + + | 28.52 | R | 40.7 | 3 weeks | high |
| Cluster 2 | |||||||||
| 6. | XAP-108 | MH | NA | + + | 30.67 | MR | 26.7 | 3 weeks | low |
| 7. | XAP-83 | MH | NA | + + | 33.33 | S | 42.9 | 2 weeks | high |
| 8. | XAP-110 | TN | NA | - + | 30.33 | MR | NA | NA | NA |
| 9. | XAP-134 | MH | No | + + | 26.30 | R | 45.3 | 3 weeks | high |
| 10. | XAP-104 | MH | R-H | + + | 27.41 | R | 34.2 | 3 weeks | high |
| 11. | XAP-116 | RJ | No | - - | 37.04 | S | 43.8 | 2 weeks | high |
| 12. | XAP-97 | KA | R-H | - + | 33.33 | S | 59.7 | 3 weeks | high |
| 13. | XAP-96 | MH | N-T, S-F, A-D, Q-H, R-H | - + | 32.96 | S | 38.4 | 2 weeks | high |
| 14. | XAP-115 | UP | No | + + | 30.43 | MR | 29.8 | 3 weeks | low |
| 15. | XAP-114 | MH | No | - + | 33.33 | S | 62.3 | 3 weeks | high |
| 16. | XAP-94 | TL | No | - - | 34.07 | S | 41.7 | 3 weeks | high |
| 17. | XAP-99 | MH | No | + + | 28.52 | R | 58.0 | 2 weeks | high |
| 18. | XAP-92 | HP | R-H | + - | 37.80 | S | 86.7 | 2 weeks | Very high |
| 19. | XAP-98 | MH | R-H | + + | 30.74 | MR | 51.4 | 2 weeks | high |
| 20. | XAP-61 | MH | R-H | + + | 27.78 | R | 48.1 | 3 weeks | high |
| SSR data not available | |||||||||
| 21. | XAP-63 | MH | Y-I (insertion) | + + | 26.67 | R | NA | NA | NA |
| 22. | XAP-88 | MH | No | + + | 27.52 | R | NA | NA | NA |
| Isolates Treatment (µg/mL) | XAP-61 | XAP-63 | XAP-88 | XAP-99 | XAP-104 | XAP-114 | XAP-119 | XAP-134 | XAP-93 | XAP-117 | XAP-96 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 250 | 22.326 (14.44) | 24.074 (16.67) | 22.622 (14.81) | 20.111 (11.85) | 23.504 (15.93) | 26.177 (19.47) | 26.296 (19.63) | 22.622 (14.81) | 24.651 (17.41) | 27.597 (21.48) | 28.114 (22.22) |
| 500 | 23.509 (15.93) | 27.151 (20.84) | 25.805 (18.96) | 20.111 (11.85) | 23.509 (15.93) | 30.364 (25.56) | 27.603 (21.48) | 26.296 (19.63) | 27.85 (21.85) | 32.515 (28.89) | 31.32 (27.04) |
| 750 | 27.861 (21.85) | 30.351 (25.56) | 29.868 (24.81) | 27.34 (21.11) | 26.818 (20.37) | 33.664 (30.74) | 29.377 (24.07) | 28.114 (22.22) | 29.61 (24.44) | 34.817 (32.59) | 36.151 (34.81) |
| 1000 | 30.118 (25.19) | 29.627 (24.44) | 30.845 (26.30) | 28.377 (22.59) | 27.34 (21.11) | 32.979 (29.63) | 30.838 (26.30) | 28.627 (22.96) | 34.128 (31.48) | 34.809 (32.59) | 35.935 (34.44) |
| 1250 | 31.071 (26.67) | 31.562 (27.41) | 31.562 (27.41) | 30.351 (25.56) | 27.58 (21.48) | 33.203 (30.00) | 32.515 (28.89) | 28.864 (23.33) | 35.487 (33.70) | 35.035 (32.96) | 35.706 (34.07) |
| 1500 | 31.806 (27.78) | 31.071 (26.67) | 31.635 (27.52) | 32.273 (28.52) | 31.562 (27.41) | 35.255 (33.33) | 32.274 (28.52) | 30.849 (26.30) | 35.939 (34.44) | 35.035 (32.96) | 35.035 (32.96) |
| Control | 0.309 (00.0) | 0.309 (00.0) | 0.309 (00.0) | 0.309 (00.0) | 0.309 (00.0) | 0.309 (00.0) | 0.309 (00.0) | 0.309 (00.0) | 0.309 (00.0) | 0.309 (00.0) | 0.309 (00.0) |
| S.Em (±) | 0.59 | 0.67 | 0.51 | 0.52 | 0.66 | 0.55 | 0.57 | 0.57 | 0.62 | 0.50 | 0.43 |
| CD at 0.01 | 2.468 | 2.823 | 2.159 | 2.177 | 2.787 | 2.31 | 2.400 | 2.405 | 2.601 | 2.106 | 1.836 |
| CV (%) | 4.258 | 4.664 | 3.590 | 3.946 | 4.984 | 3.474 | 3.863 | 4.172 | 3.987 | 3.039 | 2.606 |
| Correlation | 0.96 | 0.93 | 0.92 | 0.96 | 0.94 | 0.88 | 0.98 | 0.93 | 0.97 | 0.81 | 0.78 |
| p-value | 0.001184 | 0.00358925 | 0.004672 | 0.001184 | 0.002646 | 0.010368 | 0.000298 | 0.00358925 | 0.00066825 | 0.02536025 | 0.033638 |
| 250 | 28.871 (23.33) | 27.334 (21.11) | 29.868 (24.83) | 31.455 (27.27) | 29.121 (23.70) | 28.873 (23.33) | 25.208 (18.15) | 27.603 (21.48) | 27.861 (21.85) | 26.017 (19.26) | 29.121 (23.70) |
| 500 | 30.845 (26.30) | 28.117 (22.22) | 31.321 (27.04) | 27.861 (21.85) | 30.605 (25.93) | 31.562 (27.41) | 29.121 (23.70 | 30.605 (25.93) | 29.377 (24.07) | 28.114 (22.22) | 32.741 (29.26) |
| 750 | 32.274 (28.52) | 29.357 (24.07) | 34.359 (31.85) | 36.825 (35.93) | 35.248 (33.34) | 34.807 (32.59) | 28.868 (23.33) | 34.807 (32.59) | 29.86 (24.81) | 28.627 (22.96) | 35.264 (33.33) |
| 1000 | 32.747 (29.26) | 31.564 (27.41) | 34.809 (32.59) | 35.699 (34.07) | 33.671 (30.74) | 34.809 (32.59) | 29.377 (24.07) | 34.809 (32.59) | 32.038 (28.15) | 29.868 (24.81) | 36.374 (35.19) |
| 1250 | 33.669 (30.74) | 32.274 (28.52) | 35.258 (33.33) | 37.041 (36.30) | 33.669 (30.74) | 35.036 (32.96) | 30.364 (25.56) | 35.264 (33.33) | 32.038 (28.15) | 31.088 (26.67) | 36.826 (35.93) |
| 1500 | 33.661 (30.74) | 33.62 (30.67) | 35.255 (33.33) | 37.933 (37.80) | 35.706 (34.07) | 35.255 (33.33) | 33.932 (31.19) | 35.704 (34.07) | 33.415 (30.33) | 33.475 (30.43) | 37.485 (37.04) |
| Control | 0.309 (00.0) | 0.309 (00.0) | 0.309 (00.0) | 0.309 (00.0) | 0.309 (00.0) | 0.309 (00.0) | 0.309 (00.0) | 0.309 (00.0) | 0.309 (00.0) | 0.309 (00.0) | 0.309 (00.0) |
| S.Em ( ± ) | 0.50 | 0.69 | 0.66 | 0.67 | 0.70 | 0.67 | 0.69 | 0.67 | 0.56 | 0.58 | 0.51 |
| CD at 0.01 | 2.108 | 2.907 | 2.783 | 2.814 | 2.948 | 2.807 | 2.906 | 2.813 | 2.342 | 2.420 | 2.152 |
| CV (%) | 3.153 | 4.589 | 3.988 | 3.910 | 4.285 | 4.023 | 4.710 | 4.064 | 3.658 | 3.942 | 2.989 |
| Correlation | 0.95 | 0.99 | 0.91 | 0.79 | 0.83 | 0.87 | 0.91 | 0.89 | 0.98 | 0.98 | 0.93 |
| p-value | 0.0018 | 7 E-05 | 0.0059 | 0.0308 | 0.0204 | 0.0121 | 0.0059 | 0.0087 | 0.0003 | 0.0003 | 0.0036 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sharma, J.; Manjunatha, N.; Pokhare, S.S.; Patil, P.G.; Agarrwal, R.; Chakranarayan, M.G.; Aralimar, A.; Devagire, P.; Marathe, R.A. Genetic Diversity and Streptomycin Sensitivity in Xanthomonas axonopodis pv. punicae Causing Oily Spot Disease in Pomegranates. Horticulturae 2022, 8, 441. https://doi.org/10.3390/horticulturae8050441
Sharma J, Manjunatha N, Pokhare SS, Patil PG, Agarrwal R, Chakranarayan MG, Aralimar A, Devagire P, Marathe RA. Genetic Diversity and Streptomycin Sensitivity in Xanthomonas axonopodis pv. punicae Causing Oily Spot Disease in Pomegranates. Horticulturae. 2022; 8(5):441. https://doi.org/10.3390/horticulturae8050441
Chicago/Turabian StyleSharma, Jyotsana, Nanjundappa Manjunatha, Somnath S. Pokhare, Prakash G. Patil, Ruchi Agarrwal, Mansi G. Chakranarayan, Anita Aralimar, Priya Devagire, and Rajiv A. Marathe. 2022. "Genetic Diversity and Streptomycin Sensitivity in Xanthomonas axonopodis pv. punicae Causing Oily Spot Disease in Pomegranates" Horticulturae 8, no. 5: 441. https://doi.org/10.3390/horticulturae8050441