
Anthocyanin Accumulation and Differential Expression of the Biosynthetic Genes Result in a Discrepancy in the Red Color of Herbaceous Peony (Paeonia lactiflora Pall.) Flowers

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Measurement of the Morphological Parameters
2.3. Measurements of the Floral Quality and Color Indices
2.4. Analysis of the Anthocyanins, Flavones, and Flavonols
2.5. Gene Expression Analysis
2.6. Statistical Analysis
3. Results
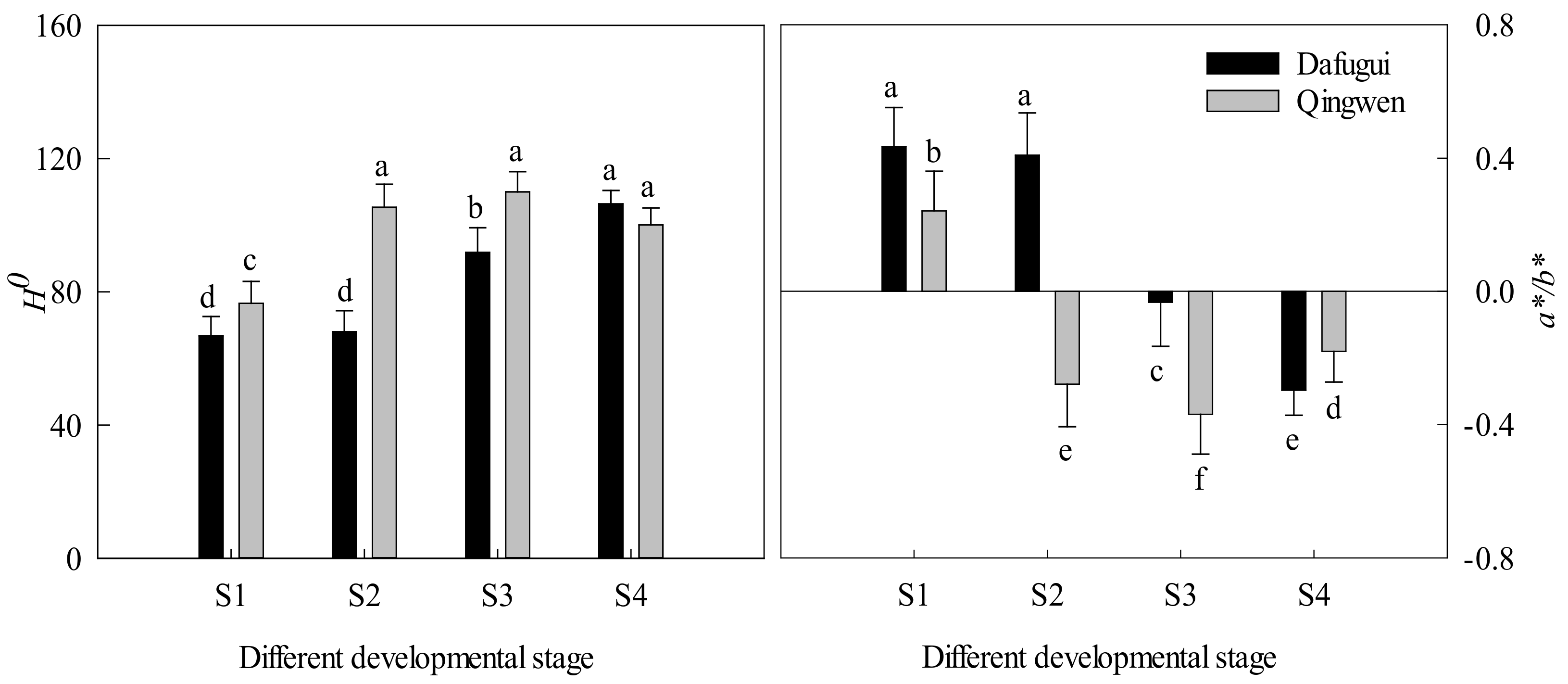
3.1. Flower Quality and Color Indices
3.2. Analysis of the Anthocyanins, Flavones, and Flavonols
3.3. Anthocyanin Biosynthetic Genes Expression Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yan, Z.G.; Li, M.C.; Xie, L.H.; Luo, X.N.; Yang, W.Z.; Yuan, Y.P.; Zhang, Y.L. A systematic comparison of 17 cultivated herbaceous peony seed based on phytochemicals and antioxidant activity. Eur. Food. Res. Technol. 2020, 246, 1919–1932. [Google Scholar] [CrossRef]
- Wang, J.G.; Zhang, Z.S. Herbaceous Peonies of China; China Forestry Publishing House: Beijing, China, 2005. [Google Scholar]
- Zhao, D.; Cheng, M.; Tang, W.; Liu, D.; Zhou, S.; Meng, J.; Tao, J. Nano-silver modifies the vase life of cut herbaceous peony (Paeonia lactiflora Pall.) flowers. Protoplasma 2018, 255, 1001–1013. [Google Scholar] [CrossRef] [PubMed]
- Wei, D.X.; Tang, Z.J.; da Silva, T.; Yu, X.N. In vitro induction of polyploidy by colchicine treatment in herbaceous peony (Paeonia lactiflora Pall.). Acta Hortic. 2017, 1171, 265–278. [Google Scholar] [CrossRef]
- Wan, Y.L.; Zhang, M.; Hong, A.Y.; Zhang, Y.X.; Liu, Y. Characteristics of microsatellites mined from transcriptome data and the development of novel markers in Paeonia lactiflora. Genes 2020, 11, 214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, D.J.; Zhang, R.J.; Sun, L.; Wu, Y.Q.; Li, C.D.; Xiang, L.T.; Zhao, Y.X. A Cultivation experiment of introduced cut lily cultivars in Shanghai area. J. Shanghai Jiao Tong Univ. (Agric. Sci.) 2016, 34, 65–83. (In Chinese) [Google Scholar] [CrossRef]
- Min, J.; Xiang, Y.C.; Tian, D.K. Consumption Demand and market potential of cut lotus flower in China. J. Anhui Agric. Sci. 2018, 46, 212–217. (In Chinese) [Google Scholar] [CrossRef]
- Elliot, A.J.; Maier, M.A. Color Psychology: Effects of Perceiving Color on Psychological Functioning in Humans. Annu. Rev. Psychol. 2014, 65, 95–120. [Google Scholar] [CrossRef]
- Tanaka, Y.; Brugliera, F.; Kalc, G.; Senior, M.; Dyson, B.; Nakamura, N.; Katsumoto, Y.; Chandler, S. Flower color modification by engineering of the flavonoid biosynthetic pathway: Practical perspectives. Biosci. Biotechnol. Biochem. 2010, 74, 1760–1769. [Google Scholar] [CrossRef] [Green Version]
- Mol, J.; Grotewold, E.; Koes, R. How genes paint flowers and seeds. Trends Plant Sci. 1998, 3, 212–217. [Google Scholar] [CrossRef]
- Khoo, H.E.; Azlan, A.; Tang, S.T.; Lim, S.M. Anthocyanidins and anthocyanins: Colored pigments as food, pharmaceutical ingredients, and the potential health benefits. Food Nutr. Res. 2017, 61, 136–1779. [Google Scholar] [CrossRef] [Green Version]
- Forkmann, G. Flavonoids as flower pigments: The formation of the natural spectrum and its extension by genetic engineering. Plant Breed. 1991, 106, 1–26. [Google Scholar] [CrossRef]
- Kong, J.M.; Chia, L.S.; Goh, N.K.; Chia, T.F.; Brouillard, R. Analysis and biological activities of anthocyanins. Phytochemistry 2003, 64, 923–933. [Google Scholar] [CrossRef]
- Mizuta, D.; Ban, T.; Miyajima, I.; Nakatsuka, A.; Kobayashi, N. Comparison of flower color with anthocyanin composition patterns in evergreen azalea. Sci. Hortic. 2009, 122, 594–602. [Google Scholar] [CrossRef]
- Tatsuzawa, F.; Saito, N.; Toki, K.; Shinoda, K.; Honda, T. Flower colors and their anthocyanins in Matthiola incana cultivars (Brassicaceae). J. Jpn. Soc. Hortic. Sci. 2012, 81, 91–100. [Google Scholar] [CrossRef] [Green Version]
- Zhu, M.; Zheng, X.; Shu, Q.; Li, H.; Zhong, P.; Zhang, H.; Xu, Y.; Wang, L.; Wang, L. Relationship between the composition of flavonoids and flower colors variation in tropical water lily (Nymphaea) cultivars. PLoS ONE 2012, 7, e34335. [Google Scholar] [CrossRef] [Green Version]
- Jia, N.; Shu, Q.Y.; Wang, L.S.; Du, H.; Xu, Y.J.; Liu, Z.A. Analysis of petal anthocyanins to investigate coloration mechanism in herbaceous peony cultivars. Sci. Hortic. 2008, 117, 167–173. [Google Scholar] [CrossRef]
- Zhao, D.Q.; Tao, J. Recent advances on the development and regulation of flower color in ornamental plants. Front. Plant Sci. 2015, 6, 261. [Google Scholar] [CrossRef] [Green Version]
- Filyushin, M.A.; Dzhos, E.A.; Shchennikova, A.V.; Kochieva, E.Z. Dependence of pepper fruit colour on basic pigments ratio and expression pattern of carotenoid and anthocyanin biosynthesis genes. Russ. J. Plant Physl. 2020, 67, 1054–1062. [Google Scholar] [CrossRef]
- Hossain, M.R.; Kim, H.T.; Shanmugam, A.; Nath, U.K.; Goswami, G.; Song, J.Y.; Park, J.I.; Nou, I.S. Expression profiling of regulatory and biosynthetic genes in contrastingly anthocyanin rich strawberry (Fragaria x ananassa) cultivars reveals key genetic determinants of fruit color. Int. J. Mol. Sci. 2018, 19, 656. [Google Scholar] [CrossRef] [Green Version]
- Karanjalker, G.R.; Ravishankar, K.V.; Shivashankara, K.S.; Dinesh, M.R.; Roy, T.K.; Rao, D.V.S. A study on the expression of genes involved in carotenoids and anthocyanins during ripening in fruit peel of green, yellow, and red colored mango cultivars. Appl. Biochem. Biotechnol. 2018, 184, 140–154. [Google Scholar] [CrossRef]
- Maritim, T.K.; Masand, M.; Seth, R.; Sharma, R.K. Transcriptional analysis reveals key insights into seasonal induced anthocyanin degradation and leaf color transition in purple tea (Camellia sinensis (L.) O. Kuntze). Sci. Rep. 2021, 11, 1244. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.H.; Fang, Z.W.; Liu, M.; Zhao, D.Q.; Jun, T. Color characteristics, pigment accumulation and biosynthetic analyses of leaf color variation in herbaceous peony (Paeonia lactifora Pall.). 3 Biotech 2020, 10, 76. [Google Scholar] [CrossRef]
- Kanemakia, A.; Otania, M.; Takanoa, M.; Fujimotoa, T.; Okuharab, H.; Nomizub, T.; Kondob, M.; Kobayashib, H.; Tatsuzawac, F.; Nakano, M. Ectopic expression of the R2R3-MYB gene from Tricyrtis sp. results in leaf color alteration in transgenic Pelargonium crispum. Sci. Hortic. 2018, 240, 411–416. [Google Scholar] [CrossRef]
- Zhao, D.; Tao, J.; Han, C.; Ge, J. Flower color diversity revealed by differential expression of flavonoid biosynthetic genes and flavonoid accumulation in herbaceous peony (Paeonia lactiflora Pall.). Mol. Biol. Rep. 2012, 39, 11263–11275. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.Q.; Sheng, Y.L.; Zhou, C.H. Method of non-destructive measurement research on leaf area and fruit fresh weight of Ginkgo biloba. North. Hortic. 2010, 7, 28–30. [Google Scholar] [CrossRef]
- McGuire, R.G. Reporting of objective color measurements. Hortscience 1992, 27, 1254–1255. [Google Scholar] [CrossRef] [Green Version]
- Voss, D.H. Relating colorimeter measurement of plant color to the Royal Horticultural Society Colour Chart. HortScience Publ. Am. Soc. Hortic. Sci. 1992, 27, 1256–1260. [Google Scholar] [CrossRef]
- Wang, L.S.; Shiraishi, A.; Hashimoto, F.; Aoki, N.; Sakata, Y. Analysis of petal anthocyanins to investigate flower coloration of Zhongyuan (Chinese) and Daikon Island (Japanese) tree peony cultivars. J. Plant Res. 2001, 114, 33–43. [Google Scholar] [CrossRef]
- Auger, B.; Marnet, N.; Gautier, V.; Maja-Grondard, A.; Leprine, F.; Renard, M.; Juyot, S.; Nesi, S.; Routaboul, J.M. A detailed survey of seed coat flavonoids in developing seeds of Brassica napus L. J. Agric. Food Chem. 2010, 58, 6246–6256. [Google Scholar] [CrossRef]
- Fan, J.; Zhu, W.; Kang, H.; Ma, H.; Tao, G. Flavonoid constituents and antioxidant capacity in flowers of different Zhongyuan tree penoy cultivars. J. Funct. Foods 2012, 4, 147–157. [Google Scholar] [CrossRef]
- Tuberoso, C.I.G.; Montoro, P.; Piacente, S.; Corona, G.; Deiana, M.; Dessi, M.A.; Pizza, C.; Cabras, P. Flavonoid characterization and antioxidant activity of hydroalcoholic extracts from Achillea ligustica All. J. Pharm. Biomed. 2009, 50, 440–448. [Google Scholar] [CrossRef]
- Villanueva, E.; Fujibayashi-Yoshii, N.; Matsuzaki, S.; Yamazaki, K.; Burana, C.; Yamane, K. Effects of trehalose and sucrose on the vase life and physiology of cut astilbe (Astilbe x arendsii Arends) flowers. Horticult. J. 2019, 88, 276–283. [Google Scholar] [CrossRef] [Green Version]
- Zhao, D.Q.; Tang, W.H.; Hao, Z.J.; Tao, J. Identification of flavonoids and expression of flavonoid biosynthetic genes in two coloured tree peony flowers. Biochem. Bioph. Res. Co. 2015, 459, 450–456. [Google Scholar] [CrossRef] [PubMed]
- Nakatsuka, T.; Nishihara, M.; Mishiba, K.; Yamamura, S. Temporal expression of flavonoid biosynthesis-related genes regulates flower pigmentation in gentian plants. Plant Sci. 2005, 168, 1309–1318. [Google Scholar] [CrossRef]
- Wang, N.; Liu, W.J.; Chen, X.S. Transcriptomic Analysis of red-fleshed apples reveals the novel role of MdWRKY11 in flavonoid and anthocyanin biosynthesis. J. Agric. Food Chem. 2018, 66, 7076–7086. [Google Scholar] [CrossRef]
- Niu, S.S.; Xu, C.J.; Zhang, W.S.; Zhang, B.; Li, X.; Lin-Wang, K.; Ferguson, I.B.; Allan, A.C.; Chen, K.S. Coordinated regulation of anthocyanin biosynthesis in chinese bayberry (Myrica rubra) fruit by a R2R3 Myb transcription factor. Planta 2010, 231, 887–899. [Google Scholar] [CrossRef]
- Kobayashi, S.; Ishimaru, M.; Ding, C.K.; Yakushiji, H.; Goto, N. Comparison of udp-glucose: Flavonoid 3-o-glucosyltransferase (UFGT) gene sequences between white grapes (Vitis vinifera) and their sports with red skin. Plant Sci. 2001, 160, 543–550. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak | tR (min) | λmax (nm) | MS1 (m/z) | MS2 (m/z) | Identification |
|---|---|---|---|---|---|
| a1 | 3.01 | 235, 275, 515 | 611.2 | 448.9, 287.2 | Cyanidin-3,5-di-O-glucoside [17,29] |
| a2 | 4.80 | 275, 510 | 625.2 | 462.9, 301.1 | Peonidin-3,5-di-O-glucoside [17] |
| f1 | 7.31 | 235, 315 | 627.0 | 464.9, 303.3 | Quercetin di-hexoside [30] |
| f2 | 10.32 | 265, 345 | 611.1 | 449.1, 287.3 | Kaempferol di-hexoside [31] |
| f3 | 11.57 | 255, 355 | 641.0 | 478.9, 317.0 | Isorhamnetin di-hexoside [31] |
| f4 | 21.75 | 235, 370 | 465.1 | 303.2 | Quercetin-3-O-rhamnoside [32] |
| f5 | 29.19 | 230, 265, 365 | 449.1 | 287.2 | Luteolin-7-O-glucoside [32] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Y.; Hao, Z.; Tang, Y.; Zhao, D. Anthocyanin Accumulation and Differential Expression of the Biosynthetic Genes Result in a Discrepancy in the Red Color of Herbaceous Peony (Paeonia lactiflora Pall.) Flowers. Horticulturae 2022, 8, 349. https://doi.org/10.3390/horticulturae8040349
Wu Y, Hao Z, Tang Y, Zhao D. Anthocyanin Accumulation and Differential Expression of the Biosynthetic Genes Result in a Discrepancy in the Red Color of Herbaceous Peony (Paeonia lactiflora Pall.) Flowers. Horticulturae. 2022; 8(4):349. https://doi.org/10.3390/horticulturae8040349
Chicago/Turabian StyleWu, Yanqing, Zhaojun Hao, Yuhan Tang, and Daqiu Zhao. 2022. "Anthocyanin Accumulation and Differential Expression of the Biosynthetic Genes Result in a Discrepancy in the Red Color of Herbaceous Peony (Paeonia lactiflora Pall.) Flowers" Horticulturae 8, no. 4: 349. https://doi.org/10.3390/horticulturae8040349