
Impact of Pseudomonas sp. SVB-B33 on Stress- and Cell Wall-Related Genes in Roots and Leaves of Hemp under Salinity


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Isolation, Phylogenetic Analysis and Growth
2.2. Silicate Solubilizing Assay
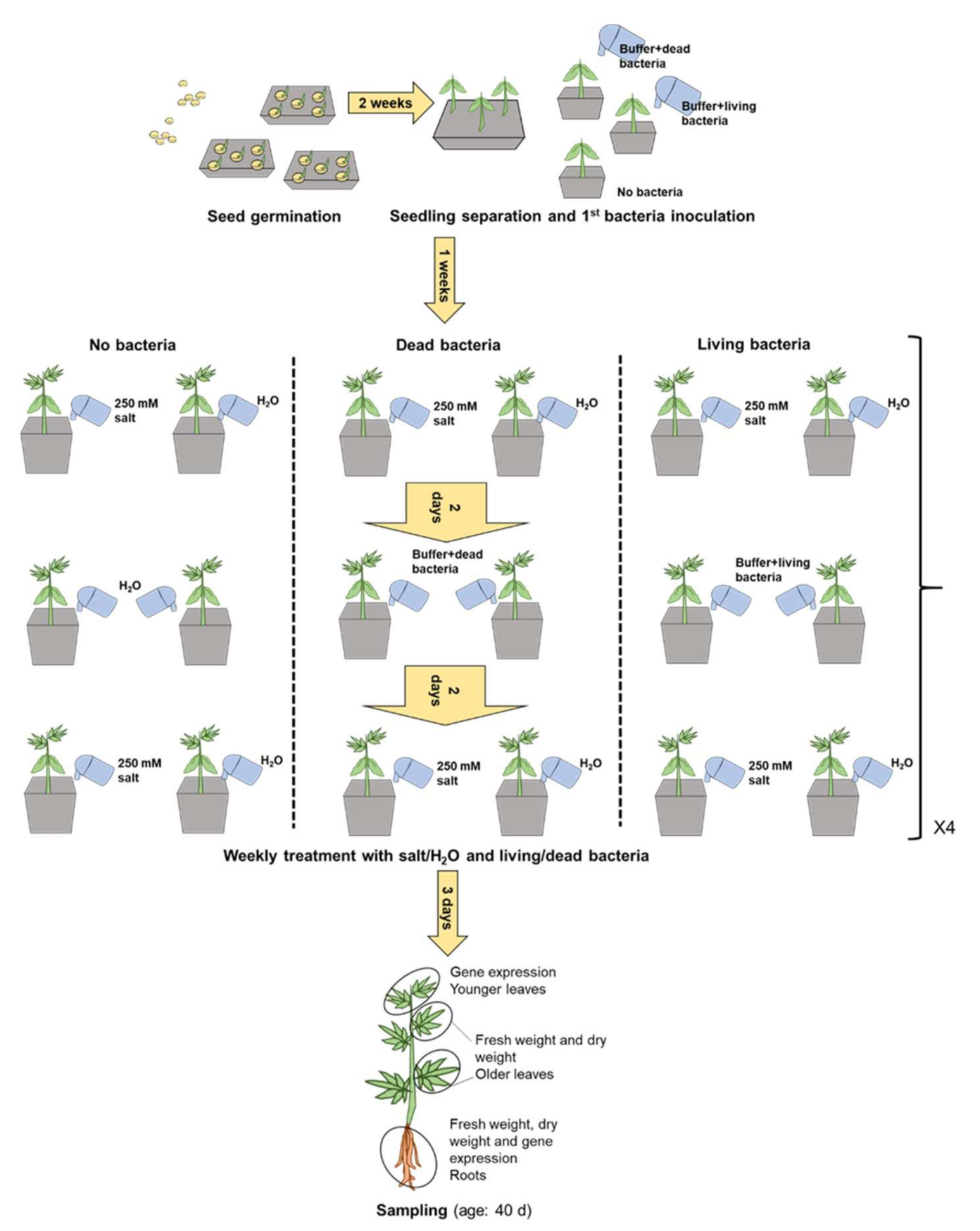
2.3. Plant Growth and Treatments
2.4. RNA Extraction, cDNA Synthesis, qPCR and Statistical Analyses
3. Results
3.1. Phylogenetic Analysis and Silicate Solubilizing Activity
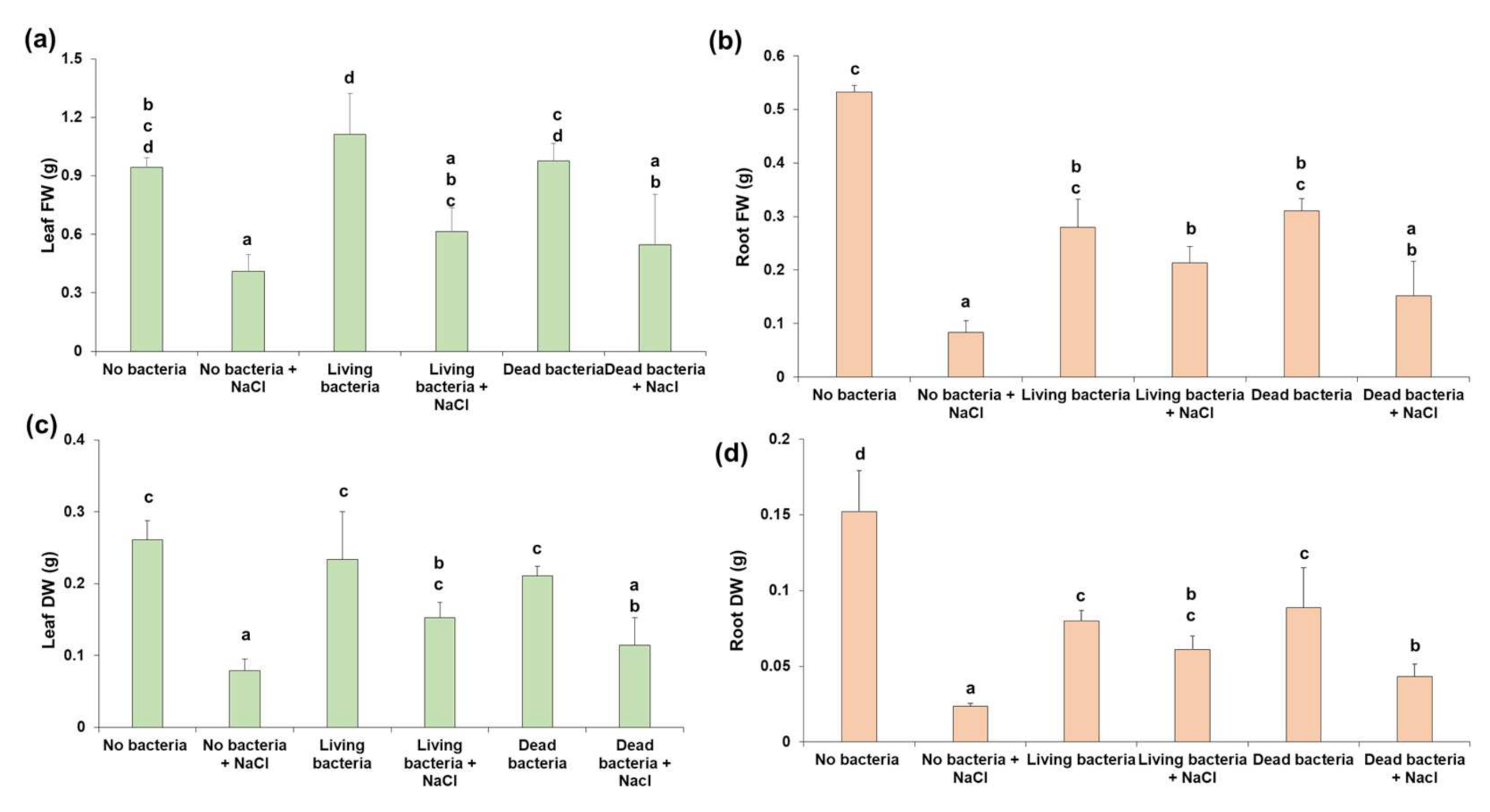
3.2. The Application of Living Bacteria to Hemp Increases the Root Biomass under Salinity
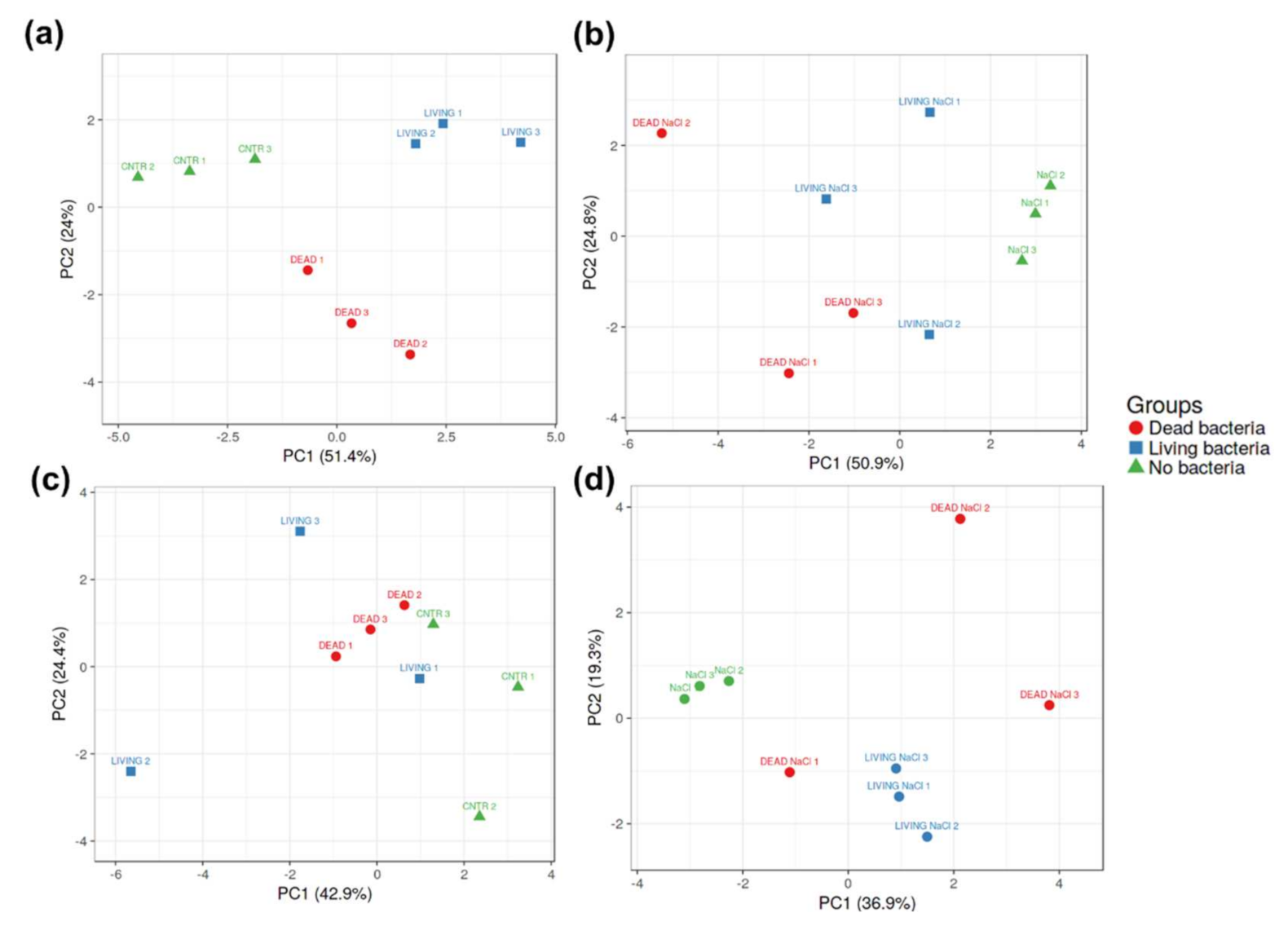
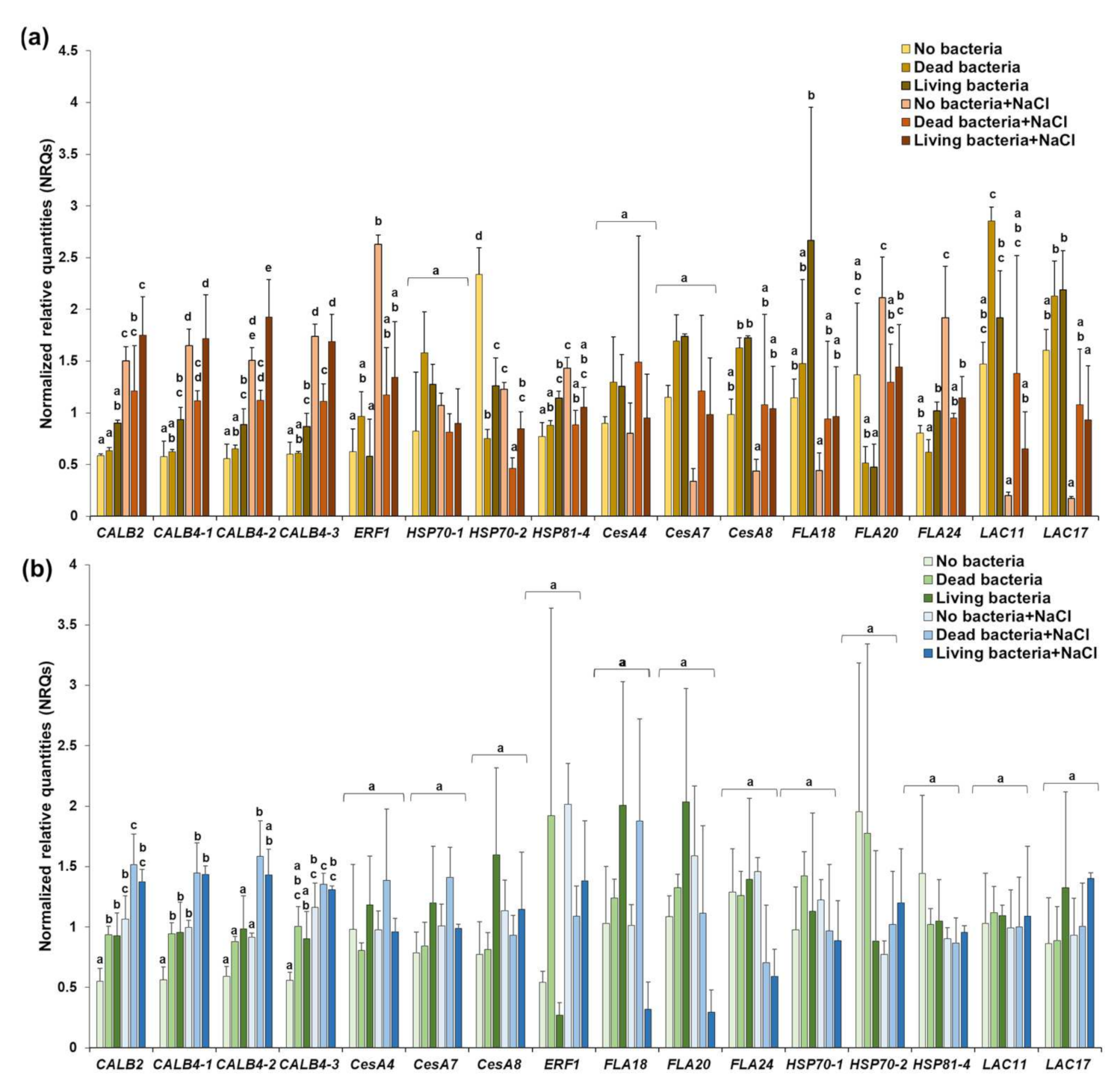
3.3. The Inoculation with Pseudomonas sp. SVB-B33 Affects Gene Expression in Hemp Roots and Leaves
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Beres, B.L.; Hatfield, J.L.; Kirkegaard, J.A.; Eigenbrode, S.D.; Pan, W.L.; Lollato, R.P.; Hunt, J.R.; Strydhorst, S.; Porker, K.; Lyon, D.; et al. Toward a Better Understanding of Genotype × Environment × Management Interactions—A Global Wheat Initiative Agronomic Research Strategy. Front. Plant Sci. 2020, 11, 828. [Google Scholar] [CrossRef] [PubMed]
- Studnicki, M.; Wijata, M.; Sobczyński, G.; Samborski, S.; Gozdowski, D.; Rozbicki, J. Effect of Genotype, Environment and Crop Management on Yield and Quality Traits in Spring Wheat. J. Cereal Sci. 2016, 72, 30–37. [Google Scholar] [CrossRef]
- Long, S.P.; Zhu, X.-G.; Naidu, S.L.; Ort, D.R. Can Improvement in Photosynthesis Increase Crop Yields? Plant Cell Environ. 2006, 29, 315–330. [Google Scholar] [CrossRef] [PubMed]
- Keller, B.; Zimmermann, L.; Rascher, U.; Matsubara, S.; Steier, A.; Muller, O. Toward Predicting Photosynthetic Efficiency and Biomass Gain in Crop Genotypes over a Field Season. Plant Physiol. 2022, 188, 301–317. [Google Scholar] [CrossRef]
- Ciura, J.; Kruk, J. Phytohormones as Targets for Improving Plant Productivity and Stress Tolerance. J. Plant Physiol. 2018, 229, 32–40. [Google Scholar] [CrossRef]
- Fahad, S.; Bajwa, A.A.; Nazir, U.; Anjum, S.A.; Farooq, A.; Zohaib, A.; Sadia, S.; Nasim, W.; Adkins, S.; Saud, S.; et al. Crop Production under Drought and Heat Stress: Plant Responses and Management Options. Front. Plant Sci. 2017, 8, 1147. [Google Scholar] [CrossRef] [Green Version]
- Pereira, C.S.; Lopes, I.; Abrantes, I.; Sousa, J.P.; Chelinho, S. Salinization Effects on Coastal Ecosystems: A Terrestrial Model Ecosystem Approach. Philos. Trans. R. Soc. B Biol. Sci. 2019, 374, 20180251. [Google Scholar] [CrossRef] [Green Version]
- Hu, H.; Liu, H.; Liu, F. Seed Germination of Hemp (Cannabis sativa L.) Cultivars Responds Differently to the Stress of Salt Type and Concentration. Ind. Crops Prod. 2018, 123, 254–261. [Google Scholar] [CrossRef]
- Guerriero, G.; Behr, M.; Hausman, J.-F.; Legay, S. Textile Hemp vs. Salinity: Insights from a Targeted Gene Expression Analysis. Genes 2017, 8, 242. [Google Scholar] [CrossRef] [Green Version]
- Di Mola, I.; Conti, S.; Cozzolino, E.; Melchionna, G.; Ottaiano, L.; Testa, A.; Sabatino, L.; Rouphael, Y.; Mori, M. Plant-Based Protein Hydrolysate Improves Salinity Tolerance in Hemp: Agronomical and Physiological Aspects. Agronomy 2021, 11, 342. [Google Scholar] [CrossRef]
- Berni, R.; Mandlik, R.; Hausman, J.-F.; Guerriero, G. Silicon-Induced Mitigatory Effects in Salt-Stressed Hemp Leaves. Physiol. Plant. 2021, 171, 476–482. [Google Scholar] [CrossRef] [PubMed]
- Guerriero, G.; Deshmukh, R.; Sonah, H.; Sergeant, K.; Hausman, J.-F.; Lentzen, E.; Valle, N.; Siddiqui, K.S.; Exley, C. Identification of the Aquaporin Gene Family in Cannabis sativa and Evidence for the Accumulation of Silicon in Its Tissues. Plant Sci. 2019, 287, 110167. [Google Scholar] [CrossRef] [PubMed]
- Guerriero, G.; Sutera, F.M.; Torabi-Pour, N.; Renaut, J.; Hausman, J.-F.; Berni, R.; Pennington, H.C.; Welsh, M.; Dehsorkhi, A.; Zancan, L.R.; et al. Phyto-Courier, a Silicon Particle-Based Nano-Biostimulant: Evidence from Cannabis sativa Exposed to Salinity. ACS Nano 2021, 15, 3061–3069. [Google Scholar] [CrossRef] [PubMed]
- Orozco-Mosqueda, M.d.C.; Flores, A.; Rojas-Sánchez, B.; Urtis-Flores, C.A.; Morales-Cedeño, L.R.; Valencia-Marin, M.F.; Chávez-Avila, S.; Rojas-Solis, D.; Santoyo, G. Plant Growth-Promoting Bacteria as Bioinoculants: Attributes and Challenges for Sustainable Crop Improvement. Agronomy 2021, 11, 1167. [Google Scholar] [CrossRef]
- Berni, R.; Guerriero, G.; Cai, G. One for All and All for One! Increased Plant Heavy Metal Tolerance by Growth-Promoting Microbes: A Metabolomics Standpoint. In Plant Metallomics and Functional Omics: A System-Wide Perspective; Sablok, G., Ed.; Springer International Publishing: Cham, Switzerland, 2019; pp. 39–54. ISBN 978-3-030-19103-0. [Google Scholar]
- Backes, A.; Charton, S.; Planchon, S.; Esmaeel, Q.; Sergeant, K.; Hausman, J.-F.; Renaut, J.; Barka, E.A.; Jacquard, C.; Guerriero, G. Gene Expression and Metabolite Analysis in Barley Inoculated with Net Blotch Fungus and Plant Growth-Promoting Rhizobacteria. Plant Physiol. Biochem. 2021, 168, 488–500. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, P.; Kanagendran, A.; Samaddar, S.; Pazouki, L.; Sa, T.-M.; Niinemets, Ü. Inoculation of Brevibacterium linens RS16 in Oryza sativa Genotypes Enhanced Salinity Resistance: Impacts on Photosynthetic Traits and Foliar Volatile Emissions. Sci. Total Environ. 2018, 645, 721–732. [Google Scholar] [CrossRef]
- Khan, M.A.; Asaf, S.; Khan, A.L.; Jan, R.; Kang, S.-M.; Kim, K.-M.; Lee, I.-J. Thermotolerance Effect of Plant Growth-Promoting Bacillus cereus SA1 on Soybean during Heat Stress. BMC Microbiol. 2020, 20, 175. [Google Scholar] [CrossRef]
- Ren, X.-M.; Guo, S.-J.; Tian, W.; Chen, Y.; Han, H.; Chen, E.; Li, B.-L.; Li, Y.-Y.; Chen, Z.-J. Effects of Plant Growth-Promoting Bacteria (PGPB) Inoculation on the Growth, Antioxidant Activity, Cu Uptake, and Bacterial Community Structure of Rape (Brassica napus L.) Grown in Cu-Contaminated Agricultural Soil. Front. Microbiol. 2019, 10, 1455. [Google Scholar] [CrossRef]
- Yu, Y.; Gui, Y.; Li, Z.; Jiang, C.; Guo, J.; Niu, D. Induced Systemic Resistance for Improving Plant Immunity by Beneficial Microbes. Plants 2022, 11, 386. [Google Scholar] [CrossRef]
- Lee, K.E.; Kang, S.M.; Adhikari, A.; Lee, I.J. Effect of Silicate Solubilizing Bacteria Pseudomonas psychrotolerans CS51 Treatment on Soybean Crops at Paddy Soil. In Proceedings of the Korean Society of Crop Science Conference; The Korean Society of Crop Science: Suwon, Korea, 2019; p. 46. [Google Scholar]
- Kubi, H.A.A.; Khan, M.A.; Adhikari, A.; Imran, M.; Kang, S.-M.; Hamayun, M.; Lee, I.-J. Silicon and Plant Growth-Promoting Rhizobacteria Pseudomonas psychrotolerans CS51 Mitigates Salt Stress in Zea mays L. Agriculture 2021, 11, 272. [Google Scholar] [CrossRef]
- Lozano, J.C.; Sbqueira, L. Prevention of the hypersensitive reaction in Tobacco leaves by heat-killed bacterial cells. Phytopathology 1970, 60, 875–879. [Google Scholar] [CrossRef]
- Rico, A.; Bennett, M.H.; Forcat, S.; Huang, W.E.; Preston, G.M. Agroinfiltration Reduces ABA Levels and Suppresses Pseudomonas Syringae-Elicited Salicylic Acid Production in Nicotiana tabacum. PLoS ONE 2010, 5, e8977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Gall, H.; Philippe, F.; Domon, J.-M.; Gillet, F.; Pelloux, J.; Rayon, C. Cell Wall Metabolism in Response to Abiotic Stress. Plants 2015, 4, 112–166. [Google Scholar] [CrossRef] [PubMed]
- Tenhaken, R. Cell Wall Remodeling under Abiotic Stress. Front. Plant Sci. 2015, 5. [Google Scholar] [CrossRef] [Green Version]
- Mignard, S.; Flandrois, J.P. 16S rRNA Sequencing in Routine Bacterial Identification: A 30-Month Experiment. J. Microbiol. Methods 2006, 67, 574–581. [Google Scholar] [CrossRef]
- Widmer, F.; Seidler, R.J.; Gillevet, P.M.; Watrud, L.S.; Di Giovanni, G.D. A Highly Selective PCR Protocol for Detecting 16S rRNA Genes of the Genus Pseudomonas (Sensu Stricto) in Environmental Samples. Appl. Environ. Microbiol. 1998, 64, 2545–2553. [Google Scholar] [CrossRef] [Green Version]
- McWilliam, H.; Li, W.; Uludag, M.; Squizzato, S.; Park, Y.M.; Buso, N.; Cowley, A.P.; Lopez, R. Analysis Tool Web Services from the EMBL-EBI. Nucleic Acids Res. 2013, 41, W597–W600. [Google Scholar] [CrossRef] [Green Version]
- Trifinopoulos, J.; Nguyen, L.-T.; von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A Fast Online Phylogenetic Tool for Maximum Likelihood Analysis. Nucleic Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef] [Green Version]
- Available online: https://www.dsmz.de/microorganisms/medium/pdf/DSMZ_Medium12.pdf (accessed on 15 March 2022).
- Vasanthi, N.; Saleena, L.M.; Raj, S.A. Silica Solubilization Potential of Certain Bacterial Species in the Presence of Different Silicate Minerals. Silicon 2018, 10, 267–275. [Google Scholar] [CrossRef]
- Guerriero, G.; Mangeot-Peter, L.; Legay, S.; Behr, M.; Lutts, S.; Siddiqui, K.S.; Hausman, J.-F. Identification of Fasciclin-like Arabinogalactan Proteins in Textile Hemp (Cannabis sativa L.): In Silico Analyses and Gene Expression Patterns in Different Tissues. BMC Genom. 2017, 18, 741. [Google Scholar] [CrossRef] [Green Version]
- Behr, M.; Sergeant, K.; Leclercq, C.C.; Planchon, S.; Guignard, C.; Lenouvel, A.; Renaut, J.; Hausman, J.-F.; Lutts, S.; Guerriero, G. Insights into the Molecular Regulation of Monolignol-Derived Product Biosynthesis in the Growing Hemp Hypocotyl. BMC Plant Biol. 2018, 18, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hellemans, J.; Mortier, G.; De Paepe, A.; Speleman, F.; Vandesompele, J. Open Access Method QBase Relative Quantification Framework and Software for Management and Automated Analysis of Real-Time Quantitative PCR Data. Genome Biol. 2007, 81, R19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-Aguilera, K.L.; Saad, C.F.; Chávez Montes, R.A.; Alves-Ferreira, M.; de Folter, S. Selection of Reference Genes for Quantitative Real-Time RT-PCR Studies in Tomato Fruit of the Genotype MT-Rg1. Front. Plant Sci. 2016, 7, 1386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Metsalu, T.; Vilo, J. ClustVis: A Web Tool for Visualizing Clustering of Multivariate Data Using Principal Component Analysis and Heatmap. Nucleic Acids Res. 2015, 43, W566–W570. [Google Scholar] [CrossRef] [PubMed]
- Fadiji, A.E.; Babalola, O.O.; Santoyo, G.; Perazzolli, M. The Potential Role of Microbial Biostimulants in the Amelioration of Climate Change-Associated Abiotic Stresses on Crops. Front. Microbiol. 2022, 12, 829099. [Google Scholar] [CrossRef] [PubMed]
- Andre, C.M.; Hausman, J.-F.; Guerriero, G. Cannabis sativa: The Plant of the Thousand and One Molecules. Front. Plant Sci. 2016, 7, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pellegrini, M.; Ercole, C.; Gianchino, C.; Bernardi, M.; Pace, L.; Del Gallo, M. Fusarium oxysporum f. Sp. Cannabis Isolated from Cannabis sativa L.: In Vitro and In Planta Biocontrol by a Plant Growth Promoting-Bacteria Consortium. Plants 2021, 10, 2436. [Google Scholar] [CrossRef]
- Pagnani, G.; Pellegrini, M.; Galieni, A.; D’Egidio, S.; Matteucci, F.; Ricci, A.; Stagnari, F.; Sergi, M.; Lo Sterzo, C.; Pisante, M.; et al. Plant Growth-Promoting Rhizobacteria (PGPR) in Cannabis sativa ‘Finola’ Cultivation: An Alternative Fertilization Strategy to Improve Plant Growth and Quality Characteristics. Ind. Crops Prod. 2018, 123, 75–83. [Google Scholar] [CrossRef]
- Lyu, D.; Backer, R.; Robinson, W.G.; Smith, D.L. Plant Growth-Promoting Rhizobacteria for Cannabis Production: Yield, Cannabinoid Profile and Disease Resistance. Front. Microbiol. 2019, 10, 1761. [Google Scholar] [CrossRef] [Green Version]
- Scott, M.; Rani, M.; Samsatly, J.; Charron, J.-B.; Jabaji, S. Endophytes of Industrial Hemp (Cannabis sativa L.) Cultivars: Identification of Culturable Bacteria and Fungi in Leaves, Petioles, and Seeds. Can. J. Microbiol. 2018, 64, 10. [Google Scholar] [CrossRef] [Green Version]
- Kang, S.-M.; Asaf, S.; Khan, A.L.; Lubna; Khan, A.; Mun, B.-G.; Khan, M.A.; Gul, H.; Lee, I.-J. Complete Genome Sequence of Pseudomonas psychrotolerans CS51, a Plant Growth-Promoting Bacterium, Under Heavy Metal Stress Conditions. Microorganisms 2020, 8, 382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Portieles, R.; Xu, H.; Yue, Q.; Zhao, L.; Zhang, D.; Du, L.; Gao, X.; Gao, J.; Portal Gonzalez, N.; Santos Bermudez, R.; et al. Heat-Killed Endophytic Bacterium Induces Robust Plant Defense Responses against Important Pathogens. Sci. Rep. 2021, 11, 12182. [Google Scholar] [CrossRef] [PubMed]
- Arkhipova, T.N.; Prinsen, E.; Veselov, S.U.; Martinenko, E.V.; Melentiev, A.I.; Kudoyarova, G.R. Cytokinin Producing Bacteria Enhance Plant Growth in Drying Soil. Plant Soil 2007, 292, 305–315. [Google Scholar] [CrossRef]
- Taverniti, V.; Guglielmetti, S. The Immunomodulatory Properties of Probiotic Microorganisms beyond Their Viability (Ghost Probiotics: Proposal of Paraprobiotic Concept). Genes Nutr. 2011, 6, 261–274. [Google Scholar] [CrossRef] [Green Version]
- Morcillo, R.J.L.; Vílchez, J.I.; Zhang, S.; Kaushal, R.; He, D.; Zi, H.; Liu, R.; Niehaus, K.; Handa, A.K.; Zhang, H. Plant Transcriptome Reprograming and Bacterial Extracellular Metabolites Underlying Tomato Drought Resistance Triggered by a Beneficial Soil Bacteria. Metabolites 2021, 11, 369. [Google Scholar] [CrossRef]
- Chun, H.J.; Baek, D.; Cho, H.M.; Lee, S.H.; Jin, B.J.; Yun, D.-J.; Hong, Y.-S.; Kim, M.C. Lignin Biosynthesis Genes Play Critical Roles in the Adaptation of Arabidopsis Plants to High-Salt Stress. Plant Signal. Behav. 2019, 14, 1625697. [Google Scholar] [CrossRef]
- Guerriero, G.; Stokes, I.; Valle, N.; Hausman, J.-F.; Exley, C. Visualising Silicon in Plants: Histochemistry, Silica Sculptures and Elemental Imaging. Cells 2020, 9, 1066. [Google Scholar] [CrossRef] [Green Version]
- Luyckx, M.; Hausman, J.-F.; Sergeant, K.; Guerriero, G.; Lutts, S. Molecular and Biochemical Insights Into Early Responses of Hemp to Cd and Zn Exposure and the Potential Effect of Si on Stress Response. Front. Plant Sci. 2021, 12, 711853. [Google Scholar] [CrossRef]
- Luyckx, M.; Hausman, J.-F.; Isenborghs, A.; Guerriero, G.; Lutts, S. Impact of Cadmium and Zinc on Proteins and Cell Wall-Related Gene Expression in Young Stems of Hemp (Cannabis sativa L.) and Influence of Exogenous Silicon. Environ. Exp. Bot. 2021, 183, 104363. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Conditions | ECe (mS) ± SD | pH ± SD |
|---|---|---|
| No bacteria | 0.561 ± 0.05 a | 5.2 ± 0.04 a |
| No bacteria + NaCl | 2.532 ± 0.15 b | 5.3 ± 0.01 a |
| Living bacteria | 0.585 ± 0.09 a | 5.5 ± 0.06 a |
| Living bacteria + NaCl | 2.631 ± 0.44 b | 5.2 ± 0.03 a |
| Dead bacteria | 0.541 ± 0.04 a | 5.2 ± 0.04 a |
| Dead bacteria + NaCl | 2.398 ± 0.29 b | 5.6 ± 0.02 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Berni, R.; Hausman, J.-F.; Villas-Boas, S.; Guerriero, G. Impact of Pseudomonas sp. SVB-B33 on Stress- and Cell Wall-Related Genes in Roots and Leaves of Hemp under Salinity. Horticulturae 2022, 8, 336. https://doi.org/10.3390/horticulturae8040336
Berni R, Hausman J-F, Villas-Boas S, Guerriero G. Impact of Pseudomonas sp. SVB-B33 on Stress- and Cell Wall-Related Genes in Roots and Leaves of Hemp under Salinity. Horticulturae. 2022; 8(4):336. https://doi.org/10.3390/horticulturae8040336
Chicago/Turabian StyleBerni, Roberto, Jean-Francois Hausman, Silas Villas-Boas, and Gea Guerriero. 2022. "Impact of Pseudomonas sp. SVB-B33 on Stress- and Cell Wall-Related Genes in Roots and Leaves of Hemp under Salinity" Horticulturae 8, no. 4: 336. https://doi.org/10.3390/horticulturae8040336