
Potential Uses of Scallop Shell Powder as a Substrate for the Cultivation of King Oyster Mushroom (Pleurotus eryngii)

Abstract
:1. Introduction
2. Materials and Methods
2.1. Inoculum Source and Spawn Preparation
2.2. Preparation of Ca Powder (SSP)
2.3. Substrate Preparation
2.4. Assay for the Growth Rate
2.5. Mushroom Cultivation
2.6. Composition Analysis
2.7. Composition Statistics
3. Results
3.1. Growth Rate of Mycelia
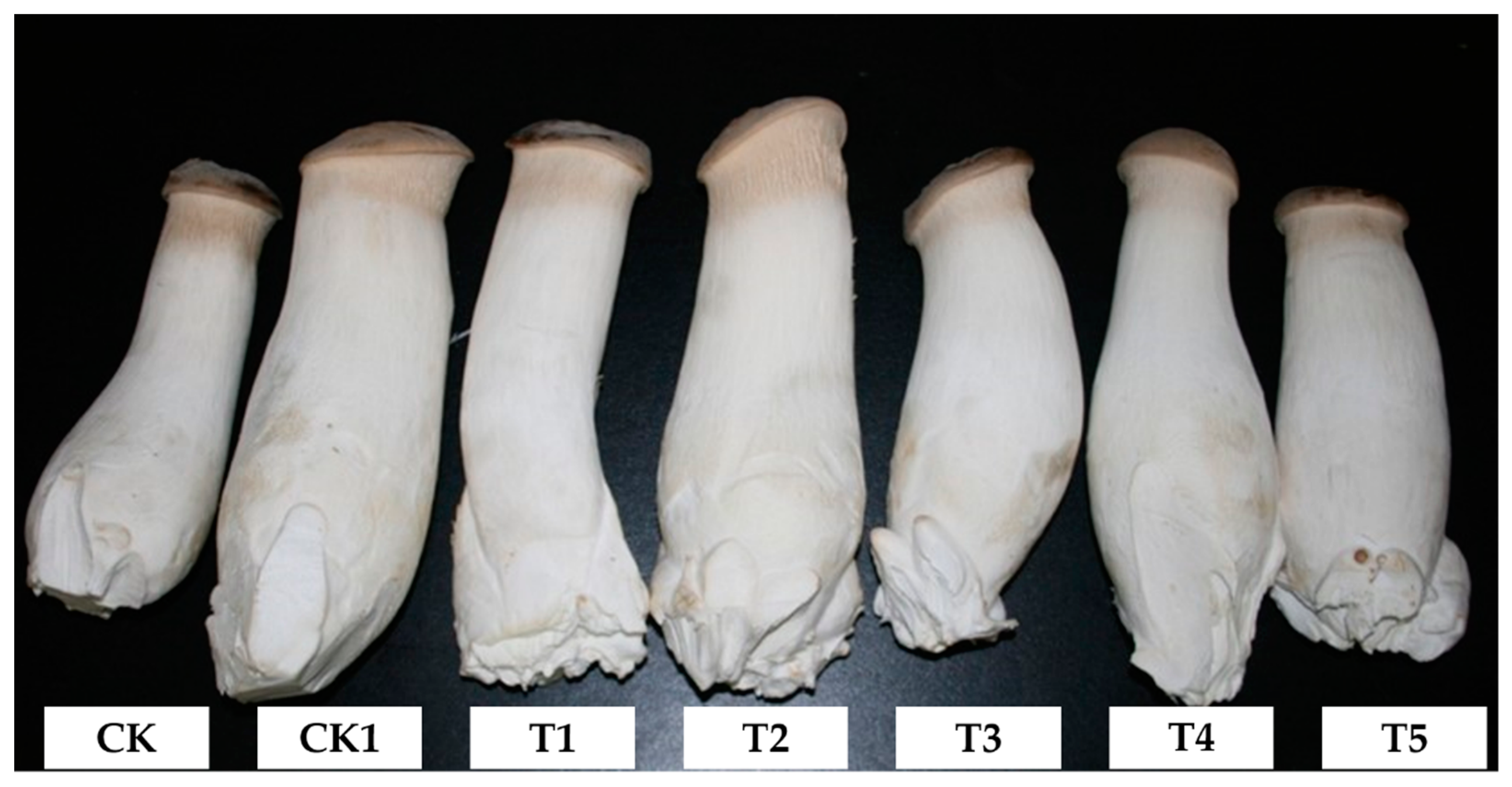
3.2. Characteristics of P. eryngii Fruiting Bodies on Different Substrates
3.3. Nutrient Content of Mushrooms
3.4. Macronutrient Elements of Mushrooms
3.5. Micronutrient Elements of Mushrooms
4. Discussion
4.1. Influence of SSP Supplementation on the Growth Rate of Mycelia
4.2. Influence of SSP Supplementation on the Morphological and Fruit Characteristics of P. eryngii
4.3. Influence of SSP Supplementation on the Nutrient Content of Mushrooms
4.4. Influence of SSP Supplementation on the Macronutrient Elements of Mushrooms
4.5. Influence of SSP Supplementation on the Micronutrient Elements of Mushrooms
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ministry of Agriculture and Rural Affairs, Fisheries Bureau. China Fishery Statistical Yearbook; China Agriculture Press: Beijing, China, 2020; p. 23.
- Wang, Y.H. Study on modification of shell powder and its application. Master’s Thesis, Dalian University of Technology, Dalian, China, 2016. [Google Scholar]
- Liu, P.J. Research on preparation and biological activity of marine shell nanomaterials. Master’s Thesis, Yantai University, Yantai, China, 2021. [Google Scholar]
- Dai, Y.P.; Wang, X.Y.; Ye, W.Z.; Fan, B.Y.; Ru, Y.; Xia, M.S.; Yao, Z.T. Study on resource utilization of mollusk shell waste. Resour. Dev. Mark. 2017, 33, 203–208. [Google Scholar]
- Jeznabadi, E.K.; Jafarpour, M.; Eghbalsaied, S. King oyster mushroom production using various sources of agricultural wastes in Iran. Int. J. Recvcl. Org. Waste Agricult. 2016, 5, 17–24. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Li, P.; Hu, H.; Wang, Y.; Sun, Y.; Huang, K. Postharvest decay of the cultivated mushroom Pleurotus eryngii caused by Lactococcus lactis subsp Lactis. J. Plant Pathol. 2013, 95, 247–253. [Google Scholar]
- Ma, G.X.; Yang, W.J.; Mariga, A.M.; Fang, Y.; Ma, N.; Pei, F.; Hu, Q.H. Purification, characterization and antitumor activity of polysaccharides from Pleurotus eryngii residue. Carbohydr. Polym. 2014, 114, 297–305. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Xu, J.; Fu, Q.; Fu, X.; Shu, T.; Bi, Y.; Song, B. Antitumor activity of a polysaccharide from Pleurotus eryngii on mice bearing renal cancer. Carbohydr. Polym. 2013, 95, 615–620. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.N.; Hu, X.L.; Li, W.X. Antioxidant, antitumor and immunostimulatory activities of the polypeptide from Pleurotus eryngii mycelium. Int. J. Biol. Macromol. 2017, 97, 323–330. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.J.; Du, F.; Zhang, H.J.; Hu, Q.X. Evaluation of korshinsk peashrub (Caragana korshinskii Kom) as a substrate for the cultivation of Pleurotus eryngii. Waste Biomass Valorization 2019, 10, 2879–2885. [Google Scholar] [CrossRef]
- Rodriguez Estrada, A.E.; Royse, D.J. Yield, size and bacterial blotch resistance of Pleurotus eryngii grown on cottonseed hulls/oak sawdust supplemented with manganese, copper and whole ground soybean. Bioresour. Technol. 2007, 98, 1898–1906. [Google Scholar] [CrossRef]
- Choi, U.K.; Bajpai, V.K.; Lee, N.H. Influence of calcinated starfish powder on growth, yield, spawn run and primordial germination of king oyster mushroom (Pleurotus eryngii). Food Chem. Toxicol. 2009, 47, 2830–2833. [Google Scholar] [CrossRef]
- Choi, U.K.; Li, O.H.; Kim, Y.C. Effect of calcinated oyster shell powder on growth, yield, spawn run, and primordial formation of king oyster mushroom (Pleurotus eryngii). Molecules 2011, 16, 2313–2322. [Google Scholar] [CrossRef] [Green Version]
- Naraian, R.; Narayan, O.P.; Srivastava, J. Differential response of oyster shell powder on enzyme profile and nutritional value of oyster mushroom Pleurotus florida PF05. BioMed. Res. Int. 2014, 2014, 386265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiu, S.W.; Chan, Y.H.; Law, S.C.; Cheung, K.T.; Moore, D. Cadmium and manganese in contrast to calcium reduce yield and nutritional values of the edible mushroom Pleurotus pulmonarius. Mycol. Res. 1998, 102, 449–457. [Google Scholar] [CrossRef]
- Zied, D.C.; Pardo-Giménez, A.; Almeida Minhonia, M.T.d.; Villas Boasc, R.L.; Alvarez-Ortid, M.; Pardo-Gonzálezd, J.E. Characterization, feasibility and optimization of Agaricus subrufescens growth based on chemical elements on casing layer. Saudi J. Biol. Sci. 2012, 19, 343–347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Royse, D.J.; Sanchez-Vazquez, J.E. Influence of precipitated calcium carbonate (CaCO3) on shiitake (Lentinula edodes) yield and mushroom size. Bioresour. Technol. 2003, 90, 225–228. [Google Scholar] [CrossRef]
- Bellettini, M.B.; Fiord, F.A.; Maieves, H.A.; Teixeira, G.L.; Ávila, S.; Hornung, P.S.; Júnior, A.M.; Ribani, R.H. Factors affecting mushroom Pleurotus spp. Saudi J. Biol. Sci. 2019, 26, 633–646. [Google Scholar] [CrossRef]
- Zhang, R.Y.; Hu, D.D.; Ma, X.T.; Li, S.G.; Gu, J.G.; Hu, Q.X. Adopting stick spawn reduced the spawn running time and improved mushroom yield and biological efficiency of Pleurotus eryngii. Sci. Hortic. 2014, 175, 156–159. [Google Scholar] [CrossRef]
- Rodriguez Estrada, A.E.; Jimenez-Gasco, M.D.M.; Royse, D.J. Improvement of yield of Pleurotus eryngii var. eryngii by substrate supplementation and use of a casing overlay. Bioresour. Technol. 2009, 100, 5270–5276. [Google Scholar] [CrossRef]
- Jonathan, S.G.; Okon, C.B.; Oyelakin, A.O.; Oluranti, O.O. Nutritional values of oyster mushroom (Pleurotus ostreatus) (Jacq Fr) Kumm cultivated on different agricultural wastes. Nat. Sci. 2012, 10, 186–191. [Google Scholar]
- Chang, S.T.; Hayes, W.A. The Biology and Cultivation of Edible Mushrooms, 1st ed.; A Subsidiary of Harcourt Brace Jovanovich: London, UK, 1978; pp. 521–557. [Google Scholar]
- Ranjbar, M.E.; Olfati, J.A. Evaluation of substrate components on shiitake mushroom properties. Int. J. Veg. Sci. 2017, 23, 145–150. [Google Scholar] [CrossRef]
- Rizki, M.; Tamai, Y. Effects of different nitrogen rich substrates and their combination to the yield performance of oyster mushroom (Pleurotus ostreatus). World J. Microbiol. Biotechnol. 2011, 27, 1695–1702. [Google Scholar] [CrossRef]
- Tabata, T.; Ogura, T. Absorption of calcium and magnesium by the fruiting body of the cultivated mushroom Hypsizigus marmoreus (Peck) Bigelow from sawdust culture media. J. Food Sci. 2003, 68, 76–79. [Google Scholar]
- Qu, J.; Du, F.; Zou, Y.; Zhang, H.; Hu, Q. Study on the formula of mulberry branch and grape branch sawdust for cultivating Pleurotus eryngii. Edible Fungi China 2021, 40, 29–33. [Google Scholar]
- Krystyna, S.-W.; Janusz, S.; Renata, Ł.; Marek, S.; Krzysztof, S.; Kazimierz, G.; Anna, M. Evaluation of polysaccharides content in fruit bodies and their antimicrobial activity of four Ganoderma lucidum (W Curt: Fr) P Karst strains cultivated on different wood type substrates. Acta Soc. Bot. Pol. 2012, 81, 17–21. [Google Scholar]
- Ryu, J.S.; Kim, M.K.; Im, C.H.; Shin, G.P. Development of cultivation media for extending the shelf-life and improving yield of king oyster mushrooms (Pleurotus eryngii). Sci. Hortic. 2015, 193, 121–126. [Google Scholar] [CrossRef]
- Oyetayo, O.V.; Ariyo, O.O. Micro and macronutrient properties of Pleurotus ostreatus (Jacq: Fries) cultivated on different wood substrates. Jordan J. Biol. Sci. 2013, 6, 223–226. [Google Scholar] [CrossRef] [Green Version]
- Akiyama, M. Amino-acid composition of heated scallop shells. J. Fac. Sci. Hokkaido. Univ. 1978, 18, 117–121. [Google Scholar]
- Naraian, R.; Srivastava, J.; Garg, S.K. Influence of dairy spent wash (DSW) on different cultivation phases and yield response of two Pleurotus mushrooms. Ann. Microbiol. 2011, 61, 853–862. [Google Scholar] [CrossRef]
- Singh, M.P.; Singh, V.K. Biodegradation of vegetable and agrowastes by Pleurotus sapidus: A novel strategy to produce mushroom with enhanced yield and nutrition. Cell. Mol. Biol. 2012, 1, 1. [Google Scholar]
- Falandysz, J.; Borovička, J. Macro and trace mineral constituents and radionuclides in mushrooms: Health benefits and risks. Appl. Microbiol. Biotechnol. 2013, 97, 477–501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naito, Y.; Yoshikawa, Y.; Yoshizawa, K.; Takenouchi, A.; Yasui, H. Beneficial effect of bis (Hinokitiolato) Zn complex on high-fat diet-induced lipid accumulation in mouse liver and kidney. In Vivo 2017, 31, 1145–1151. [Google Scholar]
- Jeejeebhoy, K. Zinc: An essential trace element for parenteral nutrition. Gastroenterology 2009, 137, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.M.; Zhang, J.; Wu, L.H.; Zhao, Y.L.; Li, T.; Li, J.Q.; Wang, Y.Z.; Liu, H.G. A mini-review of chemical composition and nutritional value of edible wild-grown mushroom from China. Food Chem. 2014, 151, 279–285. [Google Scholar] [CrossRef] [PubMed]
- Durkan, N.; Ugulu, I.; Unver, M.; Dogan, Y.; Baslar, S. Concentrations of trace elements aluminum, boron, cobalt and tin in various wild edible mushroom species from Buyuk Menderes River Basin of Turkey by ICP-OES. Trace Elem. Electrolytes 2011, 28, 242–248. [Google Scholar] [CrossRef]


{kind=link}
| Substrate | Growth Rates over Time (mm d−1) | Growth Rate (mm d−1) | |||
|---|---|---|---|---|---|
| 7~12 d | 12~17 d | 17~24 d | 24~31 d | ||
| CK | 2.25 | 2.49 | 2.89 | 3.00 | 2.64 ± 0.14 d |
| CK1 | 2.72 | 2.55 | 2.72 | 2.82 | 2.70 ± 0.18 cd |
| T1 | 2.82 | 2.86 | 3.12 | 3.19 | 2.99 ± 0.14 a |
| T2 | 2.65 | 2.26 | 2.99 | 3.38 | 2.81 ± 0.18 bc |
| T3 | 2.53 | 2.35 | 2.82 | 2.77 | 2.62 ± 0.19 d |
| T4 | 2.47 | 2.43 | 3.06 | 3.70 | 2.79 ± 0.20 ab |
| T5 | 2.24 | 2.43 | 2.94 | 2.87 | 2.61 ± 0.10 d |
| Substrate | Fresh Fruiting Body Yield (g bag−1) | Biological Efficiency (%) | Length of Fruit Body (cm) | Thickness of Stipe (cm) | Diameter of Pileus (cm) |
|---|---|---|---|---|---|
| CK | 349.8 ± 38.3 b | 74.0 ± 7.8 b | 14.9 ± 1.4 b | 5.4 ± 0.6 a | 4.5 ± 0.6 de |
| CK1 | 390.1 ± 40.5 a | 82.8 ± 8.6 a | 16.4 ± 1.5 a | 5.5 ± 0.6 a | 4.7 ± 0.6 cd |
| T1 | 399.1 ± 41.2 a | 84.5 ± 8.7 a | 16.0 ± 1.5 a | 5.7 ± 0.8 a | 5.4 ± 0.7 a |
| T2 | 399.5 ± 44.2 a | 84.6 ± 9.4 a | 16.2 ± 1.6 a | 5.4 ± 0.9 a | 5.2 ± 0.7 ab |
| T3 | 393.8 ± 35.2 a | 83.3 ± 7.4 a | 16.3 ± 2.0 a | 5.7 ± 0.5 a | 4.9 ± 0.8 bc |
| T4 | 361.1 ± 34.1 b | 76.4 ± 7.2 b | 15.7 ± 1.6 a | 5.6 ± 0.6 a | 4.8 ± 0.5 cd |
| T5 | 306.0 ± 45.0 c | 64.8 ± 9.5 c | 13.6 ± 1.5 c | 5.5 ± 0.6 a | 4.3 ± 0.5 e |
| Substrate | Fiber | Fat | Ash | Protein | Polysaccharide |
|---|---|---|---|---|---|
| CK | 5.21 ± 0.02 g | 1.71 ± 0.01 c | 5.13 ± 0.02 c | 21.55 ± 0.15 d | 4.26 ± 0.01 d |
| CK1 | 7.40 ± 0.01 b | 1.66 ± 0.01 d | 5.20 ± 0.01 b | 22.65 ± 0.25 c | 3.43 ± 0.02 f |
| T1 | 6.34 ± 0.04 d | 2.11 ± 0.01 a | 4.88 ± 0.02 e | 21.15 ± 0.25 d | 4.54 ± 0.08 b |
| T2 | 5.75 ± 0.04 f | 1.45 ± 0.01 f | 5.05 ± 0.02 d | 22.85 ± 0.25 bc | 4.96 ± 0.04 a |
| T3 | 7.81 ± 0.04 a | 1.32 ± 0.01 g | 5.03 ± 0.07 d | 22.55 ± 0.05 c | 4.18 ± 0.01 e |
| T4 | 6.54 ± 0.01 c | 1.90 ± 0.01 b | 5.32 ± 0.02 a | 23.85 ± 0.35 a | 4.43 ± 0.02 c |
| T5 | 5.90 ± 0.03 e | 1.62 ± 0.01 e | 5.28 ± 0.02 a | 23.25 ± 0.25 b | 4.28 ± 0.03 d |
| Treatments | CK | CK1 | T1 | T2 | T3 | T4 | T5 |
|---|---|---|---|---|---|---|---|
| Phosphorus (mg kg−1) | 736.0 ± 4.2 d | 741 ± 5.0 d | 766.0 ± 8.5 c | 738.0 ± 22.6 d | 744.5 ± 20.5 bc | 791.5 ± 0.7 ab | 797.5 ± 24.7 a |
| Potassium (g kg−1) | 25.4 ± 0.02 bc | 25.9 ± 0.00 b | 25.7 ± 0.06 b | 24.5 ± 0.06 bc | 23.2 ± 0.00 d | 27.4 ± 0.18 a | 25.6 ± 0.04 b |
| Magnesium (g kg−1) | 1.10 ± 0.03 c | 1.09 ± 0.03 c | 1.17 ± 0.00 b | 1.09 ± 0.01 c | 1.21 ± 0.01 b | 1.30 ± 0.08 a | 1.30 ± 0.01 a |
| Sodium(mg kg−1) | 205.0 ± 1.4 d | 190 ± 0.0 e | 230.0 ± 0.0 c | 229.5 ± 14.8 c | 228.0 ± 4.2 c | 298.0 ± 21.2 a | 250.5 ± 7.8 b |
| Calcium (mg kg−1) | 41.0 ± 0.3 e | 63.7 ± 0.6 b | 41.0 ± 0.4 e | 67.2 ± 0.8 a | 48.6 ± 0.4 d | 50.2 ± 0.1 d | 52.3 ± 2.8 c |
| Treatments | CK | CK1 | T1 | T2 | T3 | T4 | T5 |
|---|---|---|---|---|---|---|---|
| Manganese (mg kg−1) | 6.77 ± 0.04 a | 6.35 ± 0.20 b | 5.58 ± 0.04 de | 5.99 ± 0.42 bc | 5.44 ± 0.11 e | 5.87 ± 0.13 cd | 6.27 ± 0.37 b |
| Iron (mg kg−1) | 32.7 ± 0.5 e | 29.3 ± 0.6 f | 34.6 ± 0.0 cd | 35.1 ± 0.5 d | 36.6 ± 1.8 c | 44.1 ± 0.2 a | 39.2 ± 1.5 b |
| Zinc (mg kg−1) | 80.2 ± 1.3 c | 72.7 ± 3.2 e | 82.0 ± 1.8 c | 76.4 ± 2.5 d | 87.8 ± 3.0 b | 80.5 ± 3.5 c | 91.3 ± 0.4 a |
| Boron (mg kg−1) | 19.6 ± 0.5 b | 15.1 ± 0.0 d | 18.8 ± 0.8 bc | 14.2 ± 0.4 e | 18.6 ± 0.7 c | 21.6 ± 1.1 a | 21.4 ± 0.6 a |
| Aluminum (mg kg−1) | 4.53 ± 0.25 d | 4.09 ± 0.00 e | 5.00 ± 0.23 b | 4.73 ± 0.16 c | 4.12 ± 0.08 e | 5.84 ± 0.05 a | 5.07 ± 0.12 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, Y.; Li, Z.; Zhang, H.; Hu, Q.; Zou, Y. Potential Uses of Scallop Shell Powder as a Substrate for the Cultivation of King Oyster Mushroom (Pleurotus eryngii). Horticulturae 2022, 8, 333. https://doi.org/10.3390/horticulturae8040333
Zhou Y, Li Z, Zhang H, Hu Q, Zou Y. Potential Uses of Scallop Shell Powder as a Substrate for the Cultivation of King Oyster Mushroom (Pleurotus eryngii). Horticulturae. 2022; 8(4):333. https://doi.org/10.3390/horticulturae8040333
Chicago/Turabian StyleZhou, Yuanyuan, Zihao Li, Haijun Zhang, Qingxiu Hu, and Yajie Zou. 2022. "Potential Uses of Scallop Shell Powder as a Substrate for the Cultivation of King Oyster Mushroom (Pleurotus eryngii)" Horticulturae 8, no. 4: 333. https://doi.org/10.3390/horticulturae8040333