
Changes in Morpho-Anatomical and Eco-Physiological Responses of Viburnum tinus L. var lucidum as Modulated by Sodium Chloride and Calcium Chloride Salinization

 , , , , and
, , , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material, Cultivation Conditions, and Experimental Design
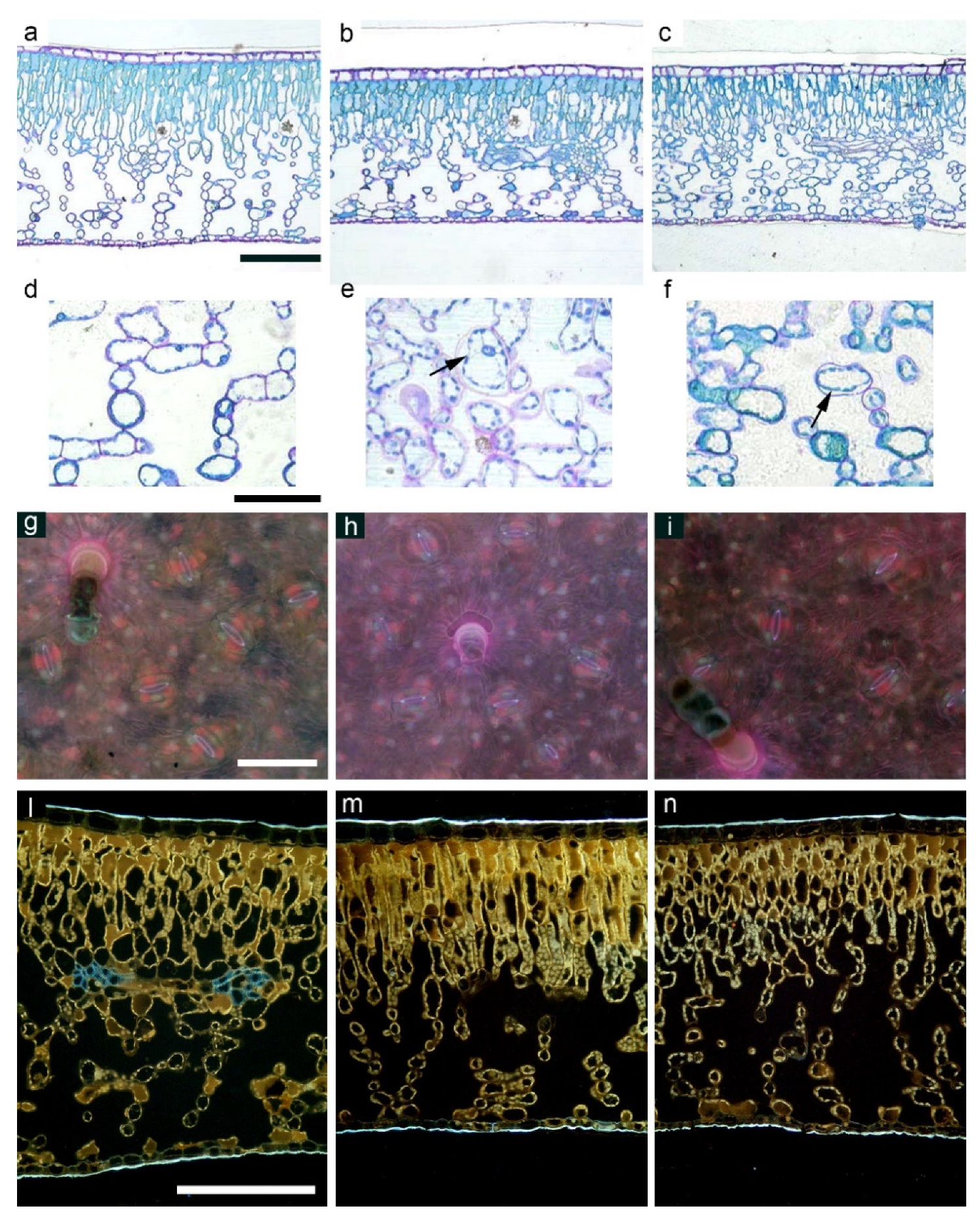
2.2. Biometry, Morphological, and Anatomical Leaf Traits
2.3. Leaf Gas-Exchanges and Chlorophyll a Fluorescence Emission
2.4. Mineral Composition and Organic Acids
2.5. Data Elaboration
3. Results
3.1. Growth and Morpho-Anatomical Traits
3.2. Leaf Gas-Exchanges and Chlorophyll a Fluorescence Emission
3.3. Mineral and Organic Acids
4. Discussion
4.1. The Effect of Salinity Treatment on Morpho-Anatomical Traits
4.2. The Effect of Salinity Treatment on Photosynthesis
4.3. The Effect of Salinity Treatment on Ion Accumulation
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ipni. The International Plant Names Index; The Royal Botanic Gardens, Kew, Harvard University Herbaria & Libraries and Australian National Botanic Gardens, 2021; Available online: http://www.ipni.org (accessed on 1 October 2021).
- Tribulato, A.; Toscano, S.; Di Lorenzo, V.; Romano, D. Effects of water stress on gas exchange, water relations and leaf structure in two ornamental shrubs in the Mediterranean area. Agronomy 2019, 9, 381. [Google Scholar] [CrossRef] [Green Version]
- Cassaniti, C.; Leonardi, C.; Flowers, T.J. The effects of sodium chloride on ornamental shrubs. Sci. Hortic. 2009, 122, 586–593. [Google Scholar] [CrossRef]
- Toscano, S.; Ferrante, A.; Romano, D.; Tribulato, A. Interactive Effects of Drought and Saline Aerosol Stress on Morphological and Physiological Characteristics of Two Ornamental Shrub Species. Horticulturae 2021, 7, 517. [Google Scholar] [CrossRef]
- Yeager, T.H.; von Merveldt, J.K.; Larsen, C.A. Ornamental plant response to percentage of reclaimed water irrigation. HortScience 2010, 45, 1610–1615. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Chen, J.J.; Xing, H.; Paudel, A.; Niu, G.; Chappell, M. Growth, visual quality, and morphological responses of 12 Viburnum taxa to saline water irrigation. HortScience 2020, 55, 1233–1241. [Google Scholar] [CrossRef]
- Giordano, M.; Petropoulos, S.A.; Cirillo, C.; Rouphael, Y. Biochemical, physiological, and molecular aspects of ornamental plants adaptation to deficit irrigation. Horticulturae 2021, 7, 107. [Google Scholar] [CrossRef]
- De Micco, V.; Aronne, G. Morpho-anatomical Traits for Plant Adaptation to Drought. In Plant Responses to Drought Stress; Springer: Berlin/Heidelberg, Germany, 2012; pp. 37–61. [Google Scholar] [CrossRef]
- Tester, M.; Davenport, R. Na+ tolerance and Na+ transport in higher plants. Ann. Bot. 2003, 91, 503–527. [Google Scholar] [CrossRef]
- Ruiz-Sánchez, M.C.; Domingo, R.; Torrecillas, A.; Pérez-Pastor, A. Water stress preconditioning to improve drought resistance in young apricot plants. Plant Sci. 2000, 156, 245–251. [Google Scholar] [CrossRef] [Green Version]
- Colmer, T.; Munns, R.; Flowers, T. Improving salt tolerance of wheat and barley: Future prospects. Aust. J. Exp. Agric. 2005, 45, 1425–1443. [Google Scholar] [CrossRef]
- Srinivasan, V.; Kumar, P.; Long, S.P. Decreasing, not increasing, leaf area will raise crop yields under global atmospheric change. Glob. Chang. Biol. 2017, 23, 1626–1635. [Google Scholar] [CrossRef] [Green Version]
- Amitrano, C.; Arena, C.; Cirillo, V.; De Pascale, S.; De Micco, V. Leaf morpho-anatomical traits in Vigna radiata L. affect plant photosynthetic acclimation to changing vapor pressure deficit. Environ. Exp. Bot. 2021, 186, 104453. [Google Scholar] [CrossRef]
- Acosta-Motos, J.-R.; Diaz-Vivancos, P.; Álvarez, S.; Fernández-García, N.; Sanchez-Blanco, M.J.; Hernández, J.A. Physiological and biochemical mechanisms of the ornamental Eugenia myrtifolia L. plants for coping with NaCl stress and recovery. Planta 2015, 242, 829–846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez-Bellot, M.J.; Nortes, P.A.; Ortuño, M.F.; Romero, C.; Fernandez-Garcia, N.; Sánchez-Blanco, M.J. Influence of arbuscular mycorrhizal fungi and treated wastewater on water relations and leaf structure alterations of Viburnum tinus L. plants during both saline and recovery periods. J. Plant Phyisiol. 2015, 188, 96–105. [Google Scholar] [CrossRef] [PubMed]
- Fraser, L.H.; Greenall, A.; Carlyle, C.; Turkington, R.; Friedman, C.R. Adaptive phenotypic plasticity of Pseudoroegneria spicata: Response of stomatal density, leaf area and biomass to changes in water supply and increased temperature. Ann. Bot. 2009, 103, 769–775. [Google Scholar] [CrossRef] [Green Version]
- Acosta-Motos, J.R.; Diaz-Vivancos, P.; Álvarez, S.; Fernández-García, N.; Sánchez-Blanco, M.J.; Hernández, J.A. NaCl-induced physiological and biochemical adaptative mechanisms in the ornamental Myrtus communis L. plants. J. Plant Physiol. 2015, 183, 41–51. [Google Scholar] [CrossRef] [Green Version]
- Amitrano, C.; Arena, C.; Rouphael, Y.; De Pascale, S.; De Micco, V. Vapour pressure deficit: The hidden driver behind plant morphofunctional traits in controlled environments. Ann. Appl. Biol. 2019, 175, 313–325. [Google Scholar] [CrossRef]
- Acosta-Motos, J.R.; Ortuño, M.F.; Bernal-Vicente, A.; Diaz-Vivancos, P.; Sanchez-Blanco, M.J.; Hernandez, J.A. Plant responses to salt stress: Adaptive mechanisms. Agronomy 2017, 7, 18. [Google Scholar] [CrossRef] [Green Version]
- Rouphael, Y.; De Micco, V.; Arena, C.; Raimondi, G.; Colla, G.; De Pascale, S. Effect of Ecklonia maxima seaweed extract on yield, mineral composition, gas exchange, and leaf anatomy of zucchini squash grown under saline conditions. J. Appl. Phycol. 2017, 29, 459–470. [Google Scholar] [CrossRef]
- De Micco, V.; Arena, C.; Amitrano, C.; Rouphael, Y.; De Pascale, S.; Cirillo, C. Effects of NaCl and CaCl2 Salinization on Morpho-anatomical and Physiological Traits of Potted Callistemon citrinus Plants. Forests 2021, 12, 1666. [Google Scholar] [CrossRef]
- Gori, R.; Lubello, C.; Ferrini, F.; Nicese, F.; Coppini, E. Reuse of industrial wastewater for the irrigation of ornamental plants. Water Sci. Technol. 2008, 57, 883–889. [Google Scholar] [CrossRef]
- García-Caparrós, P.; Lao, M.T. The effects of salt stress on ornamental plants and integrative cultivation practices. Sci. Hortic. 2018, 240, 430–439. [Google Scholar] [CrossRef]
- Cirillo, C.; De Micco, V.; Arena, C.; Carillo, P.; Pannico, A.; De Pascale, S.; Rouphael, Y. Biochemical, Physiological and Anatomical Mechanisms of Adaptation of Callistemon citrinus and Viburnum lucidum to NaCl and CaCl2 Salinization. Front. Plant Sci. 2019, 10, 742. [Google Scholar] [CrossRef] [PubMed]
- Arena, C.; Conti, S.; Francesca, S.; Melchionna, G.; Hájek, J.; Barták, M.; Barone, A.; Rigano, M.M. Eco-physiological screening of different tomato genotypes in response to high temperatures: A combined field-to-laboratory approach. Plants 2020, 9, 508. [Google Scholar] [CrossRef] [Green Version]
- Feder, N.; O’brien, T. Plant microtechnique: Some principles and new methods. Am. J. Bot. 1968, 55, 123–142. [Google Scholar] [CrossRef]
- Fukazawa, K. Ultraviolet microscopy. In Methods in Lignin Chemistry; Springer: Berlin/Heidelberg, Germany, 1992; pp. 110–121. [Google Scholar]
- Van Buggenhout, S.; Grauwet, T.; Van Loey, A.; Hendrickx, M. Structure/processing relation of vacuum infused strawberry tissue frozen under different conditions. Eur. Food Res. Technol. 2008, 226, 437–448. [Google Scholar] [CrossRef]
- De Micco, V.; Arena, C.; Aronne, G. Anatomical alterations of Phaseolus vulgaris L. mature leaves irradiated with X-rays. Plant Biol. 2014, 16, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Lichtenthaler, H.K.; Babani, F. Light adaptation and senescence of the photosynthetic apparatus. Changes in pigment composition, chlorophyll fluorescence parameters and photosynthetic activity. In Chlorophyll a Fluorescence; Springer: Berlin/Heidelberg, Germany, 2004; pp. 713–736. [Google Scholar]
- Genty, B.; Briantais, J.-M.; Baker, N.R. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. BBA-Gen. Subj. 1989, 990, 87–92. [Google Scholar] [CrossRef]
- Bilger, W.; Björkman, O. Role of the xanthophyll cycle in photoprotection elucidated by measurements of light-induced absorbance changes, fluorescence and photosynthesis in leaves of Hedera canariensis. Photosynth. Res. 1990, 25, 173–185. [Google Scholar] [CrossRef]
- Ashraf, M.A.; Akbar, A.; Parveen, A.; Rasheed, R.; Hussain, I.; Iqbal, M. Phenological application of selenium differentially improves growth, oxidative defense and ion homeostasis in maize under salinity stress. Plant Physiol. Biochem. 2018, 123, 268–280. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Aslam, M.T.; Alhammad, B.A.; Hassan, M.U.; Maqbool, R.; Chattha, M.U.; Khan, I.; Gitari, H.I.; Uslu, O.S.; Roy, R. Salinity stress in wheat: Effects, mechanisms and management strategies. Phyton-Int. J. Exp. Bot. 2021. [Google Scholar] [CrossRef]
- Jafari, S.; Garmdareh, S.E.H. Effects of salinity on morpho-physiological, and biochemical characteristics of stock plant (Matthiola incana L.). Sci. Hortic. 2019, 257, 108731. [Google Scholar] [CrossRef]
- Cirillo, C.; Rouphael, Y.; Caputo, R.; Raimondi, G.; Sifola, M.; De Pascale, S. Effects of high salinity and the exogenous application of an osmolyte on growth, photosynthesis, and mineral composition in two ornamental shrubs. J. Hortic. Sci. Biotechnol. 2016, 91, 14–22. [Google Scholar] [CrossRef]
- Kummerow, J. Structural aspects of shrubs in Mediterranean-type plant communities. In Proceedings of the Jornadas Sobre las Bases Ecol¢ Gicas para la Gesti¢ n en Ecosistemas Terrestres, Zaragoza, Spain, 13–15 June 1988. [Google Scholar]
- Bozkurt, S.; Keskin, M. Effect of deficit irrigation on the estimation of cucumber leaf area under greenhouse conditions. J. Agric. Biol. 2018, 20, 877–882. [Google Scholar]
- Blanco, F.F.; Folegatti, M.V. Estimation of leaf area for greenhouse cucumber by linear measurements under salinity and grafting. Sci. Agric. 2005, 62, 305–309. [Google Scholar] [CrossRef] [Green Version]
- Asmar, S.; Castro, E.; Pasqual, M.; Pereira, F.; Soares, J. Changes in leaf anatomy and photosynthesis of micropropagated banana plantlets under different silicon sources. Sci. Hortic. 2013, 161, 328–332. [Google Scholar] [CrossRef]
- Cirillo, C.; Rouphael, Y.; Caputo, R.; Raimondi, G.; De Pascale, S. The influence of deficit irrigation on growth, ornamental quality, and water use efficiency of three potted Bougainvillea genotypes grown in two shapes. HortScience 2014, 49, 1284–1291. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.J.; Xing, H.; Paudel, A.; Sun, Y.; Niu, G.; Chappell, M. Gas Exchange and Mineral Nutrition of 12 Viburnum Taxa irrigated with Saline Water. HortScience 2020, 55, 1242–1250. [Google Scholar] [CrossRef]
- Anjum, S.A.; Xie, X.-Y.; Wang, L.-C.; Saleem, M.F.; Man, C.; Lei, W. Morphological, physiological and biochemical responses of plants to drought stress. Afr. J. Agric. Res. 2011, 6, 2026–2032. [Google Scholar]
- Campbell, D.R.; Wu, C.A.; Travers, S.E. Photosynthetic and growth responses of reciprocal hybrids to variation in water and nitrogen availability. Am. J. Bot. 2010, 97, 925–933. [Google Scholar] [CrossRef] [Green Version]
- Raven, J.A. Speedy small stomata? J. Exp. Bot. 2014, 65, 1415–1424. [Google Scholar] [CrossRef] [Green Version]
- Roelfsema, M.R.G.; Hedrich, R. Studying guard cells in the intact plant: Modulation of stomatal movement by apoplastic factors. New Phytol. 2002, 153, 425–431. [Google Scholar] [CrossRef] [PubMed]
- Tränkner, M.; Tavakol, E.; Jákli, B. Functioning of potassium and magnesium in photosynthesis, photosynthate translocation and photoprotection. Physiol. Plant. 2018, 163, 414–431. [Google Scholar] [CrossRef] [Green Version]
- Erel, R.; Yermiyahu, U.; Ben-Gal, A.; Dag, A.; Shapira, O.; Schwartz, A. Modification of non-stomatal limitation and photoprotection due to K and Na nutrition of olive trees. Physiol. Plant Physiol. 2015, 177, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Jákli, B.; Tavakol, E.; Tränkner, M.; Senbayram, M.; Dittert, K. Quantitative limitations to photosynthesis in K deficient sunflower and their implication on water-use efficiency. Physiol. Plant. Physiol. 2017, 209, 20–30. [Google Scholar] [CrossRef] [PubMed]
- Moriana, A.; Pérez-López, D.; Prieto, M.; Ramírez-Santa-Pau, M.; Pérez-Rodriguez, J. Midday stem water potential as a useful tool for estimating irrigation requirements in olive trees. Agric. Water Manag. 2012, 112, 43–54. [Google Scholar] [CrossRef]
- Amitrano, C.; Rouphael, Y.; Pannico, A.; De Pascale, S.; De Micco, V. Reducing the Evaporative Demand Improves Photosynthesis and Water Use Efficiency of Indoor Cultivated Lettuce. Agronomy 2021, 11, 1396. [Google Scholar] [CrossRef]
- De Micco, V.; Amitrano, C.; Stinca, A.; Izzo, L.G.; Zalloni, E.; Balzano, A.; Barile, R.; Conti, P.; Arena, C. Dust accumulation due to anthropogenic impact induces anatomical and photochemical changes in leaves of Centranthus ruber growing on the slope of the Vesuvius volcano. Plant Biol. 2019, 22 (Suppl. 1), 93–102. [Google Scholar] [CrossRef]
- Arena, C.; Vitale, L.; De Santo, A.V. Paraheliotropism in Robinia pseudoacacia L.: An efficient strategy to optimise photosynthetic performance under natural environmental conditions. Plant Biol. 2008, 10, 194–201. [Google Scholar] [CrossRef]
- Vitale, L.; Arena, C.; De Santo, A.V. Seasonal changes in photosynthetic activity and photochemical efficiency of the Mediterranean shrub Phillyrea angustifolia L. Plant Biosyst. 2012, 146, 443–450. [Google Scholar] [CrossRef]
- Lattanzio, V.; Kroon, P.A.; Quideau, S.; Treutter, D. Plant phenolics—secondary metabolites with diverse functions. Recent Adv. Polyphen. Res. 2008, 1, 1–35. [Google Scholar]
- De Micco, V.; Arena, C.; Vitale, L.; Aronne, G.; Virzo De Santo, A. Anatomy and photochemical behaviour of Mediterranean Cistus incanus winter leaves under Natural outdoor and warmer indoor conditions. Botany 2011, 89, 677–688. [Google Scholar] [CrossRef]
- Arena, C.; Vitale, L.; Virzo De Santo, A. Photosynthesis and photoprotective strategies in Laurus nobilis L. and Quercus ilex L. under summer drought and winter cold. Plant Biosyst. 2008, 142, 472–479. [Google Scholar] [CrossRef]
- Karabourniotis, G.; Liakopoulos, G.; Nikolopoulos, D.; Bresta, P. Protective and defensive roles of non-glandular trichomes against multiple stresses: Structure–function coordination. J. For. Res. 2020, 31, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Franco, J.; Bañón, S.; Vicente, M.; Miralles, J.; Martínez-Sánchez, J. Root development in horticultural plants grown under abiotic stress conditions–a review. J. Hortic. Sci. 2011, 86, 543–556. [Google Scholar]
- Sánchez-Blanco, M.J.; Alvarez, S.; Ortuno, M.F.; Ruiz-Sánchez, M.C. Root system response to drought and salinity: Root distribution and water transport. Root Eng. 2014, 40, 325–352. [Google Scholar]
- Grattan, S.; Grieve, C. Salinity–mineral nutrient relations in horticultural crops. Sci. Hortic. 1998, 78, 127–157. [Google Scholar] [CrossRef]
- Munns, R.; Gilliham, M. Salinity tolerance of crops–what is the cost? New phytol. 2015, 208, 668–673. [Google Scholar]
- Dreyer, I.; Uozumi, N. Potassium channels in plant cells. FEBS J. 2011, 278, 4293–4303. [Google Scholar] [CrossRef]
- Anschütz, U.; Becker, D.; Shabala, S. Going beyond nutrition: Regulation of potassium homoeostasis as a common denominator of plant adaptive responses to environment. Physiol. Plant Physiol. 2014, 171, 670–687. [Google Scholar] [CrossRef]
- Shabala, S.; Pottosin, I. Regulation of potassium transport in plants under hostile conditions: Implication for abiotic and biotic stress tolerance. Physiol. Plant 2014, 151, 257–279. [Google Scholar] [CrossRef]
- Wu, H.; Zhang, X.; Giraldo, J.P.; Shabala, S. It is not all about sodium: Revealing tissue specificity and signalling roles of potassium in plant responses to salt stress. Plant Soil 2018, 431, 1–17. [Google Scholar] [CrossRef]
- Wang, M.; Zheng, Q.; Shen, Q.; Guo, S. The critical role of potassium in plant stress response. Int. J. Mol. Sci. 2013, 14, 7370–7390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perera, L.; Robinson, M.; Mansfield, T. Responses of the stomata of Aster tripolium to calcium and sodium ions in relation to salinity tolerance. J. Exp. Bot. 1995, 46, 623–629. [Google Scholar] [CrossRef]
- Albino-Garduño, R.; Zavaleta-Mancera, H.; Ruiz-Posadas, L.; Sandoval-Villa, M.; Castillo-Morales, A. Response of gerbera to calcium in hydroponics. J. Plant Nutr. 2007, 31, 91–101. [Google Scholar] [CrossRef]
- Bryla, D.R.; Scagel, C.F.; Lukas, S.B.; Sullivan, D.M. Ion-specific Limitations of Sodium Chloride and Calcium Chloride on Growth, Nutrient Uptake, and Mycorrhizal Colonization in Northern and Southern Highbush Blueberry. J. Am. Soc. Hort. Sci. 2021, 146, 399–410. [Google Scholar] [CrossRef]
- Franceschi, V.R.; Horner, H.T. Calcium oxalate crystals in plants. Bot. Rev 1980, 46, 361–427. [Google Scholar] [CrossRef]
- Marschner, H. Marschner’s Mineral Nutrition of Higher Plants; Elsevier/Academic Press: Amsterdam, The Netherlands, 2011; p. 684. [Google Scholar]
- Franco-Navarro, J.D.; Brumós, J.; Rosales, M.A.; Cubero-Font, P.; Talón, M.; Colmenero-Flores, J.M. Chloride regulates leaf cell size and water relations in tobacco plants. J. Exp. Bot. 2016, 67, 873–891. [Google Scholar] [CrossRef] [Green Version]
- Franco-Navarro, J.D.; Rosales, M.A.; Cubero-Font, P.; Calvo, P.; Álvarez, R.; Diaz-Espejo, A.; Colmenero-Flores, J.M. Chloride as a macronutrient increases water-use efficiency by anatomically driven reduced stomatal conductance and increased mesophyll diffusion to CO2. Plant J. 2019, 99, 815–831. [Google Scholar]
- Wu, H.; Li, Z. The importance of Cl− exclusion and vacuolar Cl− sequestration: Revisiting the role of Cl− transport in plant salt tolerance. Front. Plant. Sci. 2019, 10, 1418. [Google Scholar] [CrossRef]


{kind=link}
| CV (m3 Plant−1) | TLA/PH (m2 m−1) | LDW (g Plant−1) | SDW (g Plant−1) | ASL (cm2) | SLW (g DW cm−2) | |
|---|---|---|---|---|---|---|
| Salt treatment | ||||||
| C | 1.61 ± 0.09 a | 2.41 ± 0.08 a | 133.18 ± 6.18 a | 80.93 ± 3.12 a | 28.22 ± 0.87 a | 81.83 ± 1.51 b |
| CaCl2_T | 1.02 ± 0.15 b | 1.88 ± 0.10 b | 92.50 ± 6.03 b | 69.64 ± 3.16 b | 25.10 ± 0.81 b | 98.73 ± 1.53 a |
| NaCl_T | 0.93 ± 0.06 b | 1.54 ± 0.08 c | 81.06 ± 4.32 b | 64.85 ± 2.98 b | 21.45 ± 1.00 c | 101.03 ± 2.07 a |
| Significance | *** | *** | *** | ** | *** | *** |
| PP Thickness (μm) | SP Thickness (μm) | Cell Area (μm2) | Stomatal Size (μm2) | Stomatal Area (%) | Trichome Frequency (n mm−1) | Phenolics in PP (%) | Phenolics in PS (%) | |
|---|---|---|---|---|---|---|---|---|
| Salt treatment | ||||||||
| C | 85.9 ± 3.50 a | 176 ± 3.79 a | 188 ± 6.80 | 463 ± 7.35 a | 4.33 ± 0.08 a | 3.04 ± 0.39 b | 57.9 ± 1.33 a | 13.0 ± 1.13 a |
| CaCl2_T | 74.3 ± 1.87 c | 142 ± 3.92 b | 209 ± 9.40 | 333 ± 6.97 b | 3.13 ± 0.06 c | 6.90 ± 0.63 a | 47.6 ± 2.66 b | 9.95 ± 1.24 b |
| NaCl_T | 81.2 ± 2.87 b | 143 ± 4.53 b | 197 ± 7.21 | 339 ± 5.65 b | 3.36 ± 0.06 b | 5.73 ± 0.53 a | 48.0 ± 22.7 b | 10.1 ± 1.25 b |
| Significance | * | *** | NS | *** | *** | *** | * | ** |
| Pn (µmol CO2 m−2 s−1) | Ci (µmol mol−1) | gs (mmol m−2 s−1) | E (mol H2O m−2 s−1) | iWUE (µmol CO2 mol−1 H2O) | |
|---|---|---|---|---|---|
| Salt treatment | |||||
| C | 7.50 ± 0.34 a | 191.28 ± 11.69 a | 97.55 ± 14.20 a | 2.11 ± 0.13 a | 86.22 ± 12.19 a |
| CaCl2_T | 3.15 ± 0.35 c | 171.70 ± 11.97 a | 31.64 ± 14.54 b | 0.93 ± 0.13 c | 122.57 ± 12.48 a |
| NaCl_T | 4.73 ± 0.37 b | 172.29 ± 12.67 a | 64.61 ± 15.39 ab | 1.36 ± 0.14 b | 113.28 ± 13.21 a |
| DAT | |||||
| 106 | 5.12 ± 0.32 a | 173.33 ± 10.92 a | 58.34 ± 13.27 a | 1.22 ± 0.12 b | 123.47 ± 11.39 a |
| 121 | 5.13 ± 0.25 a | 183.52 ± 8.74 a | 70.86 ± 10.62 a | 1.71 ± 0.10 a | 91.25 ± 9.11 b |
| Significance | |||||
| S | *** | NS | ** | *** | NS |
| D | NS | NS | NS | ** | * |
| S × D | NS | NS | NS | NS | NS |
| ФPSII | qP | qN | Fv/Fo | |
|---|---|---|---|---|
| Salt treatment | ||||
| C | 0.641 ± 0.009 a | 0.923 ± 0.009 a | 0.509 ± 0.021 b | 1.842 ± 0.071 a |
| CaCl2_T | 0.540 ± 0.018 b | 0.874 ± 0.014 c | 0.597 ± 0.038 a | 1.221 ± 0.123 b |
| NaCl_T | 0.615 ± 0.014 a | 0.905 ± 0.012 b | 0.565 ± 0.042 a | 1.686 ± 0.140 a |
| DAT | ||||
| 106 | 0.593 ± 0.019 a | 0.873 ± 0.014 b | 0.499 ± 0.032 b | 1.667 ± 0.143 a |
| 121 | 0.604 ± 0.019 a | 0.928 ± 0.015 a | 0.615 ± 0.035 a | 1.499 ± 0.154 a |
| Significance | ||||
| S | *** | *** | ** | *** |
| D | NS | *** | *** | NS |
| S × D | NS | * | NS | NS |
| Na+ (mg Plant−1) | K+ (mg Plant−1) | Ca2+ (mg Plant−1) | Mg2+ (mg Plant−1) | Cl− (mg Plant−1) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Leaf | Root | Leaf | Root | Leaf | Root | Leaf | Root | Leaf | Root | |
| Salt treatment | ||||||||||
| C | 40.8 ± 5.02 b | 209 ± 23.0 b | 3744 ± 218.8 a | 1216 ± 136.5 a | 244 ± 34.3 b | 136 ± 12.3 b | 191 ± 13.9 a | 167 ± 22.6 a | 393 ± 25.1 b | 236 ± 33.4 c |
| CaCl2_T | 53.3 ± 11.2 b | 137 ± 15.7 b | 1837 ± 162.5 b | 788 ± 73.9 b | 679 ± 95.5 a | 227 ± 24.8 a | 150 ± 16.1 b | 123 ± 7.17 ab | 1209 ± 127 a | 832 ± 96.9 a |
| NaCl_T | 335 ± 54.5 a | 716 ± 42.7 a | 1607 ± 243.9 b | 418 ± 28.7 c | 73.6 ± 13.12 b | 72.4 ± 10.2 c | 75.8 ± 8.77 c | 98.3 ± 11.2 b | 588 ± 106 b | 593 ± 32.8 b |
| Significance | *** | *** | *** | *** | *** | *** | *** | * | *** | *** |
| Quinic ac. (g kg−1 DW) | Acetic ac. (g kg−1 DW) | Formic ac. (g kg−1 DW) | Malic ac. (g kg−1 DW) | Oxalic ac. (g kg−1 DW) | Fumaric ac. (g kg−1 DW.) | Citric ac. (g kg−1 DW) | Isocitric ac. (g kg−1 DW) | |
|---|---|---|---|---|---|---|---|---|
| Salt treatment | ||||||||
| C | 27.0 ± 2.28 a | 3.04 ± 0.18 | 0.38 ± 0.03 | 25.8 ± 2.5 a | 3.30 ± 0.39 b | 1.36 ± 0.10 | 8.10 ± 1.48 | 0.21 ± 0.04 |
| CaCl2_T | 13.3 ± 2.29 b | 2.39 ± 0.28 | 0.38 ± 0.03 | 26.3 ± 3.5 a | 1.54 ± 0.19 c | 1.27 ± 0.14 | 9.43 ± 1.64 | 0.27 ± 0.03 |
| NaCl_T | 19.6 ± 2.89 ab | 3.01 ± 0.42 | 0.36 ± 0.08 | 14.0 ± 2.3 b | 5.66 ± 0.84 a | 1.30 ± 0.21 | 8.56 ± 1.43 | 0.28 ± 0.03 |
| Significance | ** | NS | NS | * | *** | NS | NS | NS |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Micco, V.; Arena, C.; Amitrano, C.; Rouphael, Y.; De Pascale, S.; Cirillo, C. Changes in Morpho-Anatomical and Eco-Physiological Responses of Viburnum tinus L. var lucidum as Modulated by Sodium Chloride and Calcium Chloride Salinization. Horticulturae 2022, 8, 119. https://doi.org/10.3390/horticulturae8020119
De Micco V, Arena C, Amitrano C, Rouphael Y, De Pascale S, Cirillo C. Changes in Morpho-Anatomical and Eco-Physiological Responses of Viburnum tinus L. var lucidum as Modulated by Sodium Chloride and Calcium Chloride Salinization. Horticulturae. 2022; 8(2):119. https://doi.org/10.3390/horticulturae8020119
Chicago/Turabian StyleDe Micco, Veronica, Carmen Arena, Chiara Amitrano, Youssef Rouphael, Stefania De Pascale, and Chiara Cirillo. 2022. "Changes in Morpho-Anatomical and Eco-Physiological Responses of Viburnum tinus L. var lucidum as Modulated by Sodium Chloride and Calcium Chloride Salinization" Horticulturae 8, no. 2: 119. https://doi.org/10.3390/horticulturae8020119